miR-141表達(dá)抑制增強(qiáng)結(jié)腸癌細(xì)胞對(duì)5-Fu藥物敏感性的研究
2016-07-01 07:29:36馬一楠金迎迎王亞利陳青娟
山西醫(yī)科大學(xué)學(xué)報(bào) 2016年6期
馬一楠, 金迎迎*, 王亞利, 陳青娟, 衛(wèi) 陽(yáng)
(1西安交通大學(xué)第二附屬醫(yī)院腫瘤科,西安 710004; 2咸陽(yáng)市中心醫(yī)院腫瘤科; 3西安交通大學(xué)第二附屬醫(yī)院科研中心實(shí)驗(yàn)室; *通訊作者,E-mail:yingyingjin717@sina.com)
miR-141表達(dá)抑制增強(qiáng)結(jié)腸癌細(xì)胞對(duì)5-Fu藥物敏感性的研究
馬一楠1, 金迎迎1*, 王亞利1, 陳青娟2, 衛(wèi)陽(yáng)3
(1西安交通大學(xué)第二附屬醫(yī)院腫瘤科,西安710004;2咸陽(yáng)市中心醫(yī)院腫瘤科;3西安交通大學(xué)第二附屬醫(yī)院科研中心實(shí)驗(yàn)室;*通訊作者,E-mail:yingyingjin717@sina.com)
摘要:目的建立人結(jié)腸癌5-氟尿嘧啶(5-Fu)耐藥細(xì)胞株,并探討miR-141靶向作用PPP2R1B介導(dǎo)結(jié)腸癌細(xì)胞對(duì)5-Fu耐藥的機(jī)制。方法通過(guò)藥物敏感性實(shí)驗(yàn),選取5-Fu敏感細(xì)胞COLO-320細(xì)胞株采取大劑量沖擊聯(lián)合劑量遞增法,誘導(dǎo)篩選5-Fu耐藥結(jié)腸癌細(xì)胞株,穩(wěn)定傳代。四甲基偶氮唑鹽法(MTT)檢測(cè)耐藥細(xì)胞和親本細(xì)胞的半數(shù)抑制濃度(IC50)。PCR的微陣列技術(shù)比較親本細(xì)胞和5-Fu耐藥細(xì)胞差異表達(dá)的miRNA分子,從中篩選出1個(gè)差異表達(dá)的miR-141分子。利用數(shù)據(jù)庫(kù)Targetscan和miRBase database預(yù)測(cè)miR-141的靶基因?yàn)镻PP2R1B,利用雙熒光素酶報(bào)告基因檢測(cè)進(jìn)行鑒定。基因敲除miR-141后分析miR-141對(duì)PPP2R1B的調(diào)控作用。結(jié)果COLO-320細(xì)胞株經(jīng)大劑量沖擊聯(lián)合劑量遞增的方法誘導(dǎo)后可在5.0 μmol/L的5-Fu培養(yǎng)液中穩(wěn)定增殖,具有耐藥性,命名為COLO-320/5-Fu,該細(xì)胞株5-Fu的IC50顯著高于親代細(xì)胞(P<0.05)。芯片結(jié)果顯示共有13個(gè)miRNAs在耐藥細(xì)胞株中差異性表達(dá),其中miRNA-141表達(dá)增加最為顯著(P<0.05)。敲除該miR-141后,耐藥細(xì)胞的5-Fu敏感性顯著增加,凋亡比例增加(P<0.05)。結(jié)果顯示,PPP2R1B為miR-141的靶基因,miRNA-141表達(dá)抑制顯著上調(diào)PPP2R1B的表達(dá)水平并進(jìn)而影響Akt磷酸化過(guò)程。結(jié)論本實(shí)驗(yàn)成功構(gòu)建COLO-320/5-Fu耐藥細(xì)胞株,miR-141可能通過(guò)靶向調(diào)控PPP2R1B,進(jìn)而參與調(diào)控結(jié)腸癌細(xì)胞對(duì)5-Fu的耐藥。
關(guān)鍵詞:結(jié)腸癌;5-氟尿嘧啶;蛋白質(zhì)磷酸酶2調(diào)節(jié)亞基1B;耐藥性;微小RNA
結(jié)腸癌是嚴(yán)重危及人類健康的常見(jiàn)惡性腫瘤之一,美國(guó)癌癥數(shù)據(jù)顯示其發(fā)病率居全球第三,結(jié)腸癌相關(guān)死亡率位居全球第二[1]。5-氟尿嘧啶(5-Fu)是目前治療結(jié)腸癌使用最廣的化療藥物。其主要作用為通過(guò)阻斷胸苷合成酶的活性,中斷嘧啶胸苷的合成,進(jìn)而影響腫瘤細(xì)胞DNA復(fù)制。另外,5-Fu可以通過(guò)誘導(dǎo)細(xì)胞周期停滯、促進(jìn)腫瘤細(xì)胞凋亡來(lái)達(dá)到治療腫瘤的目的[2]。5-Fu常與其他化療藥物聯(lián)合應(yīng)用,雖能有效降低結(jié)腸癌患者的死亡率,但復(fù)發(fā)和轉(zhuǎn)移阻礙了其療效的進(jìn)一步提高。研究發(fā)現(xiàn)5-Fu耐藥是導(dǎo)致患者預(yù)后不良的主要原因[3]。因此,盡早發(fā)現(xiàn)結(jié)腸癌耐藥患者,探索5-Fu的耐藥機(jī)制,并對(duì)其進(jìn)行干預(yù)治療,對(duì)改善耐藥患者的預(yù)后尤為重要。
研究發(fā)現(xiàn)耐藥性的產(chǎn)生與許多分子機(jī)制相關(guān),包括周期阻滯、凋亡抑制、代謝異常等[4-7]。微小RNA(microRNAs,miRNA)是一類長(zhǎng)約22 nt非編碼RNA分子,其主要是通過(guò)結(jié)合其靶基因mRNA的3′-非編碼區(qū)以引發(fā)序列特異性mRNA切割或抑制翻譯調(diào)節(jié)基因表達(dá)[8]。其中miRNA調(diào)節(jié)機(jī)體近1/3生命活動(dòng),與機(jī)體生長(zhǎng)、發(fā)育、增殖、凋亡等代謝活動(dòng)密切相關(guān),病理狀態(tài)下主要參與調(diào)控腫瘤的惡性生物學(xué)行為[9,10]。大量研究表明miRNA與腫瘤的發(fā)生密切相關(guān),通過(guò)轉(zhuǎn)錄后調(diào)節(jié)腫瘤相關(guān)基因的表達(dá),包括細(xì)胞增殖、凋亡、細(xì)胞周期阻滯、血管形成和轉(zhuǎn)移等基因,促進(jìn)腫瘤的發(fā)生發(fā)展[11-13]。miRNA在腫瘤耐藥過(guò)程中發(fā)揮的作用近年來(lái)倍受關(guān)注,一些miRNA可能通過(guò)其靶基因參與化學(xué)藥物耐藥性的產(chǎn)生[14,15]。miRNA在5-Fu耐藥過(guò)程中的作用及機(jī)制尚不清楚。
