低氧環(huán)境對(duì)大鼠骨髓間充質(zhì)干細(xì)胞交感神經(jīng)重構(gòu)相關(guān)因子表達(dá)的影響
2016-06-02 09:07:04周智涓黃穗花
中西醫(yī)結(jié)合心腦血管病雜志 2016年6期
周智涓,伍 衛(wèi),黃穗花,劉 茂,陳 劍
中山大學(xué)附屬第五醫(yī)院(廣東珠海 519000)
?
低氧環(huán)境對(duì)大鼠骨髓間充質(zhì)干細(xì)胞交感神經(jīng)重構(gòu)相關(guān)因子表達(dá)的影響
周智涓,伍衛(wèi),黃穗花,劉茂,陳劍
中山大學(xué)附屬第五醫(yī)院(廣東珠海 519000)
摘要:目的觀察低氧環(huán)境對(duì)大鼠骨髓間充質(zhì)干細(xì)胞(BMSCs)三種交感神經(jīng)重構(gòu)相關(guān)因子內(nèi)皮素-1(ET-1)、促紅細(xì)胞生成素(EPO)和粒細(xì)胞集落刺激因子(G-CSF)表達(dá)的影響。方法體外分離培養(yǎng)大鼠BMSCs至第三代,應(yīng)用Western Blot技術(shù)檢測BMSCs各代細(xì)胞中ET-1、EPO和G-CSF蛋白表達(dá)水平。BMSCs培養(yǎng)至第三代后,更換無血清無糖培養(yǎng)基,將細(xì)胞培養(yǎng)皿置入缺氧培養(yǎng)箱分別培養(yǎng)2 h、6 h、12 h及24 h,分別檢測上述四組細(xì)胞ET-1、EPO、G-CSF蛋白表達(dá)水平。結(jié)果大鼠BMSCs在常氧狀態(tài)下具有分泌神經(jīng)重構(gòu)相關(guān)因子ET-1、EPO、G-CSF的功能。低氧環(huán)境可誘導(dǎo)大鼠BMSCs細(xì)胞ET-1、EPO、G-CSF蛋白表達(dá)水平上調(diào),ET-1、EPO表達(dá)水平隨缺氧時(shí)間延長而進(jìn)行性升高,差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。結(jié)論低氧環(huán)境能顯著上調(diào)大鼠BMSCs交感神經(jīng)重構(gòu)相關(guān)因子的表達(dá)水平,促進(jìn)心肌交感神經(jīng)重構(gòu),可能與BMSCs移植治療后心律失常副反應(yīng)的發(fā)生密切相關(guān)。
關(guān)鍵詞:骨髓間充質(zhì)干細(xì)胞;交感神經(jīng)重構(gòu);促紅細(xì)胞生成素;內(nèi)皮素-1;粒細(xì)胞集落刺激因子
骨髓間充質(zhì)干細(xì)胞(bone marrow-derived mesenchymal stem cells,BMSCs)是成體干細(xì)胞的一種,具有自我更新、自我維持以及多向分化的潛能,被認(rèn)為是當(dāng)前干細(xì)胞移植研究領(lǐng)域極具前景的種子細(xì)胞之一[1]。BMSCs可分化為血管內(nèi)皮細(xì)胞和血管平滑肌細(xì)胞,自分泌或促進(jìn)心肌細(xì)胞分泌促血管生長的細(xì)胞因子,從而直接和間接誘導(dǎo)新生血管形成,促進(jìn)側(cè)支循環(huán)建立,挽救缺血心肌[2-3]。
