水稻株高、穗長和每穗穎花數的遺傳研究
2016-06-21 05:45:20方兆偉劉金波王寶祥宋兆強盧百關周振玲秦德榮徐大勇
江西農業學報 2016年5期
關鍵詞:水稻
方兆偉,劉金波,王寶祥,遲 銘,宋兆強,李 健,楊 波,盧百關,劉 艷,周振玲,秦德榮,徐大勇
(江蘇省連云港市農業科學院/江蘇省現代作物生產協同創新中心,江蘇 連云港 222006)
?
水稻株高、穗長和每穗穎花數的遺傳研究
方兆偉,劉金波*,王寶祥,遲 銘,宋兆強,李 健,楊 波,盧百關,劉 艷,周振玲,秦德榮,徐大勇*
(江蘇省連云港市農業科學院/江蘇省現代作物生產協同創新中心,江蘇 連云港 222006)
摘要:以兩系不育系9311S和恢復系WHR2雜交產生的P1、P2、F1、B1、B2、F2六個世代為研究材料,利用主基因+多基因混合遺傳模型分析了株高、穗長和每穗穎花數三個性狀的遺傳效應。結果表明:穗長和每穗穎花數的遺傳均符合1對加性主基因+加性-顯性多基因模型,株高表現為1對加性主基因+加性-顯性多基因模型或1對加性-顯性主基因+加性-顯性多基因模型,且均以加性效應為主。討論了9311S的應用前景。
關鍵詞:水稻;株高;穗長;每穗穎花數;遺傳
每穗穎花數是構成水稻產量的重要因子。適當提高植株高度,能夠增加群體生物產量,從而提高產量[1-3]。水稻穗長是與水稻產量密切聯系的重要農藝性狀,對水稻產量影響較大[4]。Rao等對20個高產品種的產量及其各構成性狀進行相關分析,發現穗長是對每個品種的產量具有最明顯且直接影響的4個產量相關性狀之一[5]。因此,研究每穗穎花數、穗長和株高等性狀的遺傳,對于提高作物產量具有非常重要的理論指導意義。應用現代遺傳學的理論與方法,從不同角度對水稻產量相關性狀進行深入探討,是現階段水稻超高產育種研究的重要內容[6]。前人大多采用NCII遺傳試驗設計來對性狀進行配合力分析,探討親本在雜種優勢利用或雜交育種中的利用價值[7-12]。雖然雙列雜交試驗實施簡便,但只能利用親代和F1代有限的遺傳信息,無法區分或估計加性和顯性以外的上位性遺傳效應[13]。以前作者曾利用植物數量性狀主基因+多基因混合遺傳模型軟件對P1、P2、F1、F2世代群體的株高、穗長、總粒數、粒長等性狀進行了遺傳分析[14-18]。這4個群體有限,所表達的信息有所限制。擴大遺傳群體世代,能在較大程度上闡明信息量。兩系光溫敏核不育系9311S是由連云港市農業科學院自主選育的,該不育系植株較矮,穗大粒多,與眾多恢復系測配,雜種F1代的株高介于雙親之間,抗倒伏,穗大。為了研究9311S株高的遺傳及其高產理論,本研究選用兩系不育系9311S和高稈大穗恢復系WHR2的雜交后代P1、P2、F1、B1、B2、F2六個世代為試驗材料,利用主基因+多基因混合遺傳模型,研究了水稻的株高、穗長和每穗穎花數的遺傳及其基因效應,探討了雜種優勢產生的遺傳機制,評價了各性狀的遺傳潛力和育種價值,以期為遺傳改良、雜種優勢的利用提供理論依據。
1材料與方法
1.1供試材料
2013年春天,在海南三亞,以兩系光溫敏核不育系9311S(P1)為母本,以恢復系WHR2(P2)為父本,配制雜交組合F1(9311S/WHR2)。2013年正季,在連云港市農業科學院玉帶河試驗基地,以9311S為母本,以F1為父本,配制B1(9311S//9311S/WHR2);同時,以F1去雄做母本,以P2作父本,配制B2(9311S/WHR2//WHR2)。成熟時,收獲F1代植株上的種子,形成F2群體。
1.2試驗方法
1.2.1田間種植與性狀測定方法 2014年正季,將試驗材料P1、P2、F1、F2、B1、B2六個世代種植于連云港市農業科學院玉帶河試驗基地。5月7日播種,6月13日移栽。親本和F1代每材料種植5行,每行10株。F2、B1、B2代分別種植600、450和221株。株行距16.7 cm23.0 cm。單本種植,常規栽培管理。成熟時,測量株高。同時每株取主莖穗考察穗長和每穗穎花數。對于親本和F1,均調查30株;對于其余世代,調查173~499株。