蛋白磷酸酶2A(protein phosphatase 2A,PP2A)是真核細(xì)胞內(nèi)廣泛表達(dá)的絲氨酸/蘇氨酸蛋白磷酸酶家族的主要成員,在去磷酸化底物分子及調(diào)節(jié)大多數(shù)細(xì)胞事件和生物過(guò)程中具有關(guān)鍵性作用。PP2A是由不同亞基組成的結(jié)構(gòu)復(fù)合體,其在細(xì)胞內(nèi)生理功能的發(fā)揮有賴于各亞基的正常轉(zhuǎn)錄和表達(dá)[16]。其中,Aβ被認(rèn)為是核心亞基,該亞基由蛋白質(zhì)磷酸酶2調(diào)節(jié)亞基1B(PPP2R1B)基因編碼。PPP2R1B基因共有15個(gè)外顯子,總長(zhǎng)度約有27 kb,位于染色體11q23區(qū)域,該區(qū)域是癌細(xì)胞常缺失的一段區(qū)域[17,18],研究報(bào)道PPP2R1B基因在結(jié)直腸癌及其他多種癌組織樣本中表達(dá)下降[19,20]。體外實(shí)驗(yàn)發(fā)現(xiàn)PPP2R1B的表達(dá)改變可顯著影響PP2A對(duì)Akt信號(hào)通路的去磷酸化作用,進(jìn)而影響癌細(xì)胞對(duì)多種化療藥物的敏感性[21,22]。
本研究用5-Fu對(duì)人結(jié)腸癌細(xì)胞株進(jìn)行誘導(dǎo),建立結(jié)腸癌耐藥細(xì)胞株,探討其生物學(xué)特性,并篩選耐藥細(xì)胞和親本細(xì)胞中差異表達(dá)的miRNA分子及可能涉及的分子調(diào)控機(jī)制,為尋找耐藥逆轉(zhuǎn)劑的研究提供實(shí)驗(yàn)依據(jù)。
1材料與方法
1.1主要儀器與試劑
Eppendorf 5810R型臺(tái)式大容量高速冷凍離心機(jī)(德國(guó)Eppendorf公司),Nikon Eclipse Ti-E型倒置顯微鏡(日本Nikon公司),AppliedBiosystems7500實(shí)時(shí)PCR儀(美國(guó)ABI公司),凝膠成像系統(tǒng)和FACS Calibur流式細(xì)胞儀(美國(guó)BD公司),倒置熒光顯微鏡(德國(guó)Leica),雙熒光素酶分析系統(tǒng)(美國(guó)Promega公司)。
人結(jié)腸癌細(xì)胞株COLO320,Caco-2,RKO,HCT-116,HCT-29(美國(guó)American Types Culture Collection,ATCC),小牛血清和RPMI1640培養(yǎng)基(美國(guó)Gibco公司),TRizol試劑,Lipofectamin2000和Annexin Ⅴ/PI凋亡試劑盒(美國(guó)Invitrogen公司),miRNA表達(dá)芯片Human Apoptosis miRNA PCR Array:MIHS-114Z(德國(guó)Qiagen公司),miRNA提取試劑盒(美國(guó)Roche公司),miRNA抑制劑和模擬物(上海吉瑪公司),miRNA-141引物(上海生工),四甲基偶氮唑鹽(methylthiazolyl tetrazolium,MTT)和結(jié)晶紫染色試劑序列(美國(guó)Sigma公司),RT逆轉(zhuǎn)錄試劑盒(大連寶生物),蛋白質(zhì)磷酸酶2調(diào)節(jié)亞基1B(PPP2R1B)抗體,β-actin,蛋白激酶B(AKT)和磷酸化的蛋白激酶B(Phospho-AKT)抗體(美國(guó)Cell Signaling公司)。
1.2細(xì)胞培養(yǎng)及耐藥細(xì)胞系的建立
人結(jié)腸癌細(xì)胞株加入濃度為10%小牛血清的RPMI1640完全培養(yǎng)基,37 ℃、5%CO2孵箱培養(yǎng)。根據(jù)以往的結(jié)腸癌細(xì)胞特性的研究結(jié)果以及本次實(shí)驗(yàn)的藥物敏感性測(cè)試結(jié)果,相對(duì)于其他結(jié)腸癌細(xì)胞系,COLO320對(duì)5-Fu最為敏感。因此,本實(shí)驗(yàn)采取大劑量沖擊聯(lián)合劑量遞增的方法建立5-Fu耐藥結(jié)腸癌耐藥細(xì)胞株COLO320-R。先用50 mg/L 5-Fu的培養(yǎng)液作用約24 h,觀察有70%細(xì)胞死亡時(shí),停止藥物作用,PBS緩沖液沖洗2次,此時(shí)僅有10%-20%細(xì)胞貼壁生長(zhǎng),用含100 ml/L胎牛血清的RPMI1640培養(yǎng)液持續(xù)培養(yǎng)至細(xì)胞鋪滿瓶底80%,然后在培養(yǎng)液中加入初始劑量為0.1 μmol/L的5-Fu,細(xì)胞在此劑量下培養(yǎng)3 d,然后在不含5-Fu的細(xì)胞培養(yǎng)液中達(dá)到70%生長(zhǎng)。待細(xì)胞進(jìn)入對(duì)數(shù)生長(zhǎng)期時(shí),分瓶傳代,以梯度增加方式,給予2.0倍濃度的5-Fu繼續(xù)培養(yǎng),按照此方式逐漸增加劑量,直至細(xì)胞可以在含5.0 μmol/L 5-Fu的培養(yǎng)基中正常生長(zhǎng),將此耐藥細(xì)胞系命名為COLO320/5-Fu。
