一株拮抗香蕉枯萎病菌內(nèi)生細(xì)菌的抑菌機(jī)制研究
2016-05-30 20:16:11劉先寶蔡吉苗陳奕鵬黃貴修
熱帶農(nóng)業(yè)科學(xué) 2016年1期
劉先寶 蔡吉苗 陳奕鵬 黃貴修
摘 要 采用生物化學(xué)方法,分析了一株香蕉內(nèi)生細(xì)菌BEB99發(fā)酵液的乙酸乙酯粗提物對(duì)香蕉枯萎病菌4號(hào)小種(FOC4)的抑菌機(jī)制,結(jié)果表明,其對(duì)香蕉枯萎病菌菌絲脂質(zhì)過(guò)氧化、麥角甾醇含量、蛋白質(zhì)含量及果膠酶活性均有一定的影響,但不能產(chǎn)生幾丁質(zhì)酶。
關(guān)鍵詞 內(nèi)生細(xì)菌 ;勞爾氏菌 ;尖孢鐮刀菌;抑菌機(jī)制
分類號(hào) S432.4+2 Doi:10.12008/j.issn.1009-2196.2016.01.010
Abstract The antagonistic mechanisms of ethyl acetate crude extract of BEB99 strains for Fusarium oxysporium were studied by biological chemical method. The results revealed that ethyl acetate crude extract could influence lipid peroxidation, ergosterol content, protein content, the activity of PMG of Fusarium oxysporum. But it could not decompose the chitin.
Keywords Endophytic bacteria ; Ralstonia sp. ; Fusarium oxysporum ; antimicrobial mechanism
植物內(nèi)生細(xì)菌是指生活于植物的組織和器官內(nèi)部而不引起癥狀的細(xì)菌,其中很多菌種在病害生物防治和促進(jìn)植物生長(zhǎng)發(fā)育等方面具有潛在的應(yīng)用價(jià)值[1]。內(nèi)生細(xì)菌的生防機(jī)制主要包括競(jìng)爭(zhēng)作用、拮抗作用和誘導(dǎo)抗性,其中拮抗作用是生防菌主要的抗菌機(jī)制。生防菌產(chǎn)生的拮抗物質(zhì)主要包括細(xì)胞壁降解酶類,如幾丁質(zhì)酶或葡聚糖酶、抗菌蛋白、抗菌肽和揮發(fā)性物質(zhì)等[2]。
