馬氏珠母貝肌肉生長抑制素基因eSNP多態性與生長性狀的關聯分析
2016-04-27 02:10:37李琴林堅士何毛賢
海洋通報 2016年1期
黃 文,李琴,林堅士,何毛賢
(1.中國科學院南海海洋研究所 中國科學院熱帶海洋生物資源與生態重點實驗室 廣東省應用海洋生物學重點實驗室,廣東 廣州 510301;2.中國科學院大學,北京 100039)
?
馬氏珠母貝肌肉生長抑制素基因eSNP多態性與生長性狀的關聯分析
黃文1,2,李琴1,2,林堅士1,何毛賢1
(1.中國科學院南海海洋研究所中國科學院熱帶海洋生物資源與生態重點實驗室廣東省應用海洋生物學重點實驗室,廣東廣州510301;2.中國科學院大學,北京100039)
摘要:肌肉生長抑制素myostatin(MSTN),是目前已知抑制肌肉生長最強效的分化因子,具有負向調控肌肉生長、發育的作用,它與生長之間的這種單向作用關系,使其在養殖生物遺傳改良上具有重要的潛在應用價值。文以馬氏珠母貝MSTN基因21個eSNP位點作為候選SNP位點,對106只馬氏珠母貝進行基因分型并與8個生長性狀進行關聯分析。SNP最小等位基因頻率的范圍為0.028 3~0.490 4,多態信息含量(PIC)的范圍為0.093 6~0.375 0,觀測雜合度(Ho)及期望雜合度(He)的范圍分別為0.028 3~1.000 0及0.098 9~0.502 7。2個SNP位點與生長性狀顯著關聯:g.3154 A>G位點的GG型個體的殼高、殼長、總重、殼重、軟體部重和閉殼肌重顯著大于AG型個體(P<0.05);g.4379A>G位點GG型個體的殼高、殼長、鉸合線長、總重、軟體部重和閉殼肌重顯著大于AG型個體(P<0.05)。g.3154 A>G位點和g.4379A>G位點間無顯著交互作用。這2個SNP可作為潛在的馬氏珠母貝基因型選擇育種標記。
