云南漢族人群CD6基因多態性與HCV慢性感染的關系*
2016-06-17 03:19:57劉城秀沈云松李桃意檀林萍姚月婷俞建昆姚宇峰
貴州醫科大學學報 2016年5期
劉城秀, 沈云松, 李桃意, 張 禹, 檀林萍, 姚月婷, 俞建昆, 姚宇峰, 史 荔**
(1.中國醫學科學院&北京協和醫學院 醫學生物學研究所, 云南 昆明 650118; 2.云南省重大傳染病疫苗研發重點實驗室, 云南 昆明 650118; 3.云南省第一人民醫院&昆明理工大學附屬醫院, 云南 昆明 650032)
?
云南漢族人群CD6基因多態性與HCV慢性感染的關系*
劉城秀1,2, 沈云松3, 李桃意1,2, 張禹1,2, 檀林萍1,2, 姚月婷1,2, 俞建昆1,2, 姚宇峰1,2, 史荔1,2**
(1.中國醫學科學院&北京協和醫學院 醫學生物學研究所, 云南 昆明650118; 2.云南省重大傳染病疫苗研發重點實驗室, 云南 昆明650118; 3.云南省第一人民醫院&昆明理工大學附屬醫院, 云南 昆明650032)
[摘要]目的: 探討云南漢族人群中CD6基因多態性與慢性丙型肝炎病毒(HCV)感染的關系。方法: 隨機抽取云南地區漢族人群HCV慢性感染患者434例,健康對照人群444例;采用TaqMan探針基因分型方法對2組人群的CD6基因3個SNPs(rs17824933、rs12360861、rs11230563)進行基因分型,構建CD6基因單倍型,評估上述3個SNPs與HCV慢性感染的關系。結果: 兩組人群CD6基因SNP-rs17824933 (C>G)、 SNP-rs12360861 (G>A)和SNP-rs11230563 (C>T)基因型頻率和等位基因頻率比較,差異無統計學意義(P>0.05);2組人群的SNP-rs17824933 (C>G)、 SNP-rs12360861 (G>A)和SNP-rs11230563 (C>T)單倍型頻率比較,差異無統計學意義(P>0.05)。結論: 此次調查的云南漢族群體的CD6基因SNP-rs17824933 (C>G)、 SNP-rs12360861 (G>A)和SNP-rs11230563 (C>T)與HCV慢性感染無關。
[關鍵詞]肝炎,丙型,慢性; 多態性單核苷酸; 基因,CD6; 云南
丙型肝炎病毒(hepatitis C virus,HCV)屬黃病毒科,為單股正鏈RNA病毒,基因組全長約9.6 kb。我國HCV感染率約為3.2%,感染者已達3 800萬人[1]。感染HCV后,約20%急性感染者可自發清除HCV,其余發展為慢性持續感染,繼而發展成肝纖維化、肝硬化甚至肝細胞癌[2]。因此,宿主免疫系統遺傳因素在HCV感染后的清除或持續感染中可能起到的作用研究一直是該方向的熱點之一。有研究發現我國不同人群中的HLA-B與慢性HCV感染關系密切[3-6]。CD6在T細胞的活化和增殖過程中起著重要作用。CD6基因位于人類染色體11q13,編碼一個分子量為100~130 kDa跨膜糖蛋白,這個跨膜糖蛋白包含3個富含半胱氨酸結構域的清道夫受體(scavenger receptor cysteine-rich,SRCR)和1個活化白細胞黏附分子(activated leukocyte cell adhesion molecule,ALCAM)的結合位點[7]。CD6主要表達在CD4+和CD8+T細胞,胸腺細胞,某些B細胞和自然殺傷細胞亞群上以及腦的某些區域[8]。但是,目前國內外研究者對CD6基因多態性與HCV慢性感染的相關性研究并沒有得到明確結論。本研究對云南地區漢族人群CD6基因內含子區域SNP-rs17824933 (C>G)和編碼區域SNP-rs12360861 (G>A)、SNP-rs11230563 (C>T)位點進行分析,探討這3個位點多態性與HCV慢性感染的關系。