香蕉是中國(guó)重要的熱帶經(jīng)濟(jì)作物。由尖孢鐮刀菌古巴專化型引起的香蕉枯萎病是一種毀滅性的土傳病害。已有研究表明,利用內(nèi)生細(xì)菌生物防治香蕉枯萎病具有較好的防治效果[3]。本實(shí)驗(yàn)室以香蕉枯萎病菌為指示菌,經(jīng)篩選獲得一株具有抑菌活性的內(nèi)生細(xì)菌BEB99,該菌株經(jīng)鑒定為勞爾氏菌(Ralstonia sp.),其發(fā)酵液的石油醚和乙酸乙酯粗提物對(duì)香蕉枯萎病菌的菌絲生長(zhǎng)和孢子萌發(fā)均具有明顯的抑制作用,可造成病菌菌絲的畸形,抑制孢子萌發(fā)。為進(jìn)一步了解該菌對(duì)香蕉枯萎病菌的作用機(jī)制,筆者研究了該菌發(fā)酵液的乙酸乙酯粗提物對(duì)菌絲脂質(zhì)過(guò)氧化、麥角甾醇含量、蛋白質(zhì)含量及果膠酶活性的影響,以期為該菌株下一步的應(yīng)用研究奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 供試菌株
香蕉內(nèi)生細(xì)菌BEB99和香蕉枯萎病病菌尖孢鐮刀菌古巴專化型4號(hào)小種(Fusarium oxysporium f. sp. cubense (Smith) Snyder & Hansen race 4, FOC4)均由本實(shí)驗(yàn)室分離和保存。
1.1.2 培養(yǎng)基和試劑
NA培養(yǎng)基、優(yōu)化培養(yǎng)基(4 g/L葡萄糖,1 g/L酸水解酪蛋白,0.5 ml/L的1 mol/L MgCl2,1 ml/L的1 mol/L CaCl2,pH 6.5)、PDA培養(yǎng)基、PDB培養(yǎng)基、乙酸乙酯、幾丁質(zhì)培養(yǎng)基(膠狀幾丁質(zhì)15 g,酵母膏3 g,KH2PO4 1.36 g,NH4SO4 1 g,MgS04·5H2O 0.3 g,瓊脂15 g,蒸餾水1 000 mL)。
1.2 方法
1.2.1 BEB99菌株發(fā)酵液制備
將保存的內(nèi)生細(xì)菌BEB99菌株接種于優(yōu)化平板培養(yǎng)基中,于28℃中活化24 h;將活化的BEB99菌株接入50 mL優(yōu)化的液體培養(yǎng)基中(150 mL三角瓶),于28℃、180 r/min條件下振蕩培養(yǎng)36 h,制成種子液;再將種子液以7%的接種量接入優(yōu)化培養(yǎng)液中,于相同條件下培養(yǎng)36 h,制備BEB99菌株的發(fā)酵液。
1.2.2 BEB99菌株發(fā)酵產(chǎn)物分離
將濃縮的發(fā)酵產(chǎn)物用細(xì)胞超聲破碎儀破碎后,用250 mL乙酸乙酯有機(jī)溶劑與發(fā)酵液等體積混合,在1 000 mL分液漏斗中反復(fù)顛倒振蕩30 min,使其萃取充分,靜置過(guò)夜,萃取3次,合并有機(jī)相,將有機(jī)相減壓濃縮成浸膏,用乙酸乙酯溶劑定容,備用。