1.2.2數據分析方法 首先對試驗數據進行次數分布分析,利用南京農業大學章元明教授提供的植物數量性狀主基因+多基因混合遺傳模型軟件,分析性狀的遺傳模型,并根據模型估計主基因和多基因效應值及其方差等遺傳參數[19]。試驗數據處理在計算機軟件Excel中進行。
2結果與分析
2.19311S×WHR2組合的P1、P2、F1、B1、B2、F2世代的株高表現
從表1可以看出:9311S的平均株高為(80.92±4.09)cm, WHR2的平均株高為(146.18±4.65)cm,雙親差異顯著(P0.01); F1的平均株高為(128.48±5.41)cm,介于雙親之間,偏向于高親; B1的平均株高為(107.64±11.76)cm,偏向于低親; B2的平均株高為(130.23±13.42)cm,偏向于高親,而且出現超高親個體; F2的平均株高為(121.38±13.38)cm,偏向于高親,而且出現超高親個體; B1、B2和 F2群體均呈連續分布,只有1個峰,沒有顯示出有效應較大的主基因存在。
2.29311S×WHR2組合的P1、P2、F1、F2、B1、B2世代的穗長表現
從表2可以看出:9311S的平均穗長為(19.62±1.71)cm, WHR2的平均穗長為(27.03±1.79)cm,雙親差異顯著(P0.01); F1的平均穗長為(25.42±1.41)cm,介于雙親之間,偏向于高親; B1的平均穗長為(23.38±2.11)cm,接近于中親值; B2的平均穗長為(25.14±2.42)cm,偏向于高親; F2的平均穗長為(24.19±2.48)cm,偏向于高親,而且出現超高親個體。B1、B2和F2群體均呈連續分布,只有1個峰,沒有顯示出有效應較大的主基因存在。
2.39311S×WHR2組合的P1、P2、F1、F2、B1、B2世代的每穗穎花數表現
從表3可以看出:9311S的平均每穗穎花數為(233.03±44.30)粒, WHR2的平均每穗穎花數為(282.97±36.44)粒,雙親差異顯著(P0.01); F1的平均每穗穎花數為(355.30±57.09)粒,超過高親; B1的平均每穗穎花數為(301.54±71.88)粒,超過高親; B2的平均每穗穎花數為(321.14±82.22)粒,超過高親; F2的平均每穗穎花數為(325.60±111.00)粒,超過高親。B1、B2群體均呈連續分布,只有1個峰,沒有顯示出有效應較大的主基因存在。F2群體呈連續分布,有3個峰,顯示出有效應較大的主基因存在。
2.4遺傳參數的估計
經遺傳模型極大似然值和AIC值(赤池信息準則, Akaike’s Information Criterion)比較,似然比檢驗(LRT)(如表4),9311S×WHR2組合穗長和每穗穎花數的最適遺傳模型均為D-2模型,即這2個性狀均表現為1對加性主基因+加性-顯性多基因混合遺傳。株高的最適遺傳模型為D-1和D-2模型,即該性狀表現為1對加性-顯性主基因+加性-顯性多基因混合遺傳和1對加性主基因+加性-顯性多基因混合遺傳。
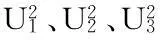
由遺傳模型(圖1)可以看出:在B1群體中株高存在2個成分分布,分布比例為1∶1,擬合后顯示為1個峰;在B2群體中株高也存在2個成分分布,分布比例為1∶1,擬合后顯示為1個峰;在F2群體中株高存在3個成分分布,分布比例為1∶2∶1,擬合后顯示為2個峰;穗長和每穗穎花數的圖形分布和擬合圖形分布與株高相似(D-1與D-2一樣結果,略)。
根據遺傳模型中成分分布及相應的均值,估計出模型中的一階參數和二階參數(表5~表6)。