然而,BMSCs移植治療心肌梗死的安全性尚不確切。BMSCs移植治療具有致心律失常的副反應(yīng),目前觀點(diǎn)認(rèn)為其心律失常的發(fā)生可能與心肌交感神經(jīng)結(jié)構(gòu)和功能重構(gòu)密切相關(guān)[4-5]。近年體外研究還發(fā)現(xiàn),內(nèi)皮素-1(endothelin-1,ET-1)、促紅細(xì)胞生成素(erythropoietin,EPO)和粒細(xì)胞集落刺激因子(granulocyte colony-stimulated factor,G-CSF)具有促進(jìn)心肌梗死區(qū)神經(jīng)生長因子(nerve growth factor,NGF)的表達(dá)、提高交感神經(jīng)張力以及營養(yǎng)神經(jīng)的作用,從而促進(jìn)心肌交感神經(jīng)重構(gòu)的發(fā)生和發(fā)展[6-8]。那么,BMSCs移植治療的致心律失常作用是否與其分泌上述神經(jīng)重構(gòu)相關(guān)因子(ET-1、EPO、G-CSF)有關(guān),以及低氧誘導(dǎo)環(huán)境下能否上調(diào)BMSCs細(xì)胞上述因子的表達(dá)水平呢?目前尚無相關(guān)文獻(xiàn)報(bào)道。本實(shí)驗(yàn)通過體外分離培養(yǎng)大鼠BMSCs,觀察常氧條件下大鼠BMSCs能否分泌神經(jīng)重構(gòu)相關(guān)因子ET-1、EPO、G-CSF及其分泌特點(diǎn);利用低氧誘導(dǎo)模擬心肌梗死缺氧環(huán)境,進(jìn)一步觀察低氧環(huán)境對(duì)大鼠BMSCs交感神經(jīng)重構(gòu)相關(guān)因子表達(dá)的影響,從而為探索BMSCs移植治療致心律失常的原因提高實(shí)驗(yàn)依據(jù)。
1材料與方法
1.1實(shí)驗(yàn)動(dòng)物清潔級(jí)雄性Sprague-Dawley(SD)大鼠,體質(zhì)量80 g~120 g,3周~4周齡,共6只,由中山大學(xué)實(shí)驗(yàn)動(dòng)物中心提供(實(shí)驗(yàn)動(dòng)物質(zhì)量合格證號(hào):SCXK粵2012-0029)。實(shí)驗(yàn)過程中對(duì)動(dòng)物的處置符合動(dòng)物倫理學(xué)標(biāo)準(zhǔn)。
1.2主要試劑和儀器胎牛血清(美國Hyclone公司);DMEM-F12培養(yǎng)基(美國Hyclone公司);甘油醛-3-磷酸脫氫酶兔抗鼠多克隆抗體(Anti-GAPDH antibody,美國Abcam公司);兔抗鼠內(nèi)皮素-1單克隆抗體(Anti-ET-1 antibody,美國Abcam公司);重組促紅細(xì)胞生成素多克隆抗體(Anti-EPO antibody,美國CST公司);兔抗鼠粒細(xì)胞集落刺激因子單克隆抗體(Anti-G-CSF antibody,美國PeproTech 公司);超凈工作臺(tái)(美國Thermo Forma公司);缺氧細(xì)胞培養(yǎng)箱(美國Thermo Forma公司);熒光倒置顯微鏡(德國Leica公司);超低溫冰箱(美國Thermo公司)等。