1對象與方法
1.1對象
隨機抽取云南省434名HCV慢性感染者為病例組,診斷依據中華醫學會肝病學分會和中華醫學會感染病學分會2015年制定的《丙型肝炎防治指南》[9],血清ALT、AST、抗HCV、HCV-RNA陽性,排除其他肝炎的患者。隨機抽取同時期云南省正常健康個體444名為對照組,納入標準為受試者血清ALT、AST、抗HCV、HCV-RNA正常。所有受試者均為居住于云南地區的彼此無血緣關系漢族個體并簽署知情同意書,允許其樣本與臨床資料用于本次研究。病例組,年齡(43.27±10.23)歲,男性250(57.60%)例,女性184例(42.40%);對照組,年齡(42.74±9.07)歲,男性227例(51.13%),女性217例(48.87%)。兩組受試者基礎資料比較,差異無統計學意義(P>0.05),具有可比性。
1.2方法
1.2.1樣本采集采集受試者空腹靜脈血5 mL,用EDTA抗凝,置-80 ℃保存,按照Qiagen血基因組DNA提取試劑盒說明書提取外周血基因組DNA,置-20 ℃保存。1.2.2CD6基因SNP位點基因分型采用實時熒光定量PCR法進行SNP分型,由Applied Biosystems公司合成檢測rs17824933 (C>G),rs12360861 (G>A)及rs11230563 (C>T)位點的引物及TaqMan探針。針對每個SNP位點所設計的兩條探針分別用VIC和FAM進行熒光標記。羅氏LightCycler 480實時熒光定量PCR儀檢測SNP位點,并用LCS480 1.5.1.62軟件進行基因分型。PCR反應體積為20 μL,反應條件為95 ℃ 10 min預變性,92 ℃變性15 s、60 ℃退火1 min(共40個循環),40 ℃延伸 5 min。以3個已知基因型(野生純合子、突變純合子、雜合子)的樣品作為標準對照(均為本課題組經基因測序樣本驗證,SNP-rs17824933(C>G)樣本其野生純合子、突變純合子、雜合子的基因型分別是CC、GG及CG,SNP-rs12360861(G>A)為GG、AA及AG;SNP-rs11230563 (C>T)為CC、TT及CT)。
1.3統計學方法
采用Hardy-Weinberg平衡檢驗基因型頻率的代表性。χ2檢驗檢測對照組與病例組3個SNP基因型、等位基因頻率差異。SHEsis軟件程序計算連鎖不平衡,根據連鎖不平衡(Linkage Disequilibrium , LD)結果構建單倍型;兩位點間的LD關系用 D′表示,D′>0.8認為位點間強連鎖[10-11]。P<0.05為差異有統計學意義。
2結果2.1CD6多態性位點的等位基因頻率與基因型頻率
CD6基因3個SNP位點rs17824933 (C>G)、rs12360861 (G>A)及rs11230563 (C>T)的基因型分布在病例組和對照組中均符合Hardy-Weinberg平衡(P>0.05)。CD6基因3個SNP位點rs17824933 (C>G)、rs12360861 (G>A)及rs11230563 (C>T)的等位基因頻率與基因型頻率在病例組和對照組中的分布比較,差異無統計學意義(P>0.05),見表1~3。
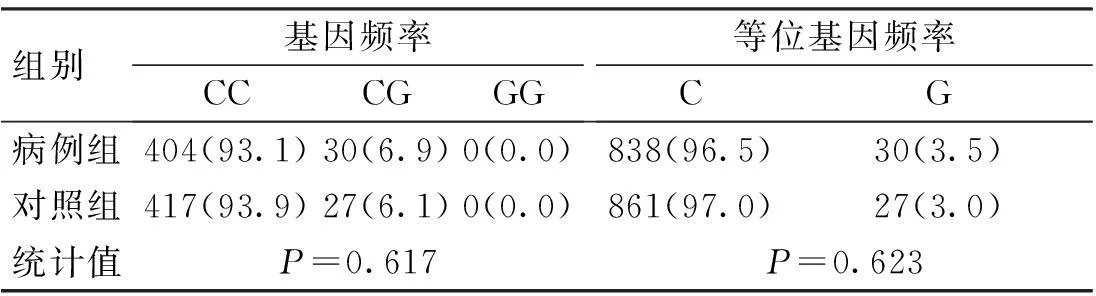
表1 CD6基因rs17824933位點的基因
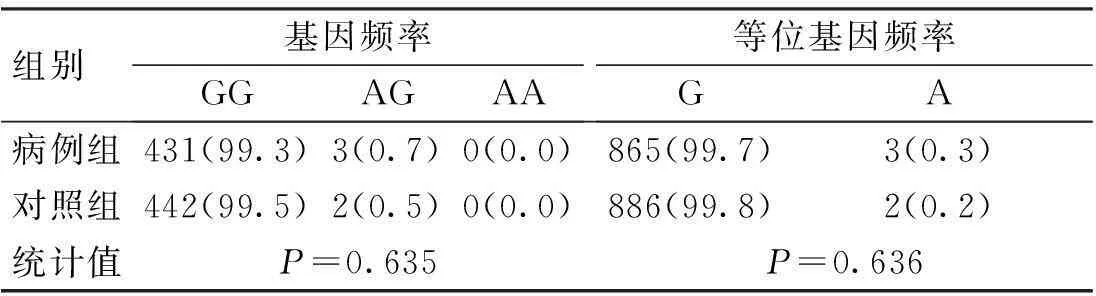
表2 CD6基因rs12360861位點的基因