1.3四甲基偶氮唑鹽(MTT)測(cè)試細(xì)胞IC50
用MTT法檢測(cè)比較耐藥細(xì)胞和親代細(xì)胞對(duì)5-Fu的敏感性。取對(duì)數(shù)生長(zhǎng)期的細(xì)胞,用2.5 g/L胰酶消化,RPMI1640培養(yǎng)液制成單細(xì)胞懸液,接種到96孔板(100 μl/孔,5×103/孔),培養(yǎng)24 h,加入不同濃度的5-Fu(設(shè)4復(fù)孔和陰性對(duì)照),再培養(yǎng)72 h,每孔加入MTT液20 μl培養(yǎng)4 h,離心,棄上清,加入二甲亞砜100 μl/孔,震蕩搖勻,紫色顆粒溶解后上酶標(biāo)儀測(cè)定吸光度,波長(zhǎng)570 nm,以藥物濃度為橫軸,細(xì)胞存活率為縱軸,繪制濃度-效應(yīng)曲線,確定半數(shù)抑制濃度(IC50)。
1.4細(xì)胞克隆形成實(shí)驗(yàn)和凋亡實(shí)驗(yàn)
在集落形成測(cè)定中,1×103轉(zhuǎn)染的細(xì)胞接種到6孔板,每孔用含不同濃度的5-Fu處理。每3 d替換1次含有5-Fu的培養(yǎng)基,直到形成可見(jiàn)菌落。菌落用4%多聚甲醛固定并用0.1%結(jié)晶紫染色。對(duì)菌落直徑>50 μm的進(jìn)行計(jì)數(shù)。流式細(xì)胞儀檢測(cè)細(xì)胞凋亡率:用胰酶消化法收集細(xì)胞,預(yù)冷的PBS洗2次,Annexin Ⅴ室溫下避光染色30 min,上流式細(xì)胞儀檢測(cè)。每組實(shí)驗(yàn)重復(fù)3次。WinMDI2.8軟件分析AnnexinⅤ-FITC單染細(xì)胞所占百分率即細(xì)胞凋亡率。
1.5miRNA拮抗劑轉(zhuǎn)染細(xì)胞
耐藥細(xì)胞和對(duì)照細(xì)胞在培養(yǎng)皿中生長(zhǎng)至70%覆蓋培養(yǎng)皿。然后按照廠商說(shuō)明用siRORT轉(zhuǎn)染試劑將5 nmol/L的miR-141拮抗劑或陰性對(duì)照分別轉(zhuǎn)染入細(xì)胞。細(xì)胞繼續(xù)培養(yǎng)72 h,然后收集細(xì)胞,RT-PCR測(cè)定轉(zhuǎn)染效率,并進(jìn)行細(xì)胞功能學(xué)測(cè)試。
1.6熒光報(bào)告基因?qū)嶒?yàn)
用Lipofectamine2000構(gòu)建包含PPP2R1B野生和突變3′-UTR結(jié)合區(qū)域序列的熒光素酶報(bào)告構(gòu)建體。耐藥細(xì)胞以約1.5×104的密度接種到96孔板/孔,然后和共轉(zhuǎn)染的miRNA-141模擬物以及熒光素酶報(bào)告構(gòu)建體共轉(zhuǎn)染,將細(xì)胞在裂解緩沖液轉(zhuǎn)染后48 h裂解。使用雙熒光素酶分析系統(tǒng)的海腎和螢火蟲(chóng)熒光素酶活性進(jìn)行定量。
1.7基于PCR的miRNA微陣列及miRNA的RT-PCR逆轉(zhuǎn)錄實(shí)驗(yàn)
分別從親代細(xì)胞和耐藥細(xì)胞中提取RNA,采用Human Apoptosis miRNA PCR Array:MIHS-114Z 試劑盒利用qRT-PCR方法測(cè)試差異miRNA表達(dá)差異。RNU48作為內(nèi)參基因計(jì)算標(biāo)準(zhǔn)化Ct值。耐藥細(xì)胞的每個(gè)miRNA值與親代對(duì)照細(xì)胞比較得到差異表達(dá)率。
定量PCR引物序列如下:miR-141Forward:5′-GCGAAAGCATTTGCCAAGAA-3′;Reverse:5′-CAATCACAGACCTGTTATTGC-3′,以RNU48作為內(nèi)參。定量PCR條件如下:95 ℃預(yù)變性2 min,94 ℃變性1 min,60 ℃退火1 min,72 ℃延伸1 min,循環(huán)40次;5 ℃保溫30 s,行熔解曲線分析。miRNA的相對(duì)含量根據(jù)2-ΔΔCt計(jì)算。
1.8蛋白印跡實(shí)驗(yàn)檢測(cè)PPP2R1B,Akt和磷酸化Akt蛋白表達(dá)水平
用蛋白裂解液于冰上裂解細(xì)胞30 min,4 ℃, 10 000 r/min,離心40 min,取上清-80 ℃儲(chǔ)存?zhèn)溆茫锨逵肂radford法測(cè)定蛋白濃度,樣品95 ℃變性5 min后,每孔上樣60 μg,SDS-PAGE(5%焦集膠、8%分離膠)電泳80 V 2 h轉(zhuǎn)膜于聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF) 膜上,用4%脫脂奶粉/TBST 室溫下封閉抗原1.5 h。一抗用4%脫脂奶粉/TBST稀釋后4℃孵育PVDF膜過(guò)夜(稀釋比例1∶400)。TBST洗膜3次,每次10 min,然后加入二抗,室溫下?lián)u床孵育1 h,再次TBST洗膜3次,每次10 min。ECL曝光顯影。
1.9統(tǒng)計(jì)學(xué)分析
應(yīng)用SPSS 13.0軟件進(jìn)行統(tǒng)計(jì)學(xué)處理,計(jì)量數(shù)據(jù)以均數(shù)±標(biāo)準(zhǔn)差表示,多組間均數(shù)比較采用單因素方差分析(One-way ANOVA),兩兩比較采用LSD法,雙側(cè)檢驗(yàn)。P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2結(jié)果