1.3BMSCs分離、培養(yǎng)及鑒定取3周~4周齡SD大鼠,無菌條件下分離中雙側(cè)股骨及脛骨,采取全骨髓貼壁分離和消化控制相結(jié)合分離純化BMSCs。顯微鏡下觀察原代(P0)BMSCs生長情況及形態(tài)變化,細(xì)胞增殖至培養(yǎng)皿80%~90%時(shí),處于對(duì)數(shù)生長期,用0.25%胰蛋白酶-EDTA消化,離心機(jī)離心、重懸,傳代培養(yǎng)。首次傳代為P1,其后依次為P2、P3等,每天觀察細(xì)胞生長情況。采用流式細(xì)胞儀檢測P3期BMSCs細(xì)胞表面抗原CD44H、CD45、CD29、CD11b/c和CD90.1的表達(dá)情況。
1.4實(shí)驗(yàn)分組為分析常氧環(huán)境BMSCs細(xì)胞分泌神經(jīng)重構(gòu)相關(guān)因子ET-1、EPO、G-CSF特點(diǎn),按照細(xì)胞培養(yǎng)周期將細(xì)胞分為3組:P1期(第一代BMSCs細(xì)胞)、P2期(第二代BMSCs細(xì)胞)、P3期(第三代BMSCs細(xì)胞)。分別檢測三代細(xì)胞ET-1、EPO、G-CSF蛋白表達(dá)水平。為分析低氧環(huán)境對(duì)大鼠BMSCs細(xì)胞分泌神經(jīng)重構(gòu)相關(guān)因子的影響,取第三代BMSCs細(xì)胞進(jìn)行試驗(yàn)。BMSCs培養(yǎng)至第三代(P3期)后,更換無血清無糖培養(yǎng)基,將細(xì)胞培養(yǎng)皿置入94%N2、5%CO2、1%O2缺氧培養(yǎng)箱分別培養(yǎng)2 h、6 h、12 h及24 h,分別檢測上述4組細(xì)胞ET-1、EPO、G-CSF蛋白表達(dá)水平。
1.5蛋白印跡每瓶細(xì)胞加入裂解液后,用槍吹打數(shù)下,置于冰上并用水平搖床搖晃使裂解液和細(xì)胞充分接觸。充分裂解后,于4 ℃,12 000 r/min離心10 min,取上清。提取總蛋白后,立即測定蛋白濃度。采用BCA法進(jìn)行蛋白定量試劑盒進(jìn)行定量。按說明書稀釋標(biāo)準(zhǔn)品以及配制BCA工作液。將提取的總蛋白加入5×SDS蛋白緩沖液,100 ℃煮沸5 min,置于超低溫冰箱保存?zhèn)溆谩⒌鞍子肂radford法定量后,選擇不同濃度的聚丙烯酰胺凝膠(G-CSF15%,ET-1 12%,EPO12%),經(jīng)SDS-PAGE電泳后將蛋白轉(zhuǎn)至PVDF膜上。5%脫脂牛奶封閉后,加入相應(yīng)的一抗,室溫下孵育1 h~2 h或者4 ℃過夜。用TBST每次5 min洗3次后,用相應(yīng)的稀釋好的二抗,室溫下孵育1 h~2 h后,用TBST沖洗3次,每次10 min,然后進(jìn)行化學(xué)發(fā)光顯影。用Gel-Proanalyzer4.0分析目標(biāo)條帶的光密度值。本實(shí)驗(yàn)采用GAPDH作為內(nèi)參照,比較不同處理后上述蛋白表達(dá)的差異。