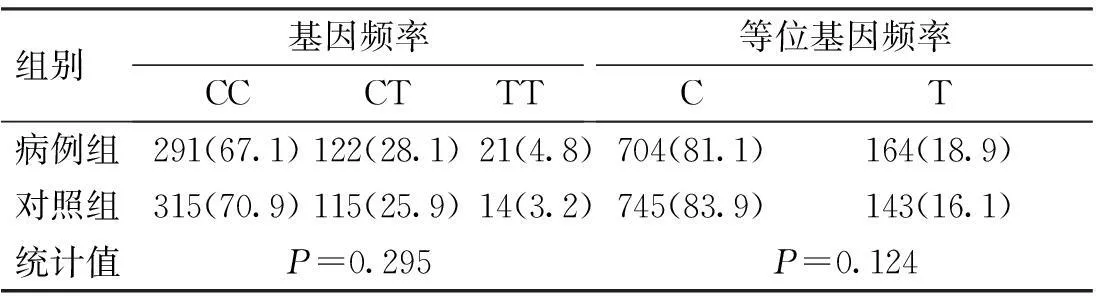
表3 CD6基因rs11230563位點的基因
2.2連鎖不平衡
CD6基因3個SNP位點rs17824933 (C>G)、rs12360861 (G>A)及rs11230563 (C>T)的連鎖不平衡分析結果顯示rs17824933 (C>G)、rs12360861 (G>A)及rs11230563 (C>T)位點之間存在強連鎖不平衡,見表4。
2.3CD6基因多態性位點的單倍型構建及頻率
構建CD6基因3個SNP位點的單倍型結果顯示,單倍型的頻率在病例組和對照組中的分布比較,差異無統計學意義(P>0.05),見表5。
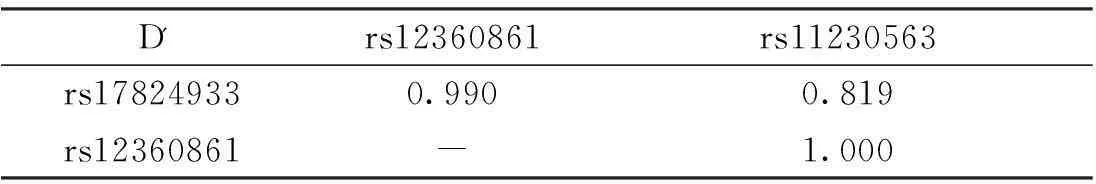
表4 CD6基因3個SNP位點連鎖不平衡分析
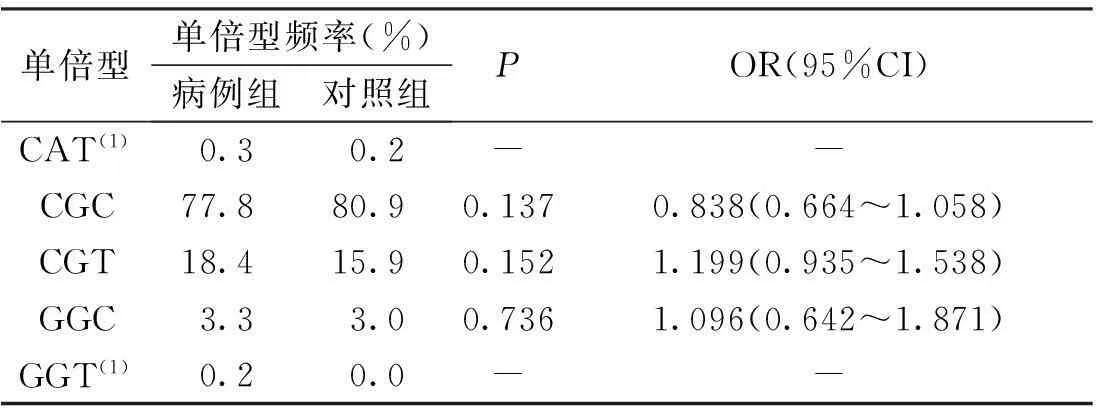
表5 病例組和對照組CD6基因3個
(1)SHEsis軟件程序忽略分析頻率<0.03的單倍型,不作統計分析
3討論
感染HCV是引起丙型肝炎發生的主要因素之一。HCV感染的主要方式有輸血傳播、靜脈吸毒傳播、性傳播和母嬰傳播。近年來,新發感染與靜脈吸毒和不潔注射有關。在云南,靜脈用藥人群中的77.7%可以檢測到抗-HCV抗體[12]。因此本研究選取云南漢族人群研究對象。
CD6是富含半胱氨酸結構域的SRCR超家族的成員,參與T細胞的活化和增殖、調節細胞間磷蛋白的表達和前炎性細胞活素的產生[13-14]。CD6刺激在維持T細胞活化過程中起重要作用,因阻斷與其配體ALCAM的相互作用而導致T細胞增殖的減少[15]。而且,由于CD6與中央超分子活化簇(central supramolecular activation cluster,cSMAC)區域的結合,會使CD6參與到免疫突觸的突變[16]。有研究表明,CD6基因多態性可能影響CD6的表達和功能,從而改變免疫應答水平而引起疾病的發生。De Jager等[17]發現,CD6基因多態性與多發性硬化癥(Multiple sclerosis,MS)有關。其中,CD6基因中的SNP-rs12360861 (G>A)位于第2富含半胱氨酸結構域的SRCR第5外顯子,編碼CD6與其配體ALCAM的結合位點;同時也處于叫做外顯子剪接增強子(exonic splicing enhancer,ESE)的順式作用元件中[18],通過無義突變、錯義突變或同義突變,ESE被破壞而影響mRNA的正確的持續合成能力[19]。因此,有學者提出假設認為G取代A會導致ESE被破壞,ALCAM與CD6配體受體結合減少,可能引起限制自體反應的T淋巴細胞通過血腦屏障的轉移,這樣對發生MS起保護作用[20]。Baratte等[21]的研究也表明第5 外顯子的選擇性剪接作用對調節T細胞的活化起到重要作用。Wagner等[20]對波蘭人群的研究發現,攜帶AA基因型的個體患MS的的風險比攜帶GG基因型的低3倍。CD6基因的另一個SNP-rs11230563 (C>T)位點位于第2富含半胱氨酸結構域的SRCR的第4 外顯子。Swaminathan等[8]對西班牙人群的研究發現,SRCR區域的非同義突變SNP-rs11230563可能引起CD6表達水平的改變以及與其配體結合能力或者信號傳導的改變;運用流式細胞熒光分析技術對MS患者的淋巴細胞進行實驗,所得結果發現初始的CD4+和CD8+T細胞比效應性和中心性記憶T細胞更容易受CD6單倍型的影響。CD6在胸腺細胞中的高表達并且認為有助于陽性選擇和抵抗細胞凋亡[22]。 SNP-rs17824933 (C>G)位于CD6基因第1內含子。