關鍵詞:馬氏珠母貝;肌肉生長抑制素MSTN;單核苷酸多態性;生長性狀;關聯分析
目前在水產動物育種領域,研究生長性狀相關基因主要有兩種方法:基因組測序篩選檢測或稱QTL定位法和候選基因法(candidate gene approach)。其中,候選基因法因具有目標明確,檢驗效率高,且有費用低廉、對群體要求較低、操作簡便等優點,廣泛應用在研究性狀關聯分子標記中,如W ang等(2012)通過對MDA5(黑色素瘤分化相關基因-5)的5’側翼區中的SNP與草魚對GCRV疾病的易感性/抗性進行關聯分析,確定了與抗性相關的草魚(Ctenopharyngodon idella)的基因型,魏可鵬等(2012)克隆了建鯉(Cyprinus carpio)IGFBP1基因,并成功篩選到了兩個與增重顯著相關的SNPs位點。
肌肉生長抑制素myostatin(MSTN),是目前已知抑制肌肉生長最強效的分化因子,具有負向調控肌肉生長、發育的作用,它與生長之間的這種單向作用關系,使其在養殖生物遺傳改良上具有重要的潛在應用價值。近年來,myostatin基因及其SNP與多種經濟性狀之間的顯著關聯已在雞(Gallus domesticus)(McFarland et al,2007)、馬(Equus caballus)(Li et al,2014)、大口黑鱸(Micropterus salmoide)(于凌云等,2010)和扇貝(Chlamys farreri;Argopecten irradians)(Hu et al,2010;Guo et al,2012)等多個物種中被鑒定,而這種關聯性可以為日后開展分子標記輔助育種奠定基礎。