2.15-Fu耐藥細(xì)胞的建立和差異表達(dá)miRNAs
根據(jù)以往結(jié)腸癌細(xì)胞特性的研究結(jié)果以及本次實(shí)驗(yàn)的藥物敏感性測(cè)試結(jié)果,相對(duì)于其他結(jié)腸癌細(xì)胞系(Caco-2,RKO,HCT-116,HCT-29),COLO320對(duì)5-Fu最為敏感(P<0.05,見(jiàn)圖1A)。按照劑量逐步遞增法,本實(shí)驗(yàn)成功構(gòu)建了5-FU耐藥的結(jié)腸癌細(xì)胞株,耐藥細(xì)胞株的IC50比親代細(xì)胞增長(zhǎng)8.8倍[(9.50±0.47)μmol/Lvs(1.13±0.14)μmol/L,P<0.05,見(jiàn)圖1B]。基于PCR的miRNA微陣列結(jié)果顯示:與親代細(xì)胞比較,耐藥細(xì)胞中的7個(gè)miRNA表達(dá)增加大于1.5倍,6個(gè)表達(dá)改變減少小于1.5倍(見(jiàn)表1)。其中miRNA-141表達(dá)水平在兩組細(xì)胞系中變化最大。
2.2miR-141的表達(dá)抑制可以逆轉(zhuǎn)耐藥細(xì)胞耐藥性
為了進(jìn)一步研究miR-141對(duì)COLO-320/5-Fu細(xì)胞增殖和藥物敏感性的影響,給予COLO-320/5-Fu轉(zhuǎn)染miR-141抑制劑。轉(zhuǎn)染效果經(jīng)過(guò)RT-PCR驗(yàn)證(P<0.05,見(jiàn)圖2A)。MTT實(shí)驗(yàn)證實(shí)miR-141抑制劑轉(zhuǎn)染的細(xì)胞,5-Fu藥物敏感性與耐藥細(xì)胞相比明顯增加[(1.22±0.19)μmol/Lvs(9.47±0.53)μmol/L,P<0.05,見(jiàn)圖2B]。
表15-Fu耐藥細(xì)胞和親代細(xì)胞差異表達(dá)的miRNA
Table 1Differential miRNA profile expression in 5-Fu resistant cell line and parental cell line
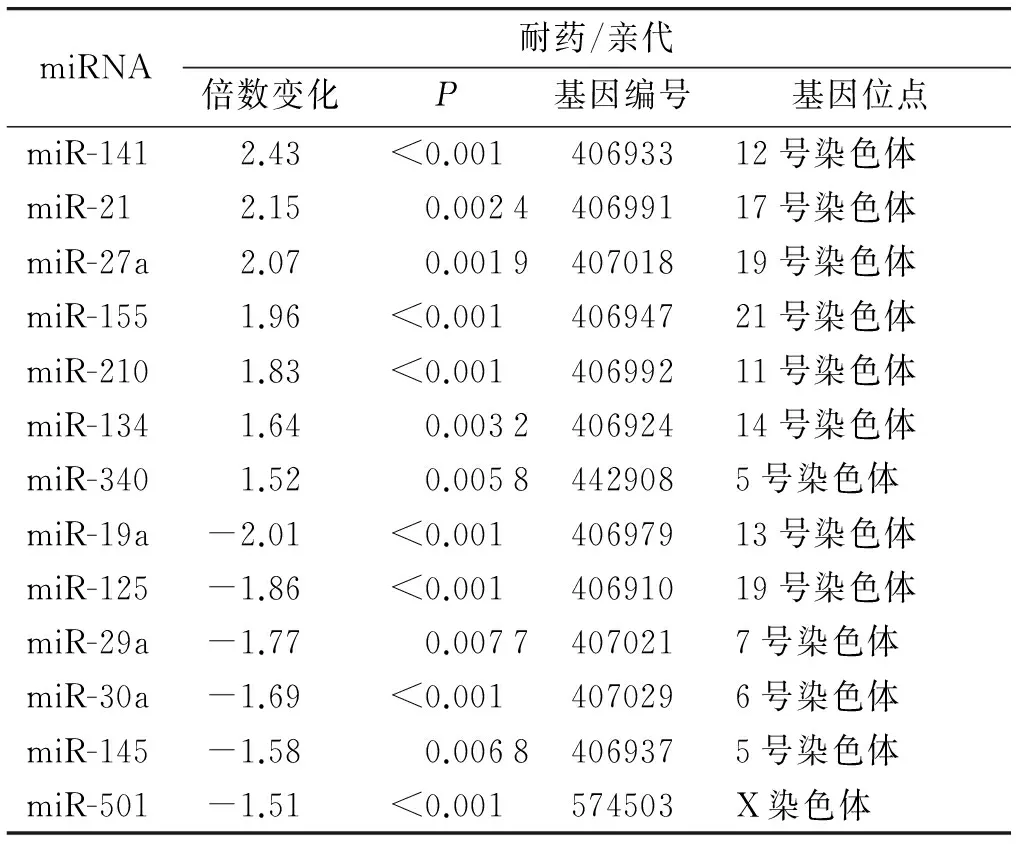
miRNA耐藥/親代倍數(shù)變化P基因編號(hào)基因位點(diǎn)miR-1412.43<0.00140693312號(hào)染色體miR-212.150.002440699117號(hào)染色體miR-27a2.070.001940701819號(hào)染色體miR-1551.96<0.00140694721號(hào)染色體miR-2101.83<0.00140699211號(hào)染色體miR-1341.640.003240692414號(hào)染色體miR-3401.520.00584429085號(hào)染色體miR-19a-2.01<0.00140697913號(hào)染色體miR-125-1.86<0.00140691019號(hào)染色體miR-29a-1.770.00774070217號(hào)染色體miR-30a-1.69<0.0014070296號(hào)染色體miR-145-1.580.00684069375號(hào)染色體miR-501-1.51<0.001574503X染色體
2.3miRNA-141影響細(xì)胞凋亡并通過(guò)靶向調(diào)控PPP2R1B導(dǎo)致5-Fu耐藥
在接受同樣劑量5-Fu處理后,轉(zhuǎn)染miR-141抑制劑的癌細(xì)胞的克隆形成與耐藥細(xì)胞相比明顯下降(23.44±1.97vs218.12±23.46,P<0.05,見(jiàn)圖3A,B)。進(jìn)一步細(xì)胞凋亡實(shí)驗(yàn)發(fā)現(xiàn),接受同樣劑量的5-Fu,轉(zhuǎn)染miR-141抑制劑的COLO-320/5-Fu細(xì)胞的凋亡變化比率與耐藥細(xì)胞相比明顯增加(3.75±0.26vs1.17±0.24,P<0.05,見(jiàn)圖3C)。
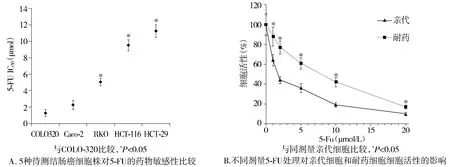
圖1 不同結(jié)腸癌細(xì)胞株5-Fu藥物敏感性測(cè)試及5-Fu耐藥細(xì)胞株的驗(yàn)證Figure 1 Results of drug sensitivity to 5-Fu in different colon cancer cell lines and verification of 5-Fu-resistance cell
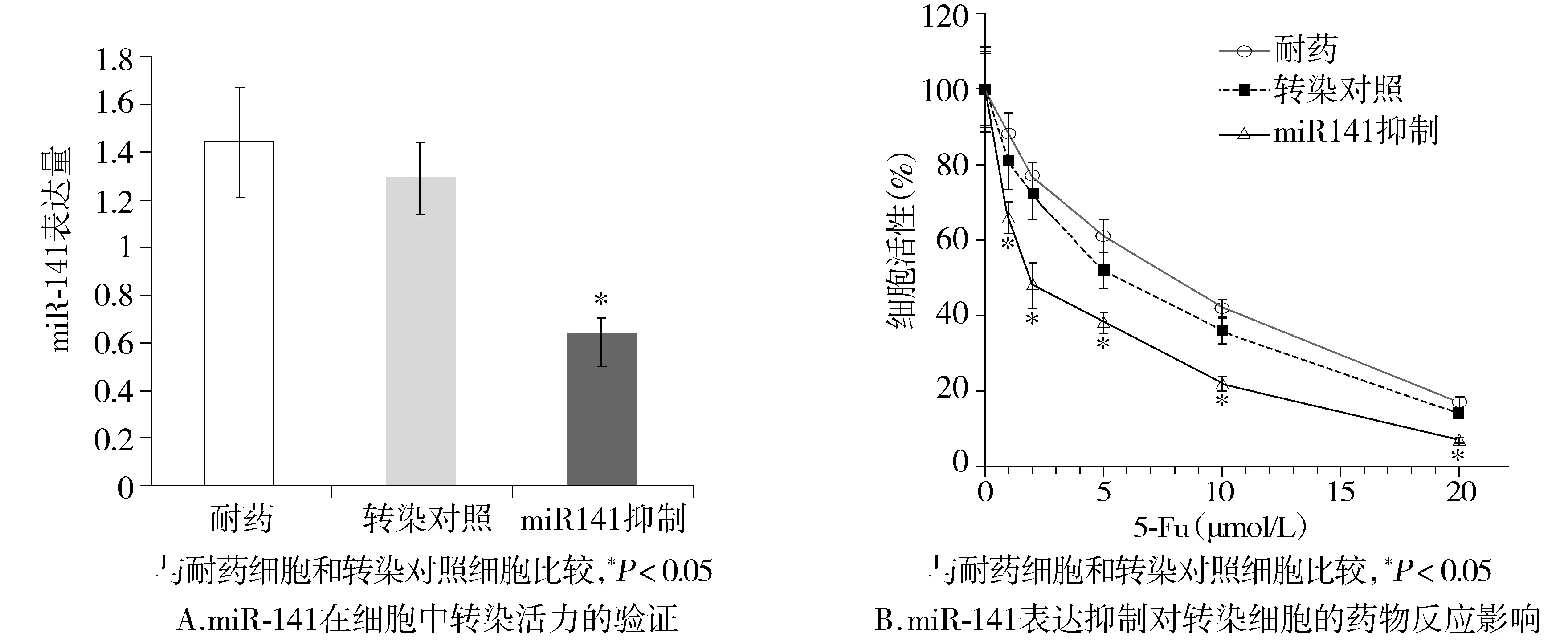
圖2 miR-141表達(dá)抑制對(duì)5-Fu耐藥性的影響Figure 2 Influence of miR-141 inhibition on sensitivity to 5-Fu
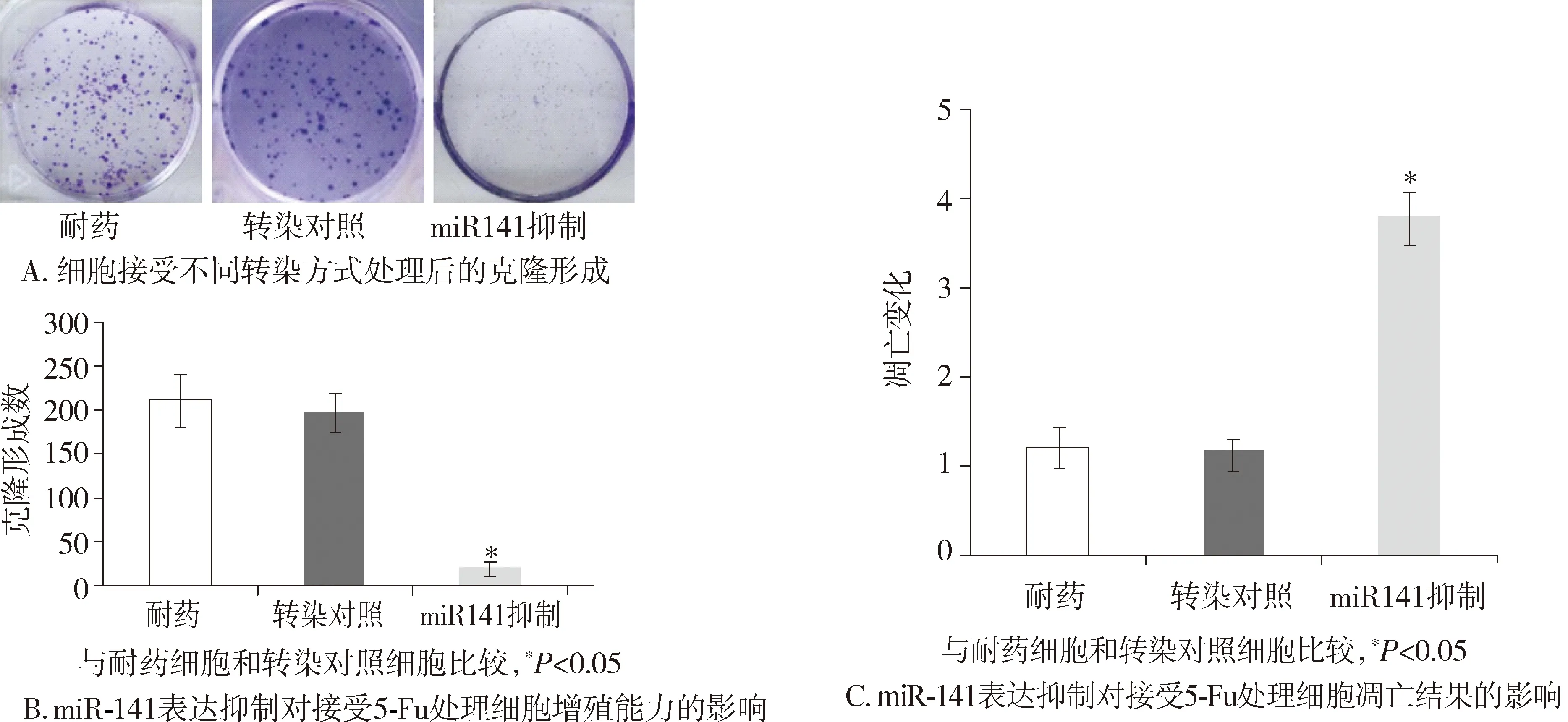
圖3 miR-141表達(dá)抑制對(duì)細(xì)胞功能的影響Figure 3 Influence of down-regulation of miRNA-141 on cell proliferation and apoptosis
2.4miRNA-141靶向調(diào)控PPP2R1B導(dǎo)致細(xì)胞對(duì)5-Fu耐藥