Swaminathan等[23]研究表明SNP-rs17824933 (C>G)位點的G等位基因可能是罹患MS的危險因素。Kofler等[24]首次做了與疾病相關的CD6基因SNP 位點rs17824933與T細胞功能關系的研究,研究結果表明在CD6基因上的MS的風險等位基因與CD4+T細胞的異常增殖相關,并且證明了與疾病相關的等位基因在免疫學特性上的影響。rs12360861第1036位堿基G>A,引起第271個氨基酸由非極性脂肪族氨基酸突變為極性中性氨基酸,rs11230563第898位堿基C>T,引起第225個氨基酸由堿性氨基酸突變為芳香族氨基酸,均為錯義突變;rs17824933是在CD6基因非編碼區的第1內含子中發生了堿基的顛換。
本研究選取云南省漢族人群,研究CD6基因內含子區域SNP-rs17824933 (C>G)和編碼區域SNP-rs12360861 (G>A)、SNP-rs11230563 (C>T)3個位點與HCV慢性感染的關系,這些SNP與HCV慢性感染的無直接關聯。參照Ensembl網站SNP位點的基因頻率,在亞洲人群中rs17824933(C>G)等位基因G的基因頻率約為3%,而在歐洲人群中G的基因頻率高達23%;rs12360861(G>A)等位基因A在亞洲人群中頻率極低約為0.1%,本研究為0.3%,與此對比在歐洲人群中A的基因頻率為19%;rs11230563(C>T)的等位基因T在亞洲人群中的基因頻率約為17%,而在歐洲人群中該基因頻率為36%,但并未找到關于這3個SNP位點在亞洲、歐洲正常人群中和HCV感染者之間的關系研究。本研究未觀察到基于基因頻率的關聯性,需要在以后的研究中進一步開展病例分層分析,同時增加其他多態位點的檢測。
4參考文獻
[1] Zhang C, Wu N, Liu J. HCV subtype characterization among injection drug users: implication for a crucial role of Zhenjiang in HCV transmission in China [J]. PLoS One, 2011(6): e16817.
[2] Sebastiani G, Gkouvatsos K, Pantopoulos K. Chronic hepatitis C and liver fibrosis [J]. World J Gastroenterol, 2014(20): 11033-11053.
[3] Zuniga J, Romero V, Azocar J. Protective KIR-HLA interactions for HCV infection in intravenous drug users [J]. Mol Immunol, 2009(46): 2723-2727.
[4] Knapp S, Warshow U, Hegazy D. Consistent beneficial effects of killer cell immunoglobulin-like receptor 2DL3 and group 1 human leukocyte antigen-C following exposure to hepatitis C virus [J]. Hepatology, 2010(51): 1168-1175.
[5] 戎霞, 黃杰庭, 熊華平. 廣州獻血人群HLA多態性與慢性HCV感染的關聯研究 [J]. 中國輸血雜志, 2014(27):1116-1119.
[6] 劉錚, 康軼青, 張麗. 河南人群病毒性丙型肝炎與HLA多態性的關聯研究 [J]. 免疫學雜志, 2015(31):790-794.