馬氏珠母貝(Pinctada fucata)是一種重要的海水養殖貝類,利用分子標記輔助育種可加快馬氏珠母貝育種進程。本研究中以馬氏珠母貝myostatin基因作為生長性狀相關的候選基因,檢測基因外顯子上的SNP位點(eSNP),并對其基因型與生長性狀進行關聯分析,同時分析了SNP位點間的聚合效應,以期為馬氏珠母貝分子標記輔助育種提供潛在的SNP標記。
1材料與方法
1.1實驗樣品
本實驗所用的馬氏珠母貝(Pinctada fucata)來源于以生長性狀為選擇指標的混合選擇群體第三代,其基礎群體為深圳野生群體,養殖于深圳大鵬澳海區。隨機從該群體(8月齡)中取106只貝,測量并記錄每只貝的殼高、殼長、鉸合線長、殼寬、總重、殼重、軟體部重和閉殼肌重,其均值分別為48.45±6.13 mm、46.72±5.92 mm、40.48± 4.39mm、17.28±1.90mm、17.04±5.18 g、9.71± 2.79 g、5.15±1.97 g、0.83±0.39 g。取每只貝的閉殼肌組織于95%乙醇中,-20°C保存。
1.2基因組DNA的提取
使用HiPure Universal DNA Mini Kit提取閉殼肌組織的DNA,用1.0%的瓊脂糖凝膠電泳檢測其完整性,紫外分光光度計測量DNA樣品A260/ A280的比值來檢測純度,提取的DNA于-20°C保存。
1.3 SNP位點分型
用于分型用的21個外顯子SNP(eSNP)位點來源于Li等(2014)鑒定的馬氏珠母貝MSTN基因,并根據其在基因組DNA的位置重新命名。利用Tetra-Primer Arms在線引物設計程序(http:// primer1.soton.ac.uk/primer1.html)設計SNP位點的基因分型引物(表1)。Tetra-Primer PCR分型引物包括兩條內引物FI、RI及兩條外引物FO、RO,其中兩條內引物3'端倒數第三個堿基引入一個錯配堿基以增加特異性。