2結(jié)果
2.1BMSCs細(xì)胞鑒定顯微鏡下形態(tài)學(xué)觀察可見BMSCs為長梭形或纖維狀細(xì)胞,形態(tài)均一,排列密集,呈放射狀或漩渦狀生長(圖1A)。利用流式細(xì)胞儀檢測干細(xì)胞表面標(biāo)志物,結(jié)果提示細(xì)胞CD29、CD90.1表達(dá)呈陽性,CD34、CD45和CD11b/c呈陰性,符合BMSCs細(xì)胞表面標(biāo)志物特點(diǎn)(圖1B)。
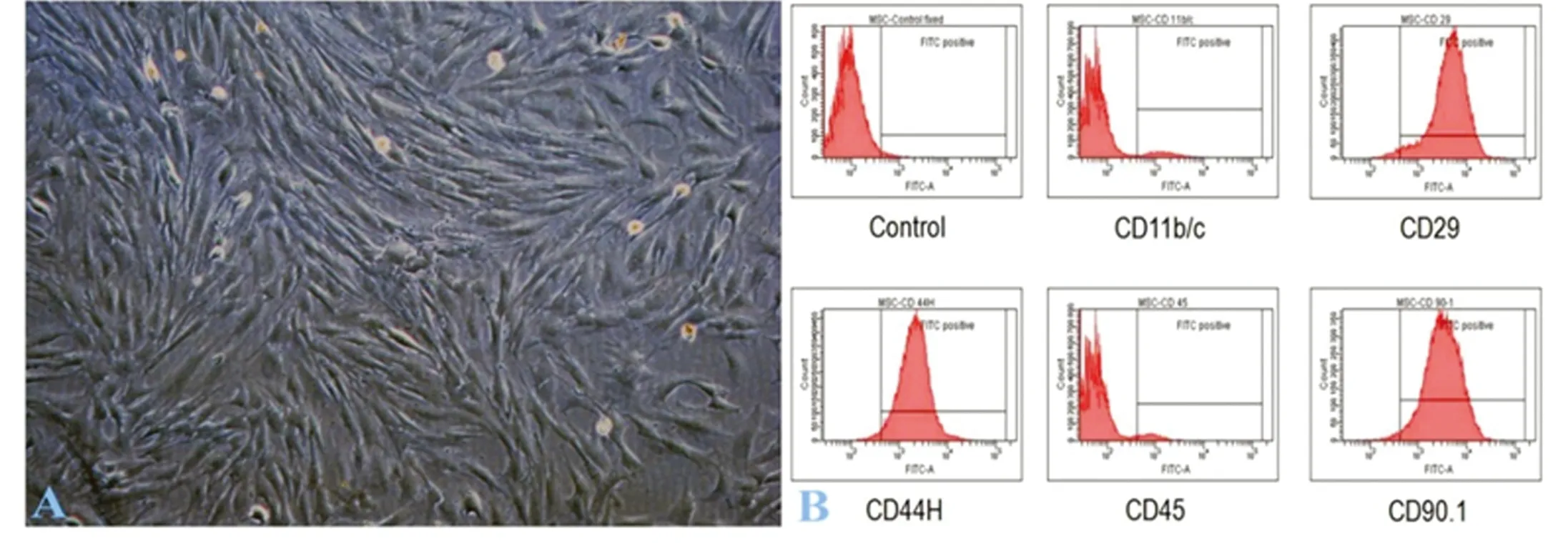
圖1 BMSCs細(xì)胞形態(tài)學(xué)及流式細(xì)胞儀鑒定
2.2常氧環(huán)境BMSCs細(xì)胞ET-1、EPO和G-CSF蛋白表達(dá)應(yīng)用Western Blot技術(shù)檢測BMSCs細(xì)胞 P1、P2及P3三個(gè)培養(yǎng)周期中ET-1、EPO和G-CSF蛋白表達(dá)水平。結(jié)果發(fā)現(xiàn),常氧環(huán)境中BMSCs細(xì)胞P1、P2、P3三個(gè)培養(yǎng)周期均可檢測到ET-1、EPO和G-CSF蛋白表達(dá),提示BMSCs細(xì)胞能夠自分泌上述三種蛋白。P3期BMSCs細(xì)胞ET-1蛋白表達(dá)較P1、P2期顯著增高,差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。P2期與P1期BMSCs細(xì)胞比較,ET-1蛋白表達(dá)差異無統(tǒng)計(jì)學(xué)意義(P>0.05)。詳見圖2。P2期BMSCs細(xì)胞EPO蛋白表達(dá)水平較P1、P3期增高(P<0.05),而P3期與P1期BMSCs細(xì)胞比較發(fā)現(xiàn)前者EPO蛋白表達(dá)水平明顯升高,差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。詳見圖3。P2和P3期BMSCs細(xì)胞G-CSF蛋白水平較P1期增高(P<0.05),而P2和P3 期之間G-CSF蛋白表達(dá)水平差異無統(tǒng)計(jì)學(xué)意義(P>0.05)。詳見圖4。
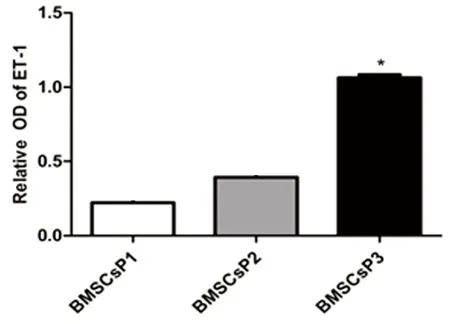
注:與BMSCs P1、P2期比較,* P<0.05。
圖2常氧環(huán)境BMSCs細(xì)胞ET-1蛋白表達(dá)
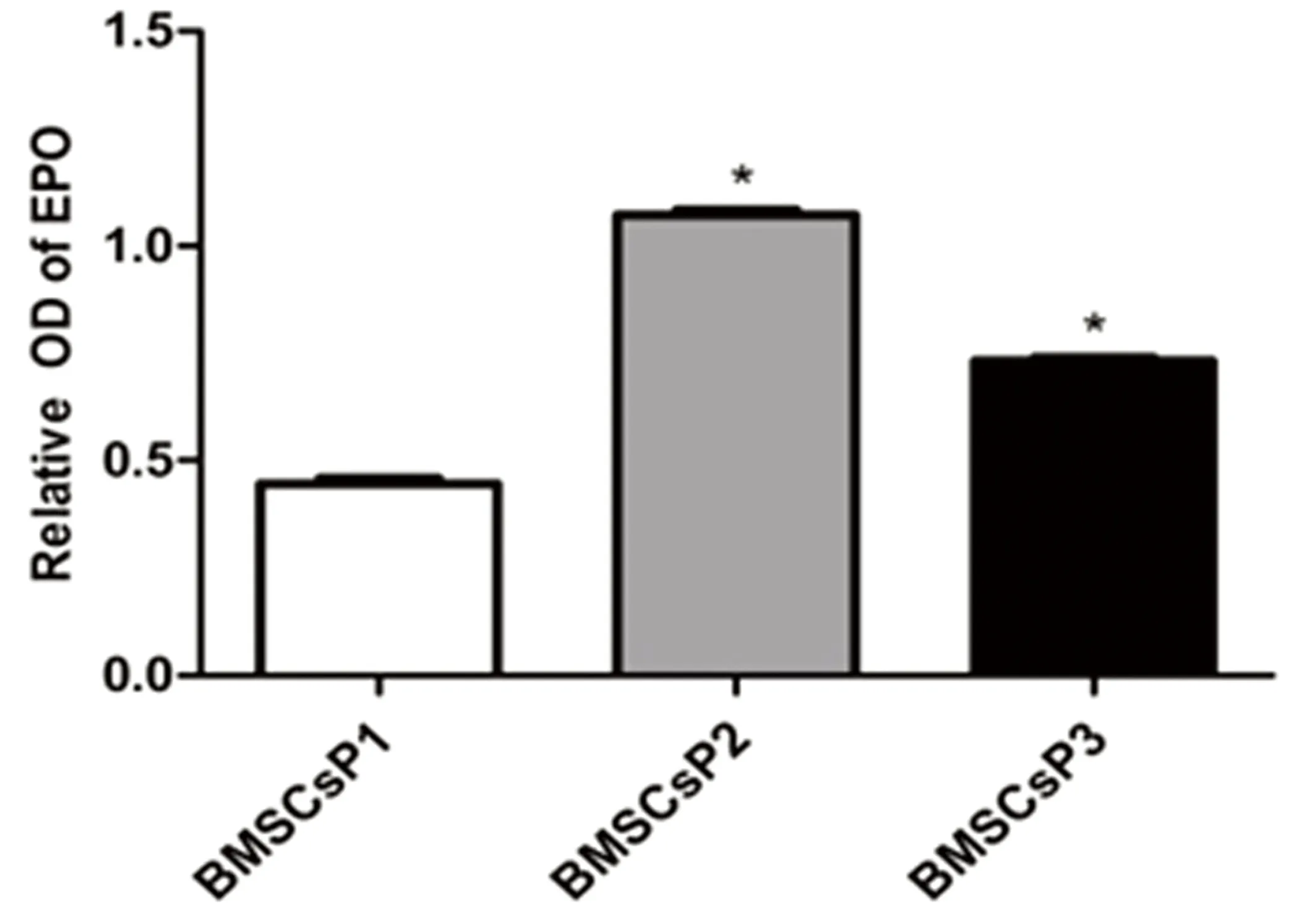
注:與BMSCs P1期比較,*P<0.05。
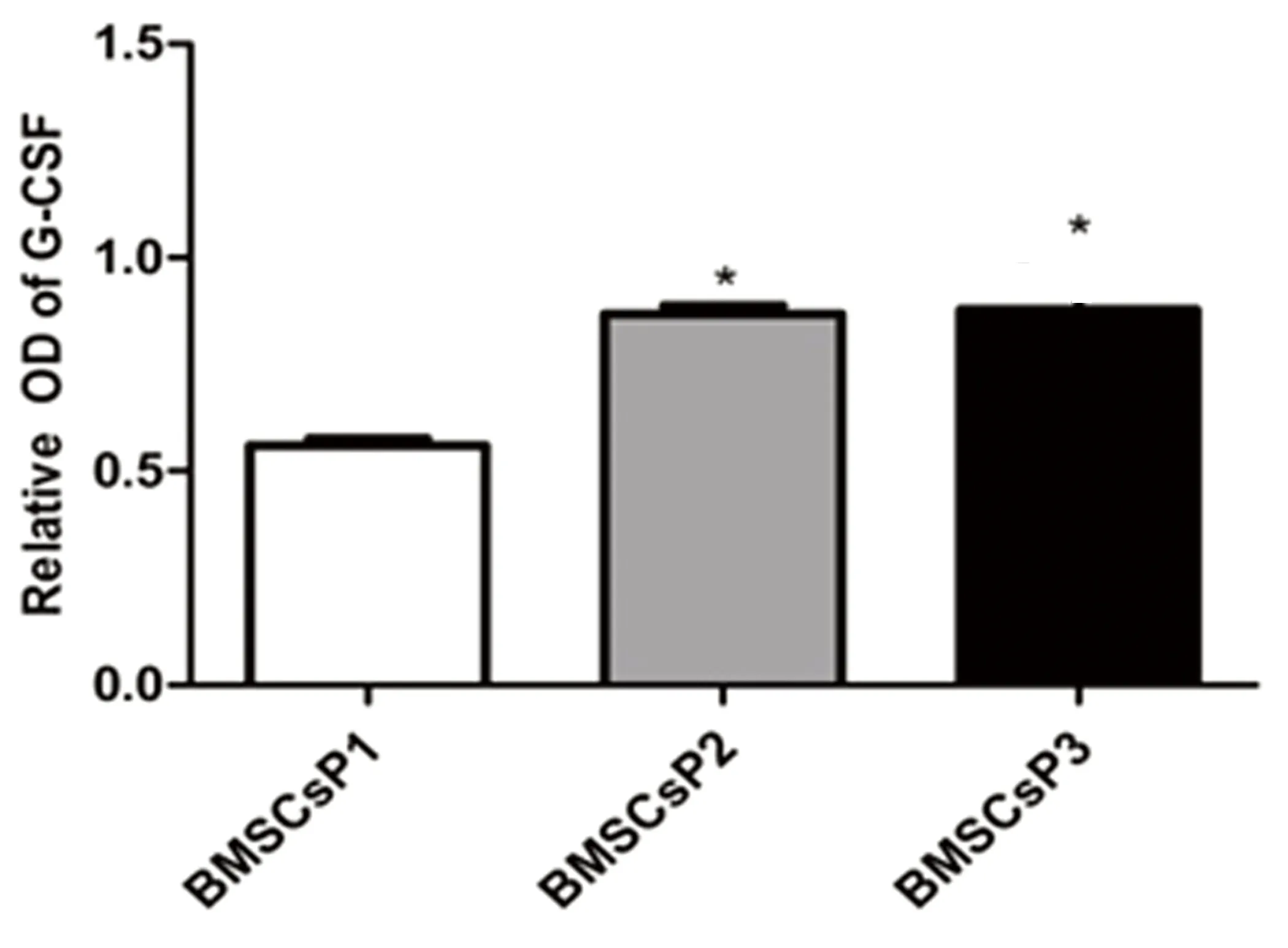
注:與BMSCs P1期比較,*P<0.05。
2.3低氧環(huán)境BMSCs細(xì)胞ET-1蛋白表達(dá)低氧誘導(dǎo)環(huán)境下BMSCs細(xì)胞ET-1蛋白表達(dá)量較常氧對(duì)照組顯著增加(P<0.05),且隨缺氧時(shí)間延長而逐漸增高,在缺氧24 h達(dá)到高峰(P<0.05),提示低氧環(huán)境可誘導(dǎo)BMSCs細(xì)胞ET-1蛋白表達(dá)水平上調(diào)。詳見圖5。