由表5~表6可知,在9311S×WHR2組合中株高適合的遺傳模型為D-1,在該遺傳模型中,控制株高的1對主基因的基因顯性度為0.49,說明控制株高的1對主基因的加性效應和顯性效應都同等重要;植株高度高對矮呈部分顯性。在B1群體中,控制株高的主基因的遺傳率為12.4%,多基因的遺傳率為71.84%,說明株高以多基因遺傳為主。在B2群體中,控制株高的主基因的遺傳率為38.56%,多基因的遺傳率為49.33%,說明株高以多基因遺傳為主。在F2群體中,控制株高的主基因的遺傳率為44.89%,多基因的遺傳率為42.93%,說明株高主基因和多基因遺傳同等重要。
注: m為中親值; d為主基因加性效應; [d]為多基因加性效應; h為主基因顯性效應; [h]為多基因顯性效應。
在D-2模型中,控制株高的1對主基因以加性效應為主。在B1群體中,控制株高的主基因的遺傳率為12.4%,多基因的遺傳率為71.84%,說明株高以多基因遺傳為主。在B2群體中,控制株高的主基因的遺傳率為38.56%,多基因的遺傳率為49.33%,說明株高以多基因遺傳為主。在F2群體中,控制株高的主基因的遺傳率為44.89%,多基因的遺傳率為42.93%,說明株高主基因和多基因遺傳同等重要。
同樣,在9311S×WHR2組合中控制穗長的1對主基因以加性效應為主。在B1群體中,控制穗長的主基因的遺傳率為39.1%,多基因的遺傳率為14.61%,說明穗長以主基因遺傳為主。在B2群體中,控制穗長的主基因的遺傳率為64.85%,多基因的遺傳率為0.0%,說明穗長以主基因遺傳為主。在F2群體中,控制穗長的主基因的遺傳率為23.58%,多基因的遺傳率為42.93%,說明穗長以多基因遺傳為主。
在9311S×WHR2組合中控制每穗穎花數的1對主基因以加性效應為主。控制每穗穎花數的多基因的平均顯性度為-23.01,超親偏向于母本。在B1群體中,控制每穗穎花數的主基因的遺傳率為66.34%,多基因的遺傳率為5.69%,說明每穗穎花數以主基因遺傳為主。在B2群體中,控制每穗穎花數的主基因的遺傳率為66.39%,多基因的遺傳率為0.0%,說明每穗穎花數以主基因遺傳為主。在F2群體中,控制每穗穎花數的主基因的遺傳率為45.79%,多基因的遺傳率為38.95%,說明每穗穎花數以主基因遺傳為主。

3討論
水稻雜種優勢主要表現在每穗穎花數的增加上[20]。了解每穗穎花數的遺傳,可以有針對性地進行親本改良和水稻高產育種。筆者、陳獻功、江建華等研究認為每穗穎花數性狀均受2對主基因+多基因共同控制[17-18,21]。筆者等利用秈粳交材料研究發現粳稻每穗穎花數的遺傳表現為1對加性-顯性主基因+加性-顯性多基因遺傳[15]。而本研究發現每穗穎花數性狀的遺傳符合1對加性主基因+加性-顯性多基因模型,以加性效應為主,這可能與本研究的遺傳材料差異大有關。而且在B1、B2和F2三個群體中每穗穎花數均表現為主基因遺傳,因此在這3個群體中按照穗粒數性狀進行選擇是有效的,這為育種工作者選育大穗型品種提供了理論依據。
當稻穗較長時,籽粒排列疏松,籽粒灌漿充實,病害尤其是稻曲病發生較輕。朱春杰等利用秈粳群體分析了雜交后代穗部性狀的變異,發現穗粒重由3個主成分決定,粒數因子、穗型因子和粒重因子分別貢獻了45.49%、17.6%和12.22%,穗較長有使粒數增多、粒重增加的趨勢[22]。鄒文廣等研究發現,穗長受基因的加性效應和顯性效應的影響,以加性效應為主,而且遺傳率低,適于在晚代選擇[23]。本研究發現穗長性狀的遺傳符合1對加性主基因+加性-顯性多基因模型,以加性效應為主。在B1和B2群體中穗長均表現為主基因遺傳,在F2群體中表現為多基因遺傳,可見回交育種能加速穗長性狀的育種進程。