為進(jìn)一步研究miRNA-141導(dǎo)致結(jié)腸癌細(xì)胞產(chǎn)生5-Fu耐藥性的調(diào)控機(jī)制,本研究使用Targetscan和miRBase database等軟件進(jìn)行潛在靶基因預(yù)測(cè),基于生物信息學(xué)分析結(jié)果,篩選出Akt通路亞基的PPP2R1B為可能的目標(biāo)基因。為驗(yàn)證miR-141對(duì)PPP2R1B的直接調(diào)控作用,本研究將PPP2R1B啟動(dòng)區(qū)與miR-141結(jié)合序列中的部分堿基突變(見(jiàn)圖4A), 以包含野生PPP2R1B啟動(dòng)區(qū)序列(PPP2R1B野生型)和突變PPP2R1B啟動(dòng)區(qū)序列(PPP2R1B突變型)的熒光酶報(bào)告載體與miR-141抑制劑共同轉(zhuǎn)染耐藥細(xì)胞,以不含PPP2R1B啟動(dòng)區(qū)的質(zhì)粒作為對(duì)照(空白載體),結(jié)果顯示,與轉(zhuǎn)染試劑空白對(duì)照和陰性轉(zhuǎn)染對(duì)照相比,miR-141抑制劑可以顯著增加耐藥細(xì)胞中包含野生PPP2R1B啟動(dòng)區(qū)序列的熒光酶活力(1.32±0.08,P<0.05),但對(duì)包含突變PPP2R1B啟動(dòng)區(qū)序列和空白載體的熒光酶活力沒(méi)有影響(1.03±0.09vs0.97±0.09,P>0.05,見(jiàn)圖4B)。由于PPP2R1B主要通過(guò)調(diào)節(jié)Akt去磷酸化過(guò)程影響細(xì)胞功能,通過(guò)蛋白印跡實(shí)驗(yàn)進(jìn)一步研究敲除miR-141對(duì)PPP2R1B及Akt去磷酸化過(guò)程的影響,結(jié)果發(fā)現(xiàn)miR-141抑制劑可導(dǎo)致PPP2R1B蛋白表達(dá)量增加(1.39±0.09),同時(shí)引起磷酸化的Akt表達(dá)量下降(0.67±0.05)(P<0.05,見(jiàn)圖4C,D)。
3討論
手術(shù)是早期結(jié)腸癌主要的治療手段,以5-Fu為基礎(chǔ)的化療則是術(shù)后輔助化療和中晚期結(jié)腸癌患者的標(biāo)準(zhǔn)治療方案。盡管目前5-Fu廣泛應(yīng)用于結(jié)腸癌患者,但部分患者經(jīng)化療后仍然出現(xiàn)腫瘤復(fù)發(fā)和轉(zhuǎn)移,這可能與患者對(duì)5-Fu產(chǎn)生原發(fā)性或獲得性耐藥有關(guān)。因此,尋找影響5-Fu藥物敏感性的生物學(xué)分子對(duì)提高結(jié)腸癌的療效具有重要意義。本研究發(fā)現(xiàn)新的miR-141/PPP2R1B/AKT/信號(hào)調(diào)節(jié)通路可以影響結(jié)腸癌細(xì)胞對(duì)5-Fu治療的反應(yīng)性。miR-141抑制劑可以增加結(jié)腸癌細(xì)胞對(duì)5-Fu的治療敏感性,其可能機(jī)制為抑制miR-141可以促進(jìn)PPP2R1B的表達(dá),導(dǎo)致AKT活化增強(qiáng),進(jìn)而促進(jìn)結(jié)腸癌細(xì)胞對(duì)5-Fu的敏感性。另一方面,抑制miR-141可以促進(jìn)5-Fu耐藥細(xì)胞的凋亡并降低其增殖能力。
本研究通過(guò)比較結(jié)腸癌親代細(xì)胞和5-Fu耐藥細(xì)胞(表1)之間的miRNA表達(dá)譜,發(fā)現(xiàn)在5-Fu耐藥細(xì)胞中miR-21、miR-27A和miR-155表達(dá)顯著增高,miR-340、miR-145和miR-451的表達(dá)顯著降低,有研究表明這些高表達(dá)的miRNA均表現(xiàn)出癌基因的性質(zhì)[23-25],而表達(dá)降低的miRNA則具有腫瘤抑制作用[24]。研究證實(shí)miRNA在許多惡性腫瘤的發(fā)生發(fā)展中起重要作用,miRNA與腫瘤關(guān)系的研究日益受到關(guān)注。由于miRNA的轉(zhuǎn)錄后調(diào)節(jié)功能,其表達(dá)的改變會(huì)使多個(gè)關(guān)鍵的基因和通路受到影響。許多miRNA在結(jié)腸癌和5-Fu耐藥過(guò)程中發(fā)揮著重要作用[27-30]。研究顯示miR-192直接參與結(jié)直腸癌氟尿嘧啶和抗葉酸化療耐藥的過(guò)程[28]。Schetter等[29]指出miR-21可能與氟尿嘧啶類化療藥物的治療反應(yīng)相關(guān),該研究分析了84例結(jié)腸腺癌組織及其配對(duì)的癌旁正常組織中miRNA的表達(dá)譜,發(fā)現(xiàn)miR-20a、miR-21、miR-106a、miR-181b和miR-203存在明顯的異常表達(dá),應(yīng)用定量PCR技術(shù)在驗(yàn)證組中進(jìn)一步分析這些miRNA的表達(dá)水平與腫瘤分期、預(yù)后以及對(duì)化療反應(yīng)之間的關(guān)系,發(fā)現(xiàn)在接受基于氟尿嘧啶類方案輔助化療的Ⅱ期和Ⅲ期結(jié)腸腺癌患者中,miR-21高表達(dá)患者對(duì)氟尿嘧啶類化療藥物的反應(yīng)較差,并與不良療效相關(guān)。