當檢測樣品為雜合型時電泳結果理論上應該有三條帶,包括兩條外引物擴增的一條帶(FI、RI),內引物和外引物擴增的兩條帶(FI、RO)以及(RI、FO);當檢測樣品為純合型時則為兩條帶,包括兩條外引物擴增的一條帶(FI、RI),內引物與外引物擴增條帶中的一條。PCR反應體系為:2×PCR Mix 2.0μL,DNA Taq酶0.125μL,引物(10μM)0.5μL,基因組DNA 0.4μL,ddH2O 3.725μL;擴增程序為:94℃預變性4min,1個循環;94°C變性35 s,最適退火溫度退火40 s,72℃延伸40s,35個循環;72℃延伸10min,1個循環。PCR產物用3.0%的瓊脂糖凝膠電泳檢測,觀察電泳圖譜(圖1),確定每個個體的基因型。
1.4 SNP與生長性狀的關聯分析
統計群體中SNP堿基置換類型及各種類型的數量。利用PIC_CALC軟件計算群體中每個SNP位點的多態信息含量(polymorphism content information,PIC)。利用Popgene軟件(Yeh etal,1997)計算每個SNP位點在群體中的等位基因頻率、基因型頻率、有效等位基因數(Ne)、期望雜合度(expected heterozygosity,He)和觀測雜合度(observed heterozygosity,Ho),并進行哈德-溫伯格平衡(Hardy-W einberg equilibrium,HW E)檢驗,SHEsis軟件分析連鎖不平衡。由于總體分布未知,因此對于106只貝生長性狀正態分布進行檢驗,采用非參數檢驗中的單樣本Kolmogorov-Smirnov檢驗,計算結果顯示8個性狀的P值均大于0.05,即符合正態分布。