1.2.3 菌絲體處理方法
將FOC4分生孢子懸浮液接種于50 mL PDB中(分生孢子最終濃度為1×107 個(gè)/mL),于28 ℃靜置培養(yǎng)5 d;加入乙酸乙酯相提取物使其終濃度分別為50、100、150 μg/mL,繼續(xù)靜置培養(yǎng)5 d;用濾紙過(guò)濾備用。
1.2.4 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲脂質(zhì)過(guò)氧化影響測(cè)定
參照趙世杰等[4]的方法測(cè)定菌株BEB99乙酸乙酯相提取物對(duì)FOC4菌絲細(xì)胞膜脂質(zhì)過(guò)氧化的影響。
1.2.5 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲麥角甾醇影響測(cè)定
參考韓平等[5]的方法測(cè)定抑菌物質(zhì)對(duì)菌絲細(xì)胞膜組成成分麥角甾醇的影響。
1.2.6 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲蛋白質(zhì)含量影響測(cè)定
參照何璐[6]的方法測(cè)定BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲蛋白質(zhì)含量的影響
1.2.7 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4病菌胞外果膠酶活性影響測(cè)定
參照何璐[6]的方法測(cè)定BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4病菌果膠酶活性的影響。
1.2.8 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌株幾丁質(zhì)影響測(cè)定
將幾丁質(zhì)培養(yǎng)基倒平板后,把牛津杯插入平板中央,吸取20 μL乙酸乙酯相提取物加入牛津杯中,以乙酸乙酯溶劑為對(duì)照,3次重復(fù);于28℃中培養(yǎng)3 d,觀察牛津杯周圍是否有透明圈。
2 結(jié)果與分析
2.1 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲脂質(zhì)過(guò)氧化的影響
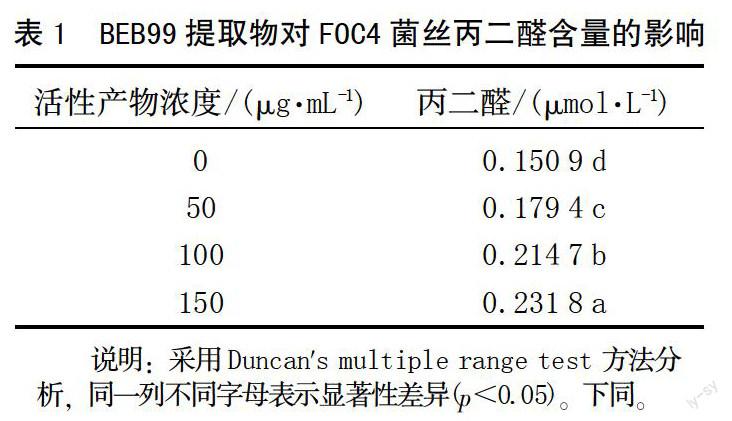
丙二醛(MDA)是細(xì)胞膜脂質(zhì)過(guò)氧化的產(chǎn)物,其含量高低表示細(xì)胞膜受損害程度。由表1可看出,經(jīng)菌株BEB99乙酸乙酯相提取物處理后,F(xiàn)OC4菌絲體細(xì)胞膜的脂質(zhì)過(guò)氧化產(chǎn)物丙二醛含量增加,均顯著高于對(duì)照,且不同處理間也存在顯著差異。隨著活性產(chǎn)物提取物濃度的升高,丙二醛含量逐漸上升,表明隨著抑菌物質(zhì)濃度的升高,病原菌細(xì)胞膜受到損害的程度越來(lái)越大。