體外實(shí)驗(yàn)發(fā)現(xiàn),在p53野生型的HCT116細(xì)胞系中過(guò)表達(dá)的miR-140作用于組蛋白去乙酰化酶4可導(dǎo)致5-Fu的耐藥[30],進(jìn)一步研究證實(shí),結(jié)腸癌干細(xì)胞樣細(xì)胞中的內(nèi)源性miR-140表達(dá)上調(diào)可使這些細(xì)胞表現(xiàn)出化療抗拒性;而利用小干擾RNA(small interfering RNA,siRNA)技術(shù)抑制結(jié)腸癌干細(xì)胞中miR-140的表達(dá),可明顯提高HCT116細(xì)胞對(duì)5-Fu的敏感性。這些研究表明,結(jié)腸癌患者接受5-Fu相關(guān)化療方案的治療效果與某些miRNA的異常表達(dá)相關(guān)。Akao等[31]發(fā)現(xiàn),miR-34a在5-Fu耐藥的結(jié)腸癌DLD-1/5-Fu細(xì)胞中的表達(dá)低于DLD-1細(xì)胞;DLD-1/5-Fu細(xì)胞經(jīng)5-Fu處理后,miR-34a持續(xù)低表達(dá)。DLD-1細(xì)胞經(jīng)5-Fu處理后,miR-34a的表達(dá)水平顯著升高。進(jìn)一步研究發(fā)現(xiàn),DLD-1/5-Fu細(xì)胞中的Sirt1(sirtuin1)表達(dá)水平顯著高于對(duì)照組。Sirt1是一種高度保守的基因和細(xì)胞凋亡負(fù)性調(diào)節(jié)因子,可編碼煙酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD)依賴的脫乙酰酶,并與化療耐藥相關(guān)。Sirt1是miR-34a的靶基因之一,因此miR-34a低表達(dá)可導(dǎo)致Sirt1高表達(dá),從而降低DLD-1/5-Fu細(xì)胞對(duì)5-Fu的敏感性。本研究發(fā)現(xiàn)miR-141在5-Fu耐藥結(jié)腸癌細(xì)胞中高表達(dá)。有研究報(bào)道結(jié)腸癌血清miR-141水平與臨床分級(jí)和預(yù)后密切相關(guān),高表達(dá)的miR-141血清水平可以作為臨床較差分級(jí)和不良預(yù)后的潛在標(biāo)志物[32]。體外實(shí)驗(yàn)和動(dòng)物實(shí)驗(yàn)也發(fā)現(xiàn),miR-141表達(dá)抑制可以降低胰腺癌細(xì)胞的增殖活性和侵襲性[33]。也有研究發(fā)現(xiàn)miR-141通過(guò)靶向調(diào)節(jié)KEAP1影響卵巢癌對(duì)順鉑的藥物敏感性[34]。結(jié)合本研究結(jié)果,提示miR-141可能通過(guò)發(fā)揮類似促癌基因的作用影響細(xì)胞對(duì)化療藥物的耐藥性。
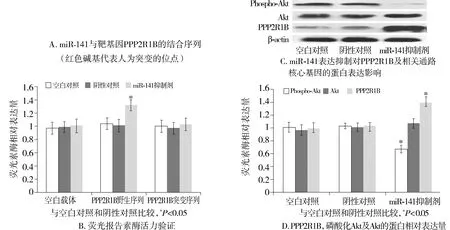
圖4 miR-141與靶向基因PPP2R1B的結(jié)合及對(duì)Akt磷酸化的影響Figure 4 Binding effect of miRNA-141 on PPP2R1B and Akt phosphorylation
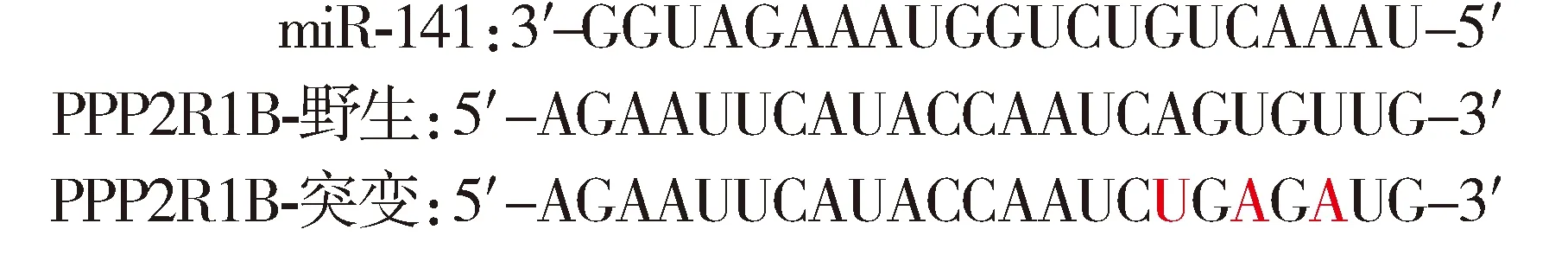
Akt是影響結(jié)腸癌細(xì)胞存活的一個(gè)重要因子,PP2A可以通過(guò)對(duì)Akt的兩個(gè)主要活性基團(tuán)位點(diǎn)蘇氨酸308(Thr308)和絲氨酸473(Ser473)進(jìn)行去磷酸化從而影響Akt的活性[16]。PPP2R1B是PP2A的一個(gè)重要組成亞基,可以通過(guò)二聚體形式調(diào)節(jié)PP2A的表達(dá)[17]。研究發(fā)現(xiàn)PPP2R1B對(duì)結(jié)腸癌細(xì)胞具有抑制作用,約有15%的原發(fā)性結(jié)腸腫瘤中出現(xiàn)PPP2R1B基因的缺失或突變,從而降低了的PPP2R1B基因的表達(dá),進(jìn)而減弱了PPP2R1B的腫瘤抑制作用[19,20]。本研究發(fā)現(xiàn)抑制miR-141可促進(jìn)PPP2R1B和磷酸化Akt的表達(dá),表明miRNA介導(dǎo)的轉(zhuǎn)錄后調(diào)控可能是一種調(diào)控結(jié)腸癌細(xì)胞PPP2R1B表達(dá)的新機(jī)制。由于癌細(xì)胞常通過(guò)多種機(jī)制減少抑癌基因的表達(dá),因此恢復(fù)抑癌基因的表達(dá)活性,從小分子RNA的角度設(shè)計(jì)治療策略成為治療結(jié)腸癌患者的新思路。有研究證實(shí)Akt信號(hào)通路可以通過(guò)調(diào)節(jié)核轉(zhuǎn)錄因子(NF-κB),環(huán)磷酸腺苷反應(yīng)元件結(jié)合蛋白(cAMP response element binding protein, CREB)以及Bcl-2相關(guān)蛋白的作用影響細(xì)胞凋亡[35,36],而本研究的凋亡和增殖實(shí)驗(yàn)觀察到miR-141表達(dá)抑制的細(xì)胞,增殖能力減弱,同時(shí)凋亡比例增加,說(shuō)明小分子RNA對(duì)腫瘤的發(fā)生發(fā)展有重大作用。本實(shí)驗(yàn)結(jié)果表明,結(jié)腸癌細(xì)胞系中抗凋亡效應(yīng)是miR-141介導(dǎo)的5-Fu的化療抗拒性的關(guān)鍵機(jī)制。由于5-Fu可治療多種惡性腫瘤,抑制miR-141的表達(dá)能否提高5-Fu治療其他腫瘤的臨床療效需要進(jìn)一步的研究。