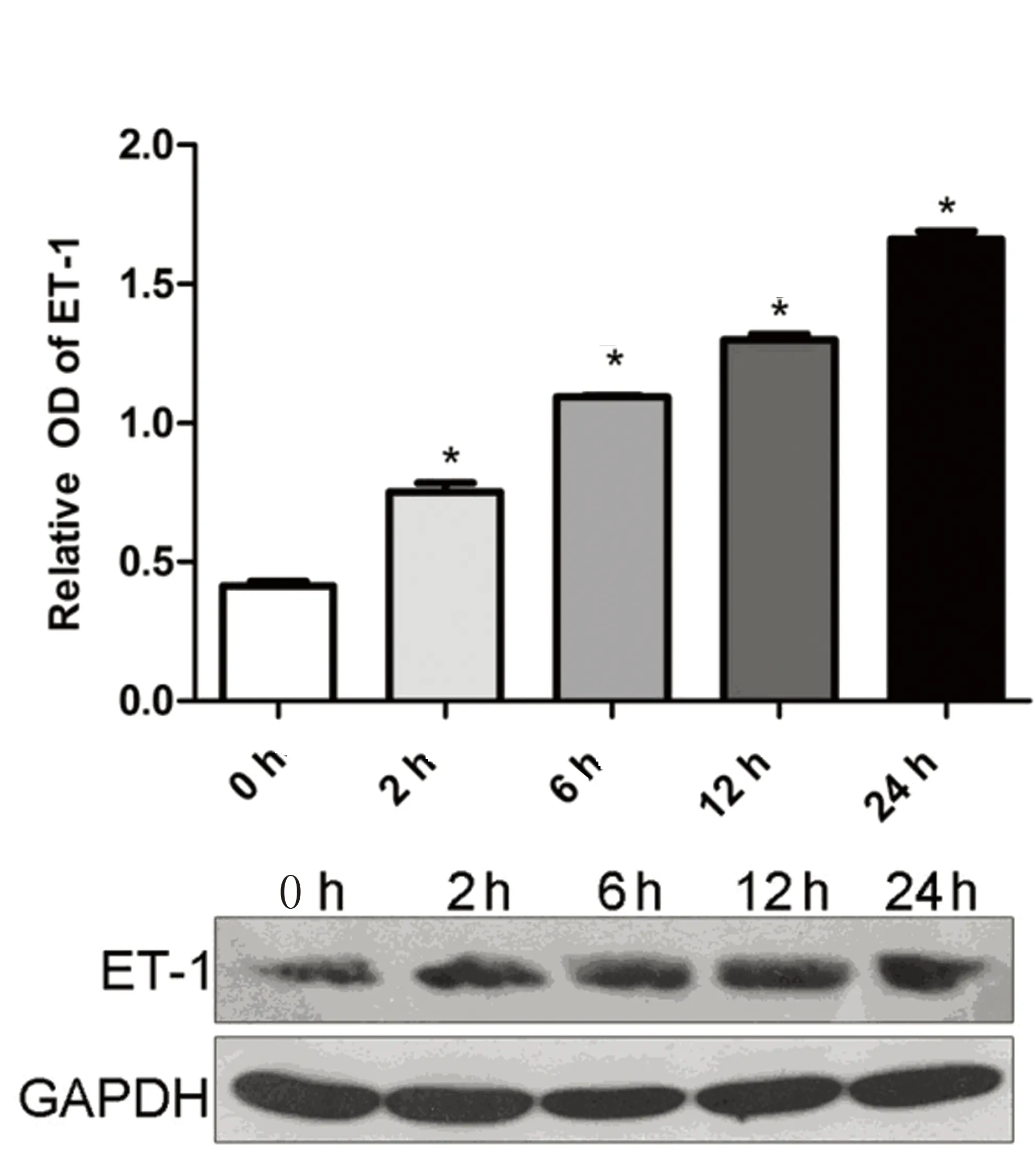
注:與對(duì)照組比較,*P<0.05。
2.4低氧環(huán)境BMSCs細(xì)胞EPO蛋白表達(dá)低氧誘導(dǎo)環(huán)境下BMSCs細(xì)胞EPO蛋白分泌量較常氧對(duì)照組明顯升高(P<0.05),并隨缺氧時(shí)間延長而進(jìn)行性升高,在缺氧24 h達(dá)到高峰(P<0.05),提示低氧環(huán)境可誘導(dǎo)BMSCs細(xì)胞EPO蛋白表達(dá)水平上調(diào)。詳見圖6。
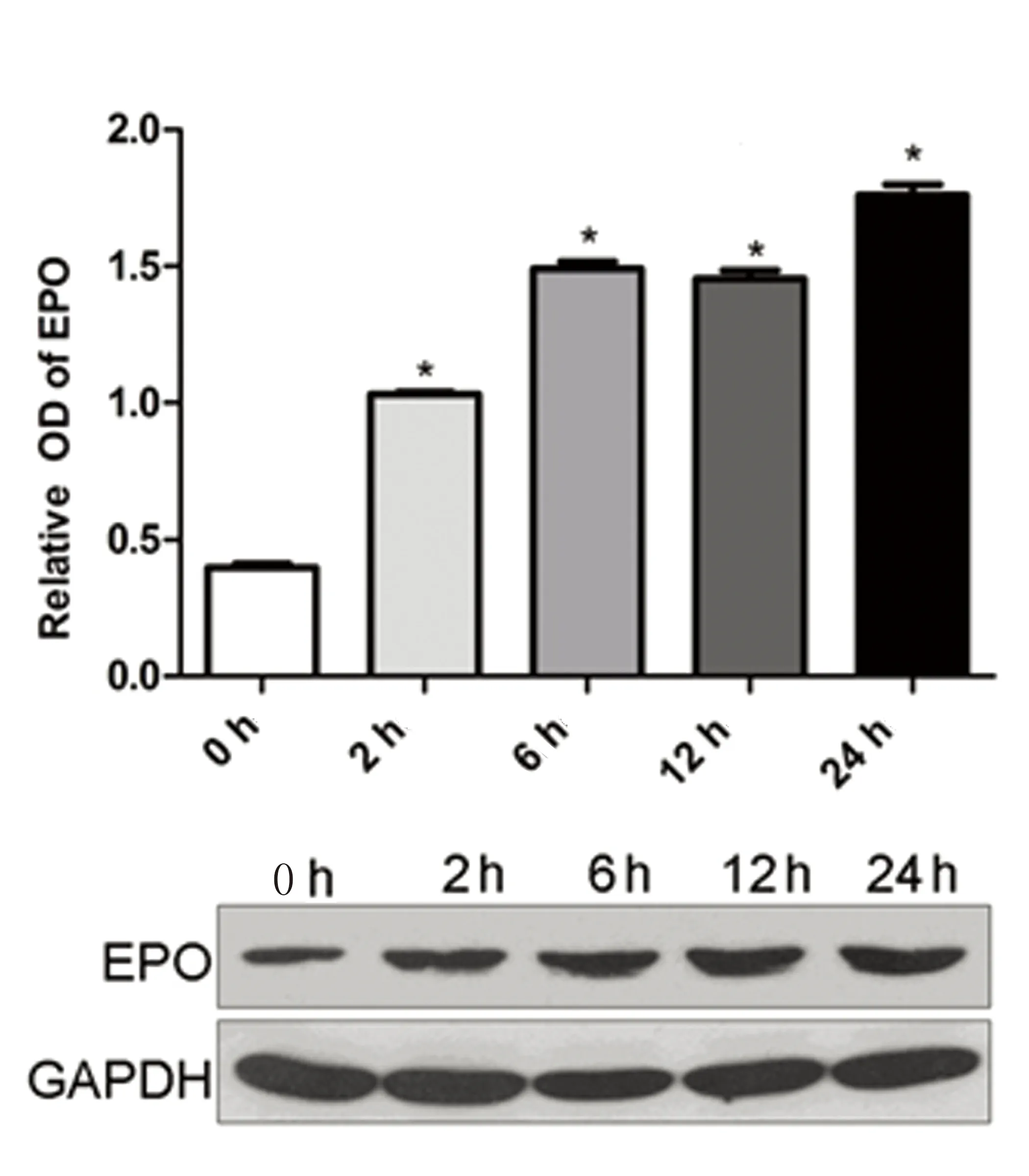
注:與對(duì)照組比較,* P<0.05。
圖6低氧環(huán)境BMSCs細(xì)胞EPO蛋白表達(dá)
2.5低氧環(huán)境BMSCs細(xì)胞G-CSF蛋白表達(dá)低氧誘導(dǎo)環(huán)境下BMSCs細(xì)胞G-CSF蛋白分泌量較常氧對(duì)照組顯著增加(P<0.05),但G-CSF蛋白表達(dá)量未隨缺氧時(shí)間延長而進(jìn)一步增加。詳見圖7。
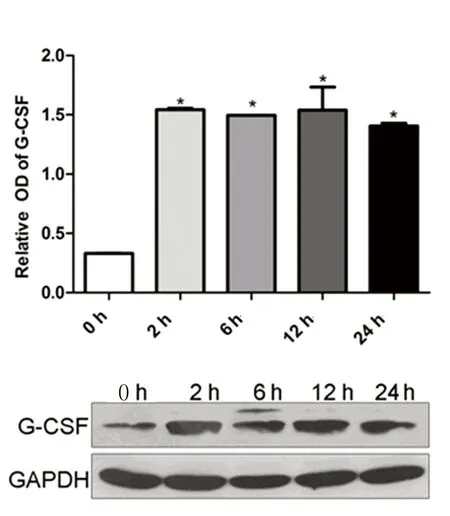
注:與對(duì)照組比較,* P<0.05。
3討論
干細(xì)胞是一類具有自我更新、自我維持和多向分化潛能的細(xì)胞,能表達(dá)、合成或分泌生長因子、細(xì)胞因子、調(diào)節(jié)肽及氣體信號(hào)分子等多種生物活性因子。而這些生物活性因子又具有調(diào)節(jié)組織細(xì)胞代謝、分化、增殖、遷移、凋亡等功能[9]。植入干細(xì)胞的分泌功能不僅直接影響植入干細(xì)胞自身的存活和分化,而且是干細(xì)胞移植發(fā)揮其他生物學(xué)效應(yīng)的重要機(jī)制[10]。本研究通過體外分離培養(yǎng)大鼠BMSCs發(fā)現(xiàn),常氧條件下大鼠BMSCs具有分泌神經(jīng)重構(gòu)相關(guān)因子ET-1、EPO、G-CSF的作用,低氧環(huán)境能夠顯著增加上述因子的表達(dá)水平,加劇心肌交感神經(jīng)重構(gòu),從而促使BMSCs移植治療后心律失常的發(fā)生。
ET-1是目前所知最強(qiáng)大的血管收縮因子,最早從豬主動(dòng)脈內(nèi)皮細(xì)胞上清液中分離發(fā)現(xiàn)。ET-1主要來源于內(nèi)皮細(xì)胞,可通過旁分泌的形式激活內(nèi)皮素受體并參與心血管系統(tǒng)多種生理病理過程[11]。研究發(fā)現(xiàn),ET-1與ET-1受體結(jié)合后,可激活P38MAPK、ERK等信號(hào)通路,誘導(dǎo)心肌細(xì)胞表達(dá)與分泌NGF,從而促進(jìn)交感神經(jīng)重構(gòu)[12]。在干細(xì)胞相關(guān)研究中發(fā)現(xiàn),脂肪源性MSCs可表達(dá)ET-1,但骨髓源性MSCs是否同樣表達(dá)ET-1,尚未見文獻(xiàn)報(bào)道。