2.2 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲麥角甾醇含量的影響
麥角甾醇是菌體細(xì)胞膜上的重要組成物質(zhì)。菌株BEB99乙酸乙酯相提取物對(duì)麥角甾醇含量的影響結(jié)果見(jiàn)表2,隨著活性產(chǎn)物提取物濃度的升高,麥角甾醇的含量不斷增加,顯著高于對(duì)照,并且不同處理間也存在顯著差異。說(shuō)明BEB99乙酸乙酯相提取物是抑制香蕉枯萎病菌的作用物質(zhì)之一。
2.3 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌絲蛋白質(zhì)含量的影響
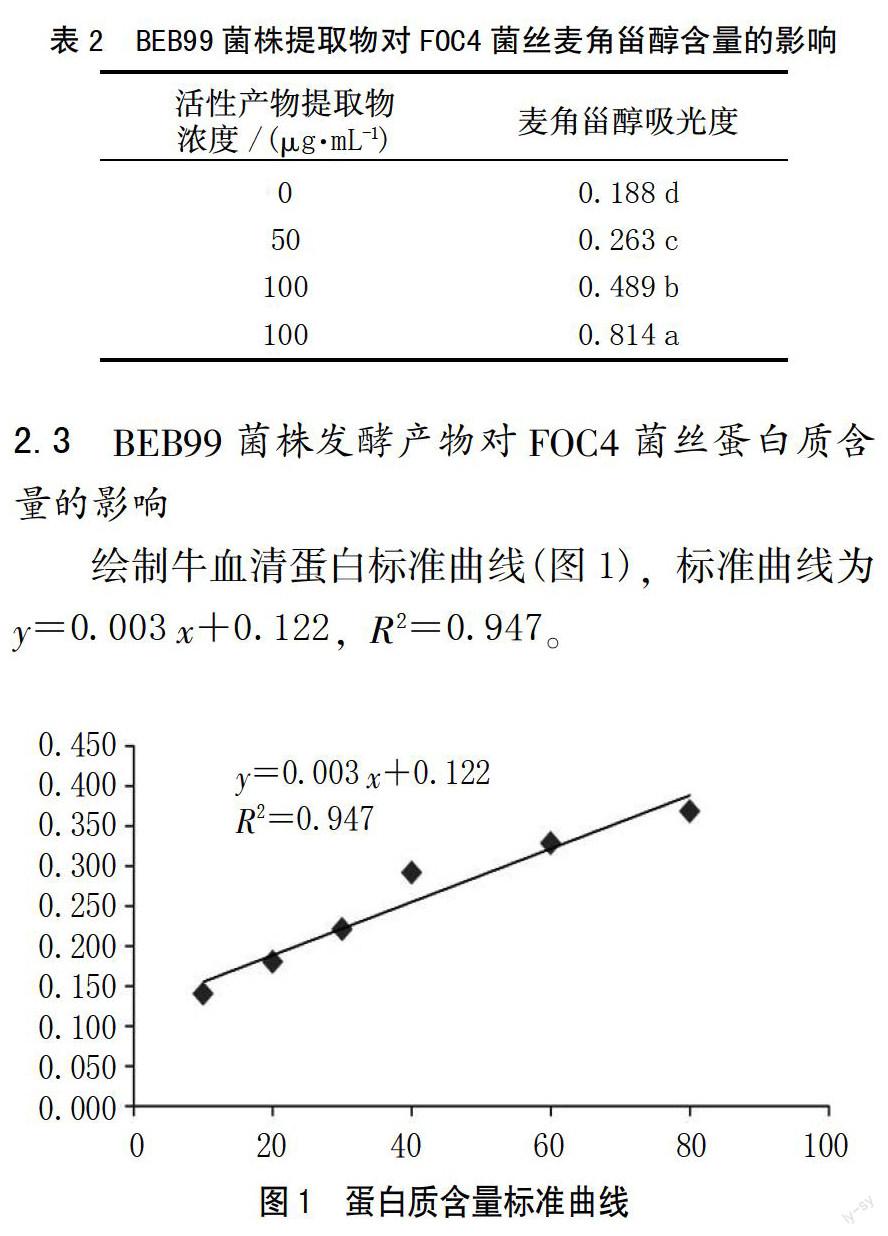
繪制牛血清蛋白標(biāo)準(zhǔn)曲線(圖1),標(biāo)準(zhǔn)曲線為y=0.003 x+0.122,R2=0.947。
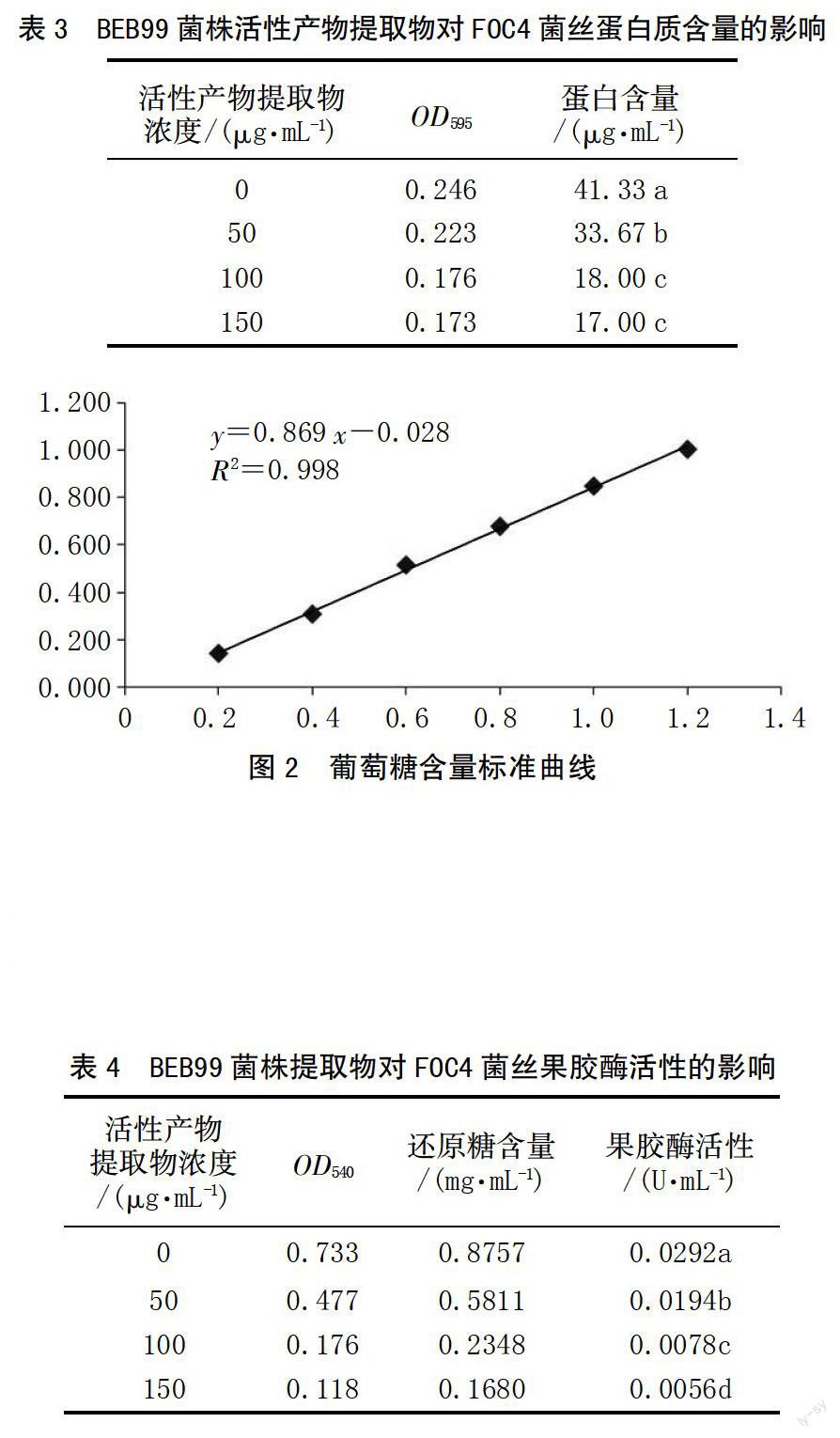
樣品溶液蛋白質(zhì)含量測(cè)定結(jié)果見(jiàn)表3,用不同濃度的菌株BEB99乙酸乙酯相提取物處理相同質(zhì)量的FOC4菌絲后,各處理樣品的蛋白質(zhì)含量隨活性產(chǎn)物提取物濃度的增大而降低,當(dāng)提取物濃度為100、150 μg/mL時(shí),蛋白質(zhì)的含量不存在顯著差異,但不同處理均與對(duì)照之間存在顯著差異,說(shuō)明提取物對(duì)FOC4菌絲中蛋白質(zhì)的含量影響顯著。
2.4 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌株胞外果膠酶活性的影響
繪制葡萄糖標(biāo)準(zhǔn)曲線(圖2),標(biāo)準(zhǔn)曲線為y=0.869 x-0.028,R2=0.998。
在540 nm處測(cè)得的OD值如表4所示,在葡萄糖標(biāo)準(zhǔn)曲線上查得還原糖含量,進(jìn)而計(jì)算出果膠酶的活性。各處理樣品的果膠酶活性隨乙酸乙酯相提取物濃度的增大而略有降低,且均顯著低于對(duì)照,同時(shí)不同處理間也存在顯著差異,說(shuō)明提取物對(duì)FOC4菌絲中果膠酶的合成存在顯著影響。
2.5 BEB99菌株發(fā)酵產(chǎn)物對(duì)FOC4菌株幾丁質(zhì)的影響