[7] Bodian DL, Skonier JE, Bowen MA. Identification of residues in CD6 which are critical for ligand binding [J]. Biochemistry, 1997(36): 2637-2641.
[8] Swaminathan B, Cuapio A, Alloza I. Fine mapping and functional analysis of the multiple sclerosis risk gene CD6 [J]. PLoS One, 2013(8): e62376.
[9] 陳紅松, 竇曉光, 段鐘平. 丙型肝炎防治指南(2015年更新版) [J]. 臨床肝膽病雜志, 2015(31):1961-1979.
[10]Shi YY, He L, SHEsis.a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci [J]. Cell Res, 2005(15): 97-98.
[11]Li Z, Zhang Z, He Z. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: update of the SHEsis (http://analysis.bio-x.cn) [J]. Cell Res, 2009(19): 519-523.
[12]Zhou YH, Yao ZH, Liu FL. High prevalence of HIV, HCV, HBV and co-infection and associated risk factors among injecting drug users in Yunnan province, China [J]. PLoS One, 2012(7): e42937.
[13]Hassan NJ, Barclay AN, Brown MH. Frontline: Optimal T cell activation requires the engagement of CD6 and CD166 [J]. Eur J Immunol, 2004(34): 930-940.
[14]Oliveira MI, Goncalves CM, Pinto M. CD6 attenuates early and late signaling events, setting thresholds for T-cell activation [J]. Eur J Immunol, 2012(42): 195-205.
[15]Zimmerman AW, Joosten B, Torensma R. Long-term engagement of CD6 and ALCAM is essential for T-cell proliferation induced by dendritic cells [J]. Blood, 2006(107): 3212-3220.
[16]Castro MA, Oliveira MI, Nunes RJ. Extracellular isoforms of CD6 generated by alternative splicing regulate targeting of CD6 to the immunological synapse [J]. J Immunol, 2007(178): 4351-4361.