張忠旭等研究認為,在以生物產量的突破為主攻目標,選育超高產中高稈彎曲穗型雜交粳稻時,可以適當放寬對株高的限制,強化縮短第1、2節間長度,增加穗及穗莖節間長度在株高中的比例,以及加強基部莖稈各性狀的物理強度,即對抗倒性狀進行優化組合,提高水稻本身的耐肥抗倒性[24]。熊振民等認為,在構成株高的諸因素中,與每穗總粒數相關最密切的性狀是株高,其次是第II節間長。在矮化育種中并非是植株高度越矮越好,保證有一定的植株高度和適當的第I節間長度是很有必要的[25]。而且,熊振民等研究發現,2個秈稻矮稈品種其矮生性均受與sd1等位的一對隱性矮生基因控制,而l個粳稻矮稈品種受到多基因控制[25]。樊葉楊等研究發現在協青早/密陽46的F2群體中,控制株高性狀的基因存在部分顯性作用,以加性-顯性為主,兩個群體中均未檢測到顯性-顯性互作[26]。本研究發現株高性狀的遺傳符合1對加性主基因+加性-顯性多基因模型或1對加性-顯性主基因+加性-顯性多基因模型,以加性效應為主。而且在B1、B2和F2三個群體中株高均主要表現為多基因遺傳,說明控制9311S株高的遺傳受多基因控制,這可能與9311S的半矮稈性有關。
從3個性狀基因的遺傳來看,3個性狀均表現為加性效應遺傳,說明加性效應為該組合的優勢起主導作用,也反映出這3個性狀的一般配合力都好。為了充分利用雜種優勢,需要在一般配合力高的基礎上選擇特殊配合力好的組合,才能適應生產的需要。為了發揮9311S的矮稈部分顯性作用,可將其與遺傳距離較遠、豐產性好的恢復系雜交,選育高產、優質、抗倒雜交組合。在半矮稈基因的遺傳背景下,通過“矮中求高”,適當提高水稻植株的高度,選育抗倒、高產、優質雜交稻組合[27]。這點在實際育種操作中得到了體現。
參考文獻:
[1] 許乃霞,楊益花,蘇祖芳.抽穗后水稻株型與高光效群體形成關系的研究[J].耕作與栽培,2009(5):17-18.
[2] 蘇祖芳,許乃霞,孫成明,等.水稻抽穗后株型指標與產量形成關系的研究[J].中國農業科學,2003,36(1):115-120.
[3] 袁隆平.選育水稻亞種間雜交組合的策略[J].雜交水稻,1996,16(2):1-3.
[4] 郭小蛟,張濤,蔣開鋒,等.水稻秈粳交F1、F2群體穗長QTL比較分析[J].中國農業科學,2013,46(23):4849-4857.
[5] Rao S, Khan M, McNeilly T, et al. Cause and effect relations of yield and yield component in rice (OryzasativaL.)[J]. Journal of Genetics and Breeding, 1997, 51: 1-5.
[6] 林志強,鄭燕,蔡英杰,等.水稻長穗大粒RIL群體產量、穗部和谷粒性狀的遺傳分析[J].福建農林大學學報,2011,40(5):449-454.
[7] 廖伏明,周坤爐,盛孝邦,等.秈型三系雜交水稻主要農藝性狀配合力研究[J].作物學報,1999,25(5):622-631.
[8] 朱鎮,趙凌,宗壽余,等.秈型兩系雜交水稻主要農藝性狀的配合力分析[J].江蘇農業學報,2004,20(4):207-212.
[9] 周軍,鄒小云,周瑤敏.秈型雜交水稻產量及其相關性狀的配合力及遺傳分析[J].江西農業學報,2008,20(12):20-23.
[10] 譚美林,馮明友,張家洪,等.雜交水稻親本主要農藝性狀配合力及遺傳力分析[J].貴州農業科學,2009,37(3):6-8.
[11] 劉金波,宋兆強,王寶祥,等.幾個新選雜交水稻不育系和恢復系的主要農藝性狀的配合力分析[J].西南農業學報,2015.
[12] 洪德林,楊開晴,潘恩飛.粳稻不同生態類型間F1的雜種優勢及其親本的配合力分析[J].中國水稻科學,2002,16(3):216-220.
[13] 顧興友,鄭少玲,嚴小龍,等.水稻苗期耐鹽性遺傳的世代平均數分析[J].作物學報,1999,25(6):686-690.