采用SPSS 17.0軟件中的GLM程序對21個SNP位點與各生長性狀進行關聯分析,得到顯著關聯生長的SNP位點(P<0.05),并檢驗兩位點間是否存在交互作用;對于顯著關聯生長性狀SNP位點經Levene檢驗結果P值均大于0.05,表明方差齊次,然后采用Sidak法比較g.3154A>G位點不同基因型間的性狀差異,對于g.4379A>G位點采用獨立樣本t檢驗比較(P<0.05)。
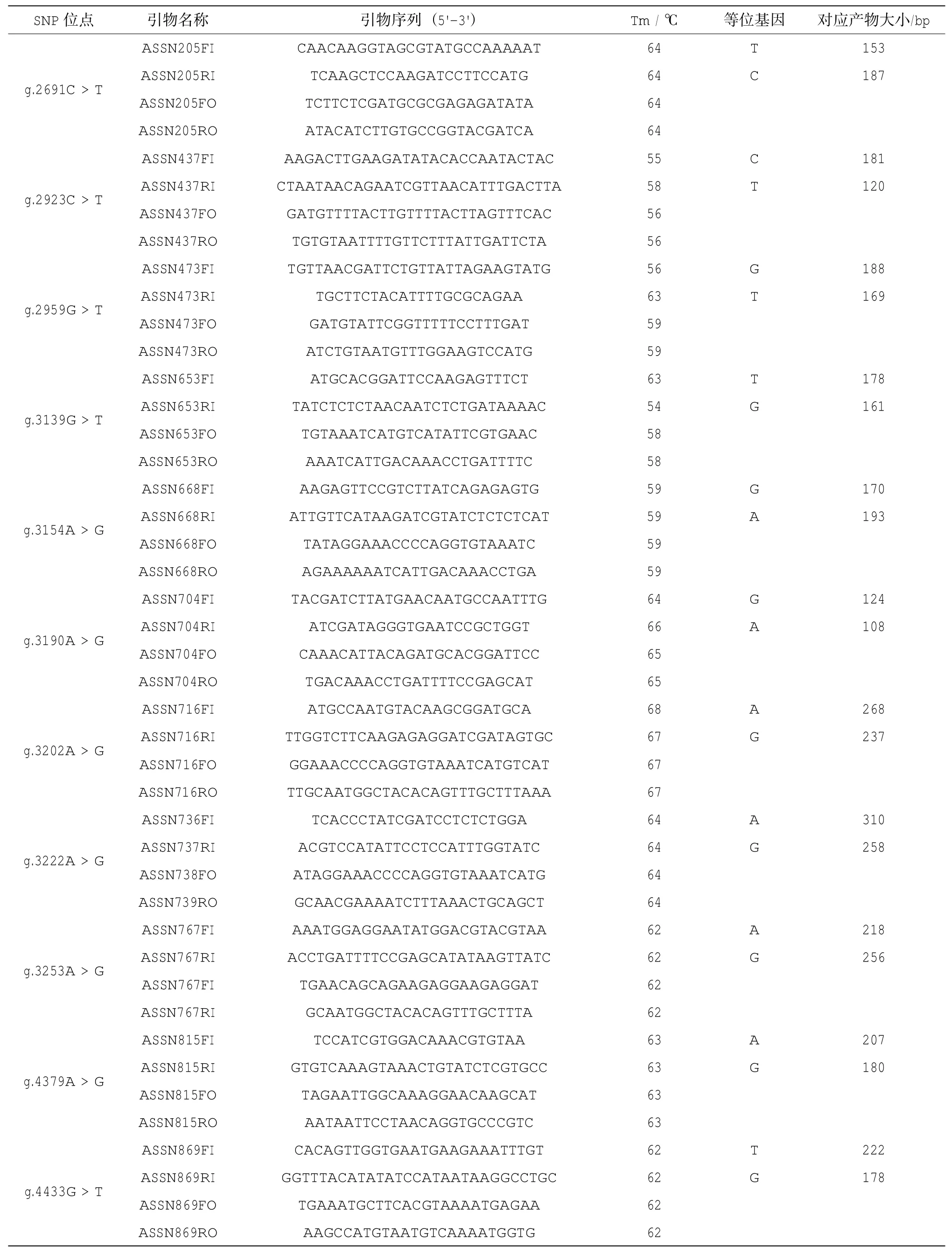
表1 21個SNP位點分型引物
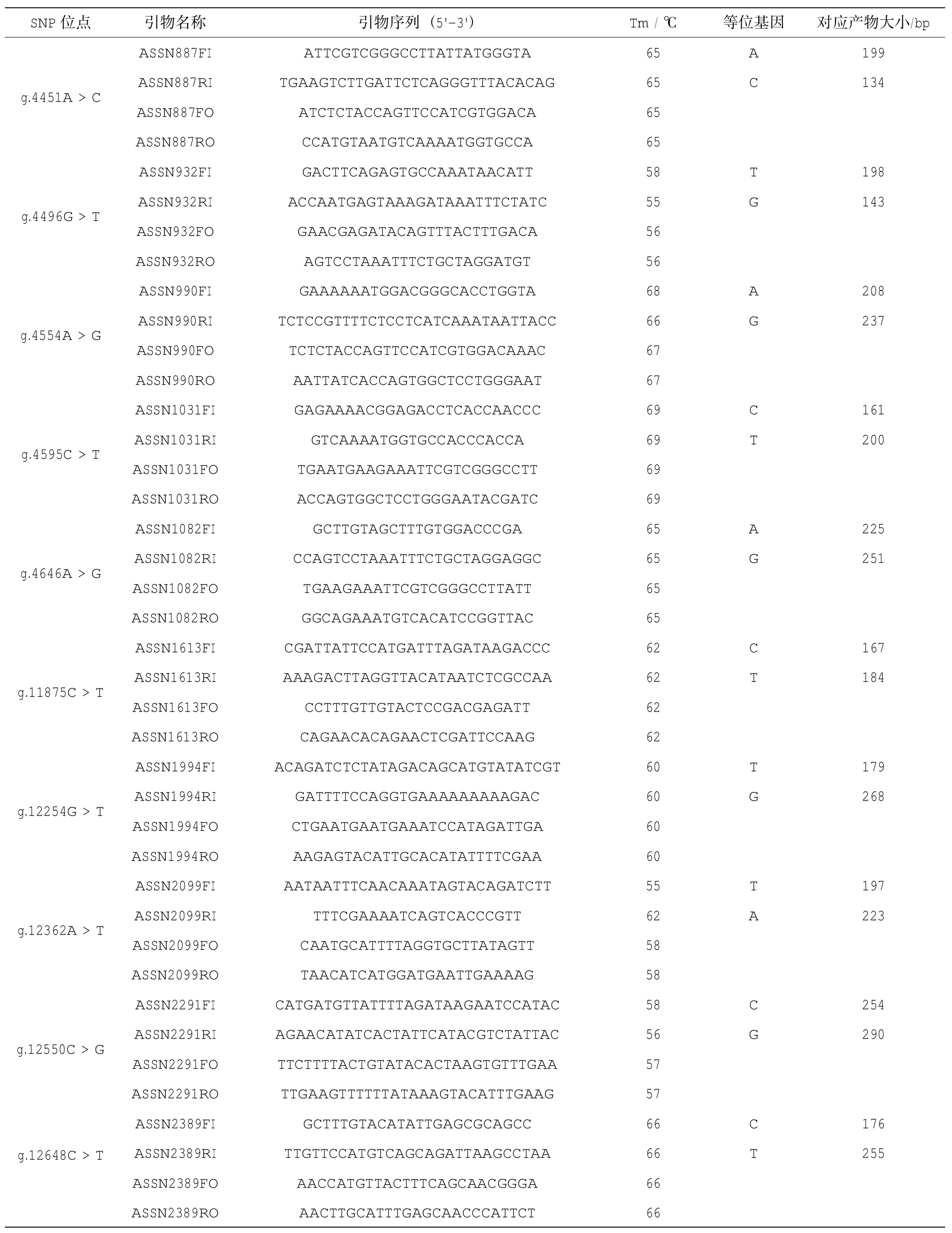
續表1 21個SNP位點分型引物
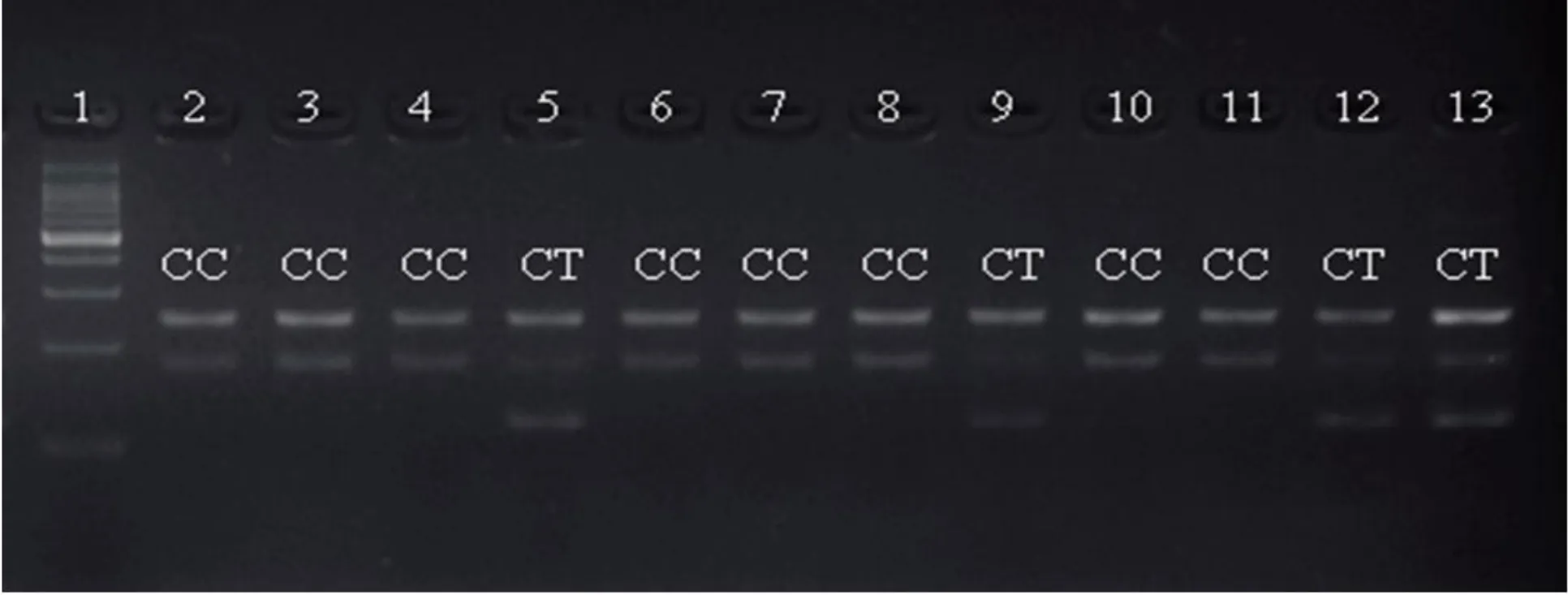
圖1 部分個體在位點SNP g.2923C>T的分型結果(1.GenStar direct-loadTM 100bp DNA LadderMarker;2-13:每個個體的基因型,CC表示純合子,CT表示雜合子)
2實驗結果
2.1 21個SNP位點的遺傳參數統計
在這21個SNP位點中,最常見的突變類型為A/G轉換,共有8個位點屬于此類,占SNP位點總數的38.1%;其次為G/T顛換和C/T轉換,各包括5個SNP位點,分別占位點總數的23.8%。除g.3222A >G、g.4595C>T檢測到一種基因型,g.4379A>G、g.12648C>T檢測到兩種基因型,其他位點3種基因型均能檢出。群體中最小等位基因頻率(minorallele frequency,MAF)的范圍為0.0519~ 0.500 0,多態信息含量(PIC)的范圍為0.093 6~ 0.375 0,觀測雜合度(Ho)及期望雜合度(He)的范圍分別為0.028 3~1.000 0及0.098 9~0.502 7(表2)。哈德-溫伯格平衡(HW E)檢驗結果表明,21個SNP中有8個符合平衡(表3)。