本實(shí)驗(yàn)研究確定了miR-141通過(guò)調(diào)節(jié)Akt通路途徑參與5-Fu耐藥機(jī)制。miR-141表達(dá)抑制可誘導(dǎo)結(jié)腸癌細(xì)胞凋亡并提高細(xì)胞對(duì)5-Fu的藥物敏感性。因此,miR-141可以為治療結(jié)腸癌藥物的耐藥反應(yīng)的處理提供潛在的治療靶標(biāo),為改善患者的臨床治療效果提供新的思路。
參考文獻(xiàn):
[1]Zhang Y,Talmon G,Wang J.MicroRNA-587 antagonizes 5-Fu-induced apoptosis and confers drug resistance by regulating PPP2R1B expression in colorectal cancer[J].Cell Death Dis,2015,6:e1845.
[2]Longley DB,Harkin DP,Johnston PG.5-fluorouracil:mechanisms of action and clinical strategies[J].Nat Rev Cancer,2003,3(5):330-338.
[3]Longley DB,Allen WL,Johnston PG.Drug resistance,predictive markers and pharmacogenomics in colorectal cancer[J].Biochim Biophys Acta,2006,1766(2):184-196.
[4]Liu YY,Han TY,Giuliano AE,etal.Ceramide glycosylation potentiates cellular multidrug resistance[J].FASEB J,2001,15(3):719-730.
[5]Longley DB,Johnston PG.Molecular mechanisms of drug resistance[J].J Pathol,2005,205(2):275-292.
[6]Synold TW,Dussault I,Forman BM.The orphan nuclear receptor SXR coordinately regulates drug metabolism and efflux[J].Nat Med,2001,7(5):584-590.
[7]Gottesman MM.Mechanisms of cancer drug resistance[J].Annu Rev Med,2002,53:615-627.
[8]Bartel DP.MicroRNAs:target recognition and regulatory functions[J].Cell,2009,136(2):215-233.
[9]Lewis BP,Burge CB,Bartel DP.Conserved seed pairing,often flanked by adenosines,indicates that thousands of human genes are microRNA targets[J].Cell,2005,120(1):15-20.
[10]Lujambio A,Lowe SW.The microcosmos of cancer[J].Nature,2012,482(7385):347-355.
[11]Calin GA,Sevignani C,Dumitru CD,etal.Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers[J].Proc Natl Acad Sci USA,2004,101(9):2999-3004.
[12]Zhang B,Pan X,Cobb GP,etal.microRNAs as oncogenes and tumor suppressors[J].Dev Biol,2007,302(1):1-12.
[13]Garzon R,Marcucci G,Croce CM.Targeting microRNAs in cancer:rationale,strategies and challenges[J].Nat Rev Drug Discov,2010,9(10):775-789.
[14]Garofalo M,Croce CM.MicroRNAs as therapeutic targets in chemoresistance[J].Drug Resist Updat,2013,16(3-5):47-59.
[15]Donzelli S,Mori F,Biagioni F,etal.MicroRNAs:short non-coding players in cancer chemoresistance[J].Mol Cell Ther,2014,2:16.
[16]Schonthal AH.Role of serine/threonine protein phosphatase 2A in cancer[J].Cancer Lett,2001,170(1):1-13.
[17]Sablina AA,Chen W,Arroyo JD,etal.The tumor suppressor PP2A Abeta regulates the RalA GTPase[J].Cell,2007,129(5):969-982.[18]Poretti G,Kwee I,Bernasconi B,etal.Chromosome 11q23.1 is an unstable region in B-cell tumor cell lines[J].Leuk Res,2011,35(6):808-813.