[14] 劉金波,胡文德,王寶祥,等.水稻谷粒外觀性狀和粒重的遺傳研究[J].浙江農業學報,2014,26(3):543-548.
[15] 劉金波,王寶祥,劉曉麗,等.粳稻穗角、著粒密度及每穗穎花數的遺傳[J].江蘇農業學報,2014,30(5):936-942.
[16] 劉金波,劉曉麗,王寶祥,等.水稻一次枝梗數和二次枝梗數的遺傳[J].江蘇農業學報,2013,29(6):1187-1192.
[17] 江建華,張啟武,洪德林.粳稻穗部性狀遺傳分析[J].植物學報,2010,45(2):182-188.
[18] 劉金波,洪德林.粳稻穗角和每穗穎花數的遺傳分析[J].中國水稻科學,2005,19(3):223-230.
[19] 蓋鈞鎰,章元明,王建康.植物數量性狀遺傳體系[M].北京:科學出版社,2003:224-260.
[20] Virmani S S, Aquino B C, Khush G S. Heterosis breeding in rice (OryzasativaL.)[J]. Theor Appl Genet, 1982, 63(4): 373-380.
[21] 陳獻功,劉金波,洪德林.粳稻直立穗與彎曲穗3個雜交組合6個世代穗角和每穗穎花數的遺傳分析[J].作物學報,2006,32(8):1143-1150.
[22] 朱春杰,徐海,郭艷華,等.秈粳稻雜交后代穗部性狀變異及其相互關系研究[J].沈陽農業大學學報,2007,38(4):457-461.
[23] 鄒文廣,許旭明,張受剛,等.水稻長穗大粒種質資源產量性狀配合力及遺傳力分析[J].福建農業學報,2014,29(4):324-328.
[24] 張忠旭,馬興全,李全英,等.北方超級雜交粳稻育種研究[J].沈陽農業大學學報,2002,43(6):667-672.
[25] 熊振民,閔紹楷,俞桂林,等.云南稻種矮源的進一步篩選及其矮生性的遺傳研究[J].江蘇農學院學報,1989,10(3):1-5.
[26] 樊葉楊,莊杰云,李強,等.水稻株高QTL分析及其與產量QTL的關系[J].作物學報,2001,27(6):915-922.
[27] 陳俊宇,張振華,莊杰云.水稻株高QTL及其與產量性狀和抽穗期關系的研究進展[J].福建稻麥科技,2012,30(1):67-71.
(責任編輯:黃榮華)
Research on Inheritance of Plant Height, Panicle Length and Number of Spikelets per Panicle in Indica Rice (Oryzasativa)
FANG Zhao-wei, LIU Jin-bo*, WANG Bao-xiang, CHI Ming, SONG Zhao-qiang, LI Jian,YANG Bo, LU Bai-guan, LIU Yan, ZHOU Zhen-ling, QIN De-rong, XU Da-yong*
(Lianyungang Academy of Agricultural Science/Collaborative Innovation Center for Modern Crop Production of Jiangsu Province, Lianyungang 222006, China)
Abstract:The genetic effects of plant height, panicle length and number of spikelets per panicle in P1, P2, F1, B1, B2, F2six generations derived from the crossing of the male sterile line 9311S and restorer line WHR2 were studied based on the major gene-polygene mixed inheritance model. The results showed that the inheritances of the panicle length and spikelets per panicle were controlled by one major gene with additive effect plus polygene with additive-dominant effects, and plant height was mainly controlled by one major gene with additive effect plus polygene with additive-dominant effects, or one major gene with additive-dominant effect plus polygene with additive-dominant effects. For three traits, additive effects were principal. The application prospects of 9311S were discussed.
Key words:OryzasativaL.; Plant height; Panicle length; Spikelets per panicle; Inheritance
收稿日期:2015-10-07
基金項目:國家水稻產業體系(CARS-01-01A);江蘇省科技支撐項目(BE2014314);江蘇省農業三新工程項目[SXGC(2015)138];
作者簡介:方兆偉(1963─),男,江蘇連云港人,副研究員,從事水稻遺傳育種研究工作。*通訊作者:劉金波、徐大勇。
中圖分類號:S511.032
文獻標志碼:A
文章編號:1001-8581(2016)05-0014-06
中央財政農業技術推廣資金項目[TG(14)062];連云港科技攻關項目(CN1415)。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