本研究觀察到BMSCs在P1、P2、P3三代細(xì)胞均可表達(dá)ET-1,且在純化的P3中,ET-1表達(dá)量最高。通過體外單一細(xì)胞培養(yǎng),排除了造血干細(xì)胞、造血細(xì)胞如單核細(xì)胞、淋巴細(xì)胞、成纖維細(xì)胞、巨噬細(xì)胞等干擾后,本研究首次發(fā)現(xiàn)骨髓來源的干細(xì)胞BMSCs同樣具有分泌ET-1的作用。本研究還觀察低氧處理對(duì)BMSCs細(xì)胞 24 h內(nèi)ET-1表達(dá)量的影響。首次發(fā)現(xiàn)低氧可增強(qiáng)BMSCsET-1的表達(dá),并發(fā)現(xiàn)在一定缺氧時(shí)程內(nèi),ET-1表達(dá)量隨缺氧時(shí)程延長而增高,繼續(xù)延長缺氧時(shí)間,ET-1表達(dá)量逐漸回降。這一結(jié)果提示低氧可調(diào)節(jié)BMSCs自分泌功能并可能在缺氧早期即發(fā)揮這一作用,提示這種機(jī)制可能與BMSCs移植治療的致心律失常副反應(yīng)相關(guān)。
另外,BMSCs由中胚層分化而成,可分泌多種支持造血因子,為BMSCs的存活提供營養(yǎng)支持及支架作用。既往研究發(fā)現(xiàn),BMSCs可分泌和表達(dá)EPO和G-CSF[13-15]。EPO作為強(qiáng)大的神經(jīng)營養(yǎng)因子,具有促進(jìn)軸突再生的作用,還可促進(jìn)腦和脊髓的星形膠質(zhì)細(xì)胞分泌NGF,從而發(fā)揮神經(jīng)保護(hù)作用[16]。G-CSF是一種促進(jìn)粒細(xì)胞生長的細(xì)胞因子,前期研究證實(shí)G-CSF可通過與粒細(xì)胞集落刺激因子受體結(jié)合,促進(jìn)干細(xì)胞遷移及定位[14,17]。Lee等[8]研究發(fā)現(xiàn)G-CSF可促進(jìn)心肌梗死后大鼠梗死區(qū)域NGF表達(dá)及交感神經(jīng)重分布。而Kuhlmann和Baldo等學(xué)者則持相反觀點(diǎn),認(rèn)為G-CSF或可通過增加Connexin43表達(dá)而減少心肌梗死室性心律失常的發(fā)生[1,14]。在本研究中,通過蛋白印跡法檢測未分化的BMSCs細(xì)胞EPO和G-CSF蛋白表達(dá)水平,也發(fā)現(xiàn)BMSCs可分泌上述細(xì)胞因子,且晚期成熟的BMSCs中更為明顯,與國內(nèi)外研究結(jié)論一致[15,18]。同時(shí),本研究還發(fā)現(xiàn),缺氧環(huán)境可誘導(dǎo)EPO和G-CSF蛋白表達(dá)增加,前者隨缺氧時(shí)間增加而增加,后者則不隨缺氧作用時(shí)間變化而進(jìn)一步變化。該現(xiàn)象說明缺氧可影響B(tài)MSCs細(xì)胞EPO表達(dá),或可通過持續(xù)分泌EPO而促進(jìn)NGF表達(dá),促進(jìn)交感神經(jīng)重構(gòu),從而誘發(fā)室性心律失常。另外,缺氧也可促進(jìn)BMSCs細(xì)胞G-CSF蛋白表達(dá),但是否能夠通過G-CSF表達(dá)水平變化這一途徑而影響心肌交感神經(jīng)重構(gòu)尚有待后續(xù)研究探索。