幾丁質(zhì)在培養(yǎng)基上呈白色不溶性微粒狀,在幾丁質(zhì)水解酶的作用下被水解成可溶性的小分子,呈透明狀。取20 μL乙酸乙酯相提取物注入牛津杯,在幾丁質(zhì)培養(yǎng)基上培養(yǎng)3 d后發(fā)現(xiàn),牛津杯周圍未出現(xiàn)透明的水解圈,即菌株BBE99乙酸乙酯相提取物不能水解病原菌細(xì)胞壁結(jié)構(gòu)的組成成分幾丁質(zhì)。
3 討論
目前,有關(guān)生防內(nèi)生菌抗菌機(jī)制的研究大部分僅限于其對(duì)病原菌活體的防效及其對(duì)菌絲生長(zhǎng)、孢子形成和萌發(fā)的影響,對(duì)于其毒理作用涉及較少[7]。本研究分析了菌株BEB99的乙酸乙酯相提取物對(duì)FOC4菌絲細(xì)胞組成和代謝的影響。脂質(zhì)和麥角甾醇是細(xì)胞膜的主要組成部分,脂質(zhì)過(guò)氧化反映了細(xì)胞膜受損害的程度,而丙二醛是脂質(zhì)過(guò)氧化的產(chǎn)物。研究發(fā)現(xiàn),隨著菌株BEB99的乙酸乙酯相提取物濃度的升高,細(xì)胞膜的脂質(zhì)過(guò)氧化增強(qiáng)(即脂質(zhì)過(guò)氧化產(chǎn)物丙二醛含量增加),麥角甾醇含量增加,表明損壞細(xì)胞膜結(jié)構(gòu)是菌株BEB99抑制香蕉枯萎病菌的作用方式之一,這與王芳[7]提出的作用機(jī)制(內(nèi)生枯草芽抱桿菌抗菌物質(zhì)與病原菌細(xì)胞膜上的卵磷脂類物質(zhì)結(jié)合形成復(fù)合體導(dǎo)致膜的穿孔,從而破壞串珠鐮刀菌質(zhì)膜)不同。幾丁質(zhì)是真菌細(xì)胞壁的重要組成物質(zhì)。本研究發(fā)現(xiàn),菌株BEB99乙酸乙酯相提取物不能產(chǎn)生幾丁質(zhì)酶來(lái)水解幾丁質(zhì)或者破壞幾丁質(zhì)的合成,而其對(duì)香蕉枯萎病菌細(xì)胞壁其他組成物質(zhì)是否有影響還需進(jìn)一步研究。
在菌株BEB99抗菌物質(zhì)影響病原菌菌體細(xì)胞內(nèi)代謝物質(zhì)的合成方面,研究了其對(duì)香蕉枯萎病菌菌絲體中蛋白質(zhì)含量的影響,結(jié)果表明,利用不同濃度的菌株BEB99乙酸乙酯相提取物處理相同質(zhì)量的FOC4菌絲后,各處理樣品的蛋白質(zhì)含量隨提取物濃度增大而降低,當(dāng)提取物濃度為100、150 μg/mL時(shí),蛋白質(zhì)的含量不存在顯著差異,但各處理均與對(duì)照之間存在顯著差異,說(shuō)明菌株BEB99乙酸乙酯相提取物對(duì)FOC4菌絲中蛋白質(zhì)的含量存在顯著影響。董章勇等[8]提出,果膠酶是香蕉枯萎病菌的主要致病毒力因子。