[19]Takagi Y,Futamura M,Yamaguchi K,etal.Alterations of the PPP2R1B gene located at 11q23 in human colorectal cancers[J].Gut,2000,47(2):268-271.[20]Wang SS,Esplin ED,Li JL,etal.Alterations of the PPP2R1B gene in human lung and colon cancer[J].Science,1998,282(5387):284-287.[21]Koti M,Gooding RJ,Nuin P,etal.Identification of the IGF1/PI3K/NF kappaB/ERK gene signalling networks associated with chemotherapy resistance and treatment response in high-grade serous epithelial ovarian cancer[J].BMC Cancer, 2013,13:549.
[22]Hamano R,Miyata H,Yamasaki M,etal.Overexpression of miR-200c induces chemoresistance in esophageal cancers mediated through activation of the Akt signaling pathway[J].Clin Cancer Res,2011,17(9):3029-3038.
[23]Ma X,Choudhury SN,Hua X,etal.Interaction of the oncogenic miR-21 microRNA and the p53 tumor suppressor pathway[J].Carcinogenesis,2013,34(6):1216-1223.
[24]Wu XJ,Li Y,Liu D,etal.miR-27a as an oncogenic microRNA of hepatitis B virus-related hepatocellular carcinoma[J].Asian Pac J Cancer Prev,2013,14(2):885-889.
[25]Rather MI,Nagashri MN,Swamy SS,etal.Oncogenic microRNA-155 down-regulates tumor suppressor CDC73 and promotes oral squamous cell carcinoma cell proliferation:implications for cancer therapeutics[J].J Biol Chem,2013,288(1):608-618.
[26]Gits CM,van Kuijk PF,Jonkers MB,etal.MicroRNA expression profiles distinguish liposarcoma subtypes and implicate miR-145 and miR-451 as tumor suppressors[J].Int J Cancer,2014,135(2):348-361.
[27]Slaby O,Svoboda M,Michalek J,etal.MicroRNAs in colorectal cancer: translation of molecular biology into clinical application[J].Mol Cancer,2009,8:102.
[28]Song B,Wang Y,Kudo K,etal.miR-192 Regulates dihydrofolate reductase and cellular proliferation through the p53-microRNA circuit[J].Clin Cancer Res, 2008,14(24):8080-8086.
[29]Schetter AJ,Leung SY,Sohn JJ,etal.MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma[J].JAMA,2008,299(4):425-436.
[30]Song B,Wang Y,Xi Y,etal.Mechanism of chemoresistance mediated by miR-140 in human osteosarcoma and colon cancer cells[J].Oncogene,2009,28(46):4065-4074.
[31]Akao Y,Noguchi S,Iio A,etal.Dysregulation of microRNA-34a expression causes drug-resistance to 5-Fu in human colon cancer DLD-1 cells[J].Cancer Lett,2011,300(2):197-204.
[32]Cheng H,Zhang L,Cogdell DE,etal.Circulating plasma MiR-141 is a novel biomarker for metastatic colon cancer and predicts poor prognosis[J]. PLoS One,2011,6(3):e17745.
[33]Zhao G,Wang B,Liu Y,etal.miRNA-141,downregulated in pancreatic cancer,inhibits cell proliferation and invasion by directly targeting MAP4P4[J].Mol Cancer Ther,2013,12(11): 2569-2580.[34]van Jaarsveld MT,Helleman J,Boersma AW,etal.miR-141 regulates KEAP1 and modulates cisplatin sensitivity in ovarian cancer cells[J].Oncogene,2013,32(36):4284-4293.
[35]Koti M,Gooding RJ,Nuin P,etal.Identification of the IGF1/PI3K/NF-kB/ERK gene signaling networks associated with chemotherapy resistance and treatment response in high-grade serous epithelial ovarian cancer[J].BMC Cancer,213,13:549.
[36]Sakamaki J,Daitoku H,Ueno K,etal.Arginine methylation of BCL-2 antagonist of cell death(BAD) counteracts its phosphorylation and inactivation by Akt[J].Proc Natl Sci USA,2011,108(15):6085-6090.
Inhibition of miR-141 increases sensitivity of colon cancer cells to 5-Fu
MA Yinan1, JIN Yingying1*,WANG Yali1, CHEN Qingjuan2, WEI Yang3
(1DepartmentofOncology,SecondAffiliatedHospitalofXi’anJiaotongUniversity,Xi’an710004,China;2DepartmentofOncology,XianyangCenterHospital;3ScientificResearchCenter,SecondAffiliatedHospitalofXi’anJiaotongUniversity;*Correspondingauthor,E-mail:yingyingjin717@sina.com)
Abstract:ObjectiveTo establish a 5-fluorouracil(5-Fu)-resistant human colon cancer cell line and to explore the mechanism of resistance of miRNA-141 interaction with PPP2R1B to 5-Fu.MethodsBased on the results of drug sensitivity test, the 5-Fu sensitive cell line COLO-320 was initially given a high dose of 5-Fu followed by a stepwise treatment to establish a 5-Fu-resistant colon cancer cell line. IC50of 5-Fu-resistant cell line and parental cell line were calculated with MTT. PCR-based microarray technology was used to compare the differential expression of miRNAs between 5-Fu-resistant cells and the parental cells. The potential target gene of miR-141 was predicted with Targetscan, miRBase database. Furthermore, the predicated target gene was validated with a luciferase analysis. The miR-141 knockdown experiments were performed to investigate the binding effect of miR-141 on PPP2R1B.ResultsAn 5-Fu resistance colon cell line(COLO-320/5-Fu) was acquired at a final concentration of 5.0 μmol/L following the stepwise drug increments. The IC50of resistant cell line was significantly higher than that of parental cells(P<0.05). MicroRNA panel results indicated that a total of 13 miRNAs were differentially expressed, and miR-141 was highly overexpressed in resistant cells(P<0.05). After knockdown of miRNA-141 expression, the sensitivity of resistant cell to 5-Fu and the percentage of apoptosis increased(P<0.05). The luciferase result revealed that PPP2R1B was a target gene of miR-141. The inhibition of miRNA-141 significantly increased the expression level of PPP2R1B and consequently disturbed the Akt phosphorylation process.ConclusionThe COLO-320/5-Fu-resistant cell line is successfully established. The miRNA-141 may regulate the resistance of colon cancer cell to 5-Fu through targeted regulation of PPP2R1B.
Key words:colon cancer;5-fluorouracil;PPP2R1B;drug resistance;microRNA
作者簡(jiǎn)介:馬一楠,女,1985-10生,碩士,住院醫(yī)師,E-mail:2251990377@qq.com
收稿日期:2016-03-28
中圖分類號(hào):R735.35
文獻(xiàn)標(biāo)志碼:A
文章編號(hào):1007-6611(2016)06-0510-08
DOI:10.13753/j.issn.1007-6611.2016.06.006
猜你喜歡
小獼猴智力畫(huà)刊(2022年9期)2022-11-04 02:31:42
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
西南國(guó)防醫(yī)藥(2016年7期)2016-12-01 06:01:15
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年1期)2016-01-14 03:41:26
中國(guó)當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
河南醫(yī)學(xué)研究(2014年3期)2014-02-27 14:51:48