低氧環(huán)境可能通過以下途徑影響B(tài)MSCs細(xì)胞交感神經(jīng)重構(gòu)相關(guān)因子的表達(dá):低氧可增加抗凋亡蛋白Bcl-2的表達(dá),抑制P53信號(hào)表達(dá),抑制白介素1β轉(zhuǎn)化酶生成,減少氧化應(yīng)激損傷和氧自由基生成,穩(wěn)定線粒體電位,從而保持移植入宿主的干細(xì)胞存活率[19];低氧可增強(qiáng)了干細(xì)胞的分化能力[20];低氧還可調(diào)節(jié)干細(xì)胞某些基因和受體表達(dá)并影響多個(gè)細(xì)胞信號(hào)通路,如ERK1/2、P38 MAPK、JNK通路等,參與其下游多種功能的調(diào)節(jié),促進(jìn)多種生物活性因子的合成與分泌[19,21]。
本研究具有一些不足之處。首先,本研究僅為初期體外實(shí)驗(yàn),暫未進(jìn)行在體實(shí)驗(yàn)或人體研究。其次,既往研究提示外源性ET-1可誘導(dǎo)BMSCs分化成心肌樣細(xì)胞,本研究采用第1代~第3代BMSCs細(xì)胞進(jìn)行研究,結(jié)果發(fā)現(xiàn)早期BMSCs可表達(dá)ET-1,而BMSCs自分泌的ET-1又能否促進(jìn)其自身分化并影響其他細(xì)胞因子的分泌,本研究未進(jìn)一步探討。再次,本研究僅報(bào)告了上述三種細(xì)胞因子的蛋白表達(dá)水平,關(guān)注的細(xì)胞因子種類有限,且未報(bào)告低氧對(duì)BMSCs轉(zhuǎn)錄水平的影響;本研究未能縱向探討B(tài)MSCs細(xì)胞分泌上述細(xì)胞因子后如何通過旁分泌影響周圍心肌細(xì)胞NGF表達(dá),如何進(jìn)一步誘導(dǎo)心律失常發(fā)生。為此,本課題組正在進(jìn)行后續(xù)研究。最后,本研究未進(jìn)一步觀察不同氧濃度下BMSCs分泌各細(xì)胞因子水平是否存在差異,且觀察時(shí)間僅限于24 h內(nèi),未能橫向多層次比較,有待更多同行的進(jìn)一步合作與研究。
綜上所述,大鼠BMSCs細(xì)胞具有分泌神經(jīng)重構(gòu)相關(guān)因子ET-1、EPO、G-CSF的功能,低氧環(huán)境能顯著上調(diào)其上述細(xì)胞因子的表達(dá)水平,促進(jìn)心肌交感神經(jīng)重構(gòu),可能與BMSCs移植治療后心律失常副反應(yīng)的發(fā)生密切相關(guān)。
參考文獻(xiàn):
[1]Baldo MP,Davel AP,Nicoletti-Carvalho JE,et al.Granulocyte colony-stimulating factor reduces mortality by suppressing ventricular arrhythmias in acute phase of myocardial infarction in rats.[J].J Cardiovasc Pharmacol,2008,52(4):375-380.