本研究發(fā)現(xiàn),供試菌株的乙酸乙酯相提取物對(duì)香蕉枯萎病菌合成的果膠酶具有顯著影響,隨著其濃度的升高,果膠酶活性顯著降低,這將降低病原菌致病毒力,這與王芳[7]提出的的抗菌物質(zhì)對(duì)番茄葉霉病菌胞外酶活性的抑制效果相一致。綜合本研究的結(jié)果可知,菌株BEB99乙酸乙酯相提取物能夠抑制香蕉枯萎病病原菌的菌絲果膠酶活性和蛋白質(zhì)的含量,并通過(guò)破壞細(xì)胞膜結(jié)構(gòu)產(chǎn)生抑菌作用,而抗菌物質(zhì)對(duì)病菌菌絲細(xì)胞壁的組成物質(zhì)幾丁質(zhì)沒(méi)有影響,其作用機(jī)理的研究可為更加有效地應(yīng)用菌株BEB99奠定基礎(chǔ),在實(shí)踐上具有重要的應(yīng)用價(jià)值,值得進(jìn)一步開(kāi)發(fā)利用。
參考文獻(xiàn)
[1] Jesús Mercado-Blanco, Ben J J. Lugtenberg. Biotechnological Applications of Bacterial Endophytes[J]. Current Biotechnology, 2014, 3(3): 60-75.
[2] Ryan R P, Germaine K, Franks A, et al. Bacterial endophytes: recent developments andapplications[J]. FEMS Microbiol Lett, 2008, 278(1): 1-9.
[3] Ting A S Y, Meon S, Kadir J, et al. Endophytic microorganisms as potential growth promoters of banana[J]. BioControl, 2008, 53(3): 541-553.
[4] 趙世杰,許長(zhǎng)成,鄒 琦,等. 植物組織中丙二醛測(cè)定方法的改進(jìn)[J]. 植物生理學(xué)通訊,1991,30(3):207-210.
[5] 韓 平,劉西莉,劉鵬飛,等. 高效液相色譜法研究啶菌惡唑?qū)Ψ鸦颐共【溄晴薮忌锖铣傻挠绊慬J]. 分析化學(xué),2006,10(2):78-82.
[6] 何 璐. 苦參內(nèi)生真菌抑菌物質(zhì)純化鑒定及作用機(jī)制研究[D]. 沈陽(yáng): 沈陽(yáng)農(nóng)業(yè)大學(xué), 2011.
[7] 王 芳. 苦參內(nèi)生拮抗細(xì)菌篩選及其抗菌物質(zhì)純化鑒定[D]. 沈陽(yáng):沈陽(yáng)農(nóng)業(yè)大學(xué),2009.
[8] 董章勇,王 琪,秦世雯,等. 香蕉枯萎病菌1號(hào)和4號(hào)生理小種細(xì)胞壁降解酶的比較[J]. 植物病理學(xué)報(bào),2010,40(5):463-468.