[17]De Jager PL, Jia X, Wang J. Meta-analysis of genome scans and replication identify CD6, IRF8 and TNFRSF1A as new multiple sclerosis susceptibility loci [J]. Nat Genet, 2009(41): 776-782.
[18]Desmet FO, Hamroun D, Lalande M. Human Splicing Finder: an online bioinformatics tool to predict splicing signals [J]. Nucleic Acids Res, 2009(37): e67.
[19]Baralle D, Baralle M. Splicing in action: assessing disease causing sequence changes [J]. J Med Genet, 2005(42): 737-748.
[20]Wagner M, Bilinska M, Pokryszko-Dragan A. ALCAM and CD6-multiple sclerosis risk factors [J]. J Neuroimmunol, 2014(276): 98-103.
[21]da Gloria VG, Martins de Araujo M, Mafalda Santos A. T cell activation regulates CD6 alternative splicing by transcription dynamics and SRSF1 [J]. J Immunol, 2014(193): 391-399.
[22]Singer NG, Fox DA, Haqqi TM. CD6: expression during development, apoptosis and selection of human and mouse thymocytes [J]. Int Immunol, 2002(14): 585-597.
[23]Swaminathan B, Matesanz F, Cavanillas ML. Validation of the CD6 and TNFRSF1A loci as risk factors for multiple sclerosis in Spain [J]. J Neuroimmunol, 2010(223): 100-103.
[24]Kofler DM, Severson CA, Mousissian N. The CD6 multiple sclerosis susceptibility allele is associated with alterations in CD4+ T cell proliferation [J]. J Immunol, 2011(187): 3286-3291.
(2016-01-06收稿,2016-03-25修回)
中文編輯: 吳昌學; 英文編輯: 趙毅
The Association between CD6 Polymorphisms and Chronic HCV Infection of Han Population in Yunnan Province
LIU Chengxiu1,2, SHEN Yunsong3, LI Taoyi1,2, ZHANG Yu1,2, TAN Linping1,2,YAO Yueting1,2, YU Jiankun1,2, YAO Yufeng1,2, SHI Li1,2
(1.InstituteofMedicalBiology,ChineseAcademyofMedicalSciences&PekingUnionMedicalCollege,Kunming650118,>Yunnan,China; 2.YunnanKeyLaboratoryofVaccineResearch&DevelopmentonSevereInfectiousDisease,Kunming650118,Yunnan,China; 3.TheFirstPeople'sHospitalinYunnanProvince&theAffiliatedHospitalofKunmingScienceandTechnologyUniversity,Kunming650032,Yunnan,China)
[Abstract]Objective: To investigate the association between CD6 gene polymorphism and chronic HCV chronic infection of Han population in Yunnan province. Methods: 434 HCV chronic infectious patients and 444 healthy individuals of Han Chinese population in Yunnan province were recruited for this study. Three single nucleotide polymorphisms (SNPs) of SNP-rs17824933, SNP-rs12360861 and SNP-rs11230563 of CD6 gene were determined by real-time TaqMan polymerase chain reaction and the haplotypes were constructed. Then evaluating the association correlation of these SNPs and haplotypes with HCV chronic infection. Results: The comparison of frequencies of allele and genotype of both groups of SNP-rs17824933 (C>G) , SNP-rs12360861 (G>A) and SNP-rs11230563 (C>T) showed that differences were not statistically significant (P>0.05). The frequency of haplotypes constructed by SNP-rs17824933 (C>G), SNP-rs12360861 (G>A) and SNP-rs11230563 (C>T) showed no significant difference (P>0.05). Conclusion: SNP-rs17824933 (C>G), SNP-rs12360861 (G>A) and SNP-rs11230563 (C>T) in the CD6 were not associated with HCV chronic infection in the Han population in Yunnan province.
[Key words]hepatitis C,chronic; polymorphism,single nucleotide; gene,CD6; Yunnan
*[基金項目]國家自然科學基金(81160197); 吳階平醫學基金(320.6750.13395)
[中圖分類號]R512.6
[文獻標識碼]A
[文章編號]1000-2707(2016)05-0527-05
DOI:10.19367/j.cnki.1000-2707.2016.05.008
**通信作者 E-mail:shili.imb@gmail.com
網絡出版時間:2016-05-13網絡出版地址:http://www.cnki.net/kcms/detail/52.5012.R.20160513.2015.006.html