[2]Gnecchi M,Melo LG.Bone marrow-derived mesenchymal stem cells: isolation,expansion,characterization,viral transduction,and production of conditioned medium[J].Methods Mol Biol,2009,482:281-294.
[3]Wen Z,Zheng S,Zhou C,et al.Repair mechanisms of bone marrow mesenchymal stem cells in myocardial infarction[J].J Cell Mol Med,2011,15(5):1032-1043.
[4]Zhang J,Wang LL,Du W,et al.Hepatocyte growth factor modification enhances the anti-arrhythmic properties of human bone marrow-derived mesenchymal stem cells[J].PLoS One,2014,9(10):e111246.
[5]Macia E,Boyden PA.Stem cell therapy is proarrhythmic[J].Circulation,2009,119(13):1814-1823.
[6]Lee JH,Kemp DM.Human adipose-derived stem cells display myogenic potential and perturbed function in hypoxic conditions[J].Biochem Biophys Res Commun,2006,341(3):882-888.
[7]Kolettis TM.Ventricular tachyarrhythmias during acute myocardial infarction:the role of endothelin-1[J].Life Sci,2014,118(2):136-140.
[8]Lee TM,Chen CC,Chang NC.Granulocyte colony-stimulating factor increases sympathetic reinnervation and the arrhythmogenic response to programmed electrical stimulation after myocardial infarction in rats[J].Am J Physiol Heart Circ Physiol,2009,297(2):H512-H522.
[9]Burdon TJ,Paul A,Noiseux N,et al.Bone marrow stem cell derived paracrine factors for regenerative medicine: current perspectives and therapeutic potential[J].Bone Marrow Res,2011,2011:207326.
[10]Maldonado-Soto AR,Oakley DH,Wichterle H,et al.Stem cells in the nervous system[J].Am J Phys Med Rehabil,2014,93(11 Suppl 3):S132-S144.
[11]Barton M,Yanagisawa M.Endothelin: 20 years from discovery to therapy[J].Can J Physiol Pharmacol,2008,86(8):485-498.
[12]Ieda M,F(xiàn)ukuda K,Hisaka Y,et al.Endothelin-1 regulates cardiac sympathetic innervation in the rodent heart by controlling nerve growth factor expression[J].J Clin Invest,2004,113(6):876-884.
[13]Bunn HF.Erythropoietin[J].Cold Spring Harb Perspect Med,2013,3(3):a11619.
[14]Kuhlmann MT,Kirchhof P,Klocke R,et al.G-CSF/SCF reduces inducible arrhythmias in the infarcted heart potentially via increased connexin43 expression and arteriogenesis[J].J Exp Med,2006,203(1):87-97.
[15]Wei L,F(xiàn)raser JL,Lu ZY,et al.Transplantation of hypoxia preconditioned bone marrow mesenchymal stem cells enhances angiogenesis and neurogenesis after cerebral ischemia in rats[J].Neurobiol Dis,2012,46(3):635-645.
[16]Tan H,Zhong Y,Shen X,et al.Erythropoietin promotes axonal regeneration after optic nerve crush in vivo by inhibition of RhoA/ROCK signaling pathway[J].Neuropharmacology,2012,63(6):1182-1190.
[17]Shim W,Mehta A,Lim SY,et al.G-CSF for stem cell therapy in acute myocardial infarction: friend or foe?[J].Cardiovasc Res,2011,89(1):20-30.
[18]Liu SP,Lee SD,Lee HT,et al.Granulocyte colony-stimulating factor activating HIF-1alpha acts synergistically with erythropoietin to promote tissue plasticity.[J].PLoS One,2010,5(4):e10093.
[19]Peterson KM,Aly A,Lerman A,et al.Improved survival of mesenchymal stromal cell after hypoxia preconditioning: role of oxidative stress[J].Life Sci,2011,88(1-2):65-73.
[20]Ren H,Cao Y,Zhao Q,et al.Proliferation and differentiation of bone marrow stromal cells under hypoxic conditions[J].Biochem Biophys Res Commun,2006,347(1):12-21.
[21]Zhou Y,Guan X,Wang H,et al.Hypoxia induces osteogenic/angiogenic responses of bone marrow-derived mesenchymal stromal cells seeded on bone-derived scaffolds via ERK1/2 and p38 pathways[J].Biotechnol Bioeng,2013,110(6):1794-1804.
(本文編輯郭懷印)
(收稿日期:2015-10-21)
中圖分類號(hào):R329
文獻(xiàn)標(biāo)識(shí)碼:A
doi:10.3969/j.issn.1672-1349.2016.06.013
文章編號(hào):1672-1349(2016)06-0599-05
通訊作者:伍衛(wèi),E-mail:wuwei9@mail.sysu.edu.cn
基金項(xiàng)目:廣東省醫(yī)學(xué)科研基金資助項(xiàng)目(No.B2013155)
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
中老年保健(2021年12期)2021-08-24 03:30:40
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中國傳媒大學(xué)學(xué)報(bào)(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科技傳播(2019年22期)2020-01-14 03:06:54
新世紀(jì)智能(英語備考)(2019年12期)2020-01-13 06:07:18
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年11期)2019-01-07 01:28:02