2.2 SNP與生長性狀的關聯分析及均值比較分析
經GLM分析,在這21個SNP位點中,只有g.3154A>G、g.4379A>G這2個位點與生長性狀顯著相關(P<0.05)(表4-5)。進一步分析表明,對于g.3154A>G位點,GG型個體的殼高、殼長、總重、殼重、軟體部重和閉殼肌重顯著大于AG型個體(P<0.05),分別大10.99%、9.08%、26.12%、26.65%、35.28%和43.33%,AA型個體除殼寬顯著大于AG型個體(P<0.05),其他性狀均與另外兩種基因型無顯著差異;g.4379A>G位點,GG型個體的殼高、殼長、鉸合線長、總重、軟體部重和閉殼肌重顯著大于AG型個體(P<0.05),分別大6.20%、6.96%、5.65%、13.58%、18.01%、27.40%(表6)。
對兩個與軟體部重和閉殼肌重都顯著關聯的位點(g.3154A >G位點和g.4379A >G)用SHEsis軟件連鎖不平衡分析,表明這兩個位點連鎖不緊密(D'= 0.26和R2= 0.04),用GLM做兩因素交互分析,未發現兩位點存在顯著交互作用(表7)。

表2 馬氏珠母貝M STN基因21個eSNP位點的基因型頻率及等位基因頻率

表3 馬氏珠母貝M STN 21個eSNP位點的群體遺傳參數
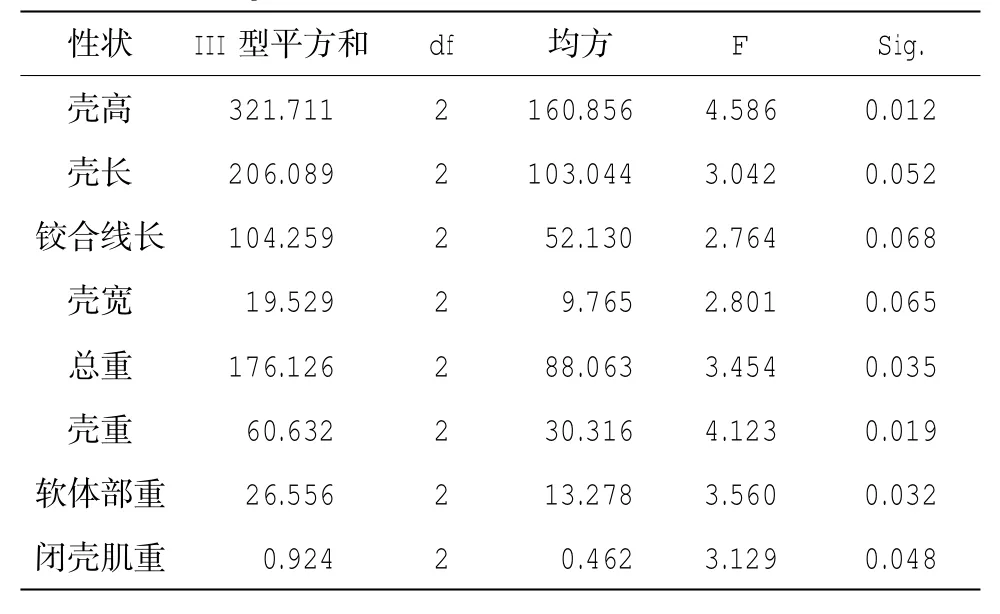
表4 g.3154A>G位點與生長性狀的關聯性分析
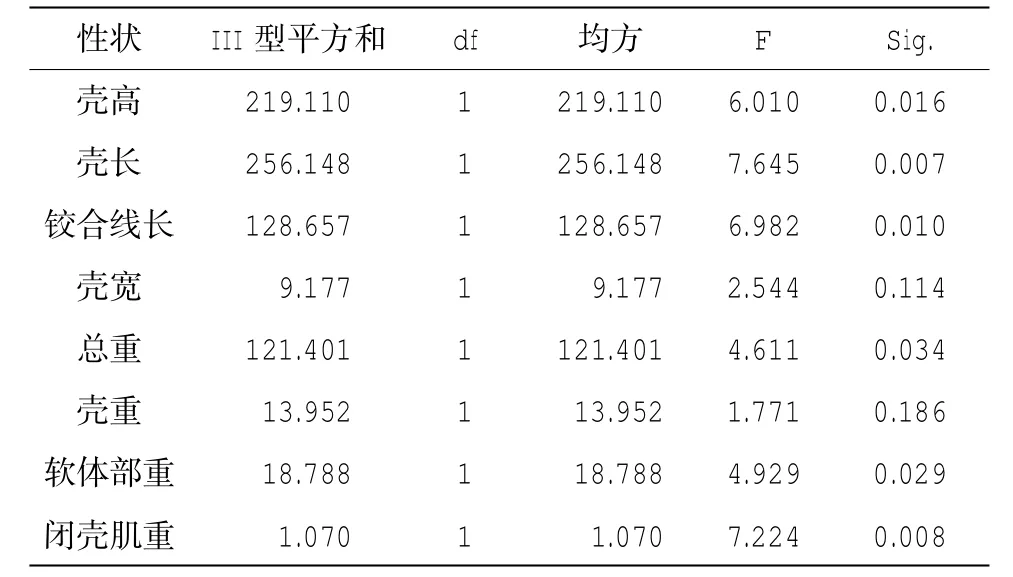
表5 g.4379A>G位點與生長性狀的關聯性分析

表6 兩個顯著關聯生長性狀的SNP位點的均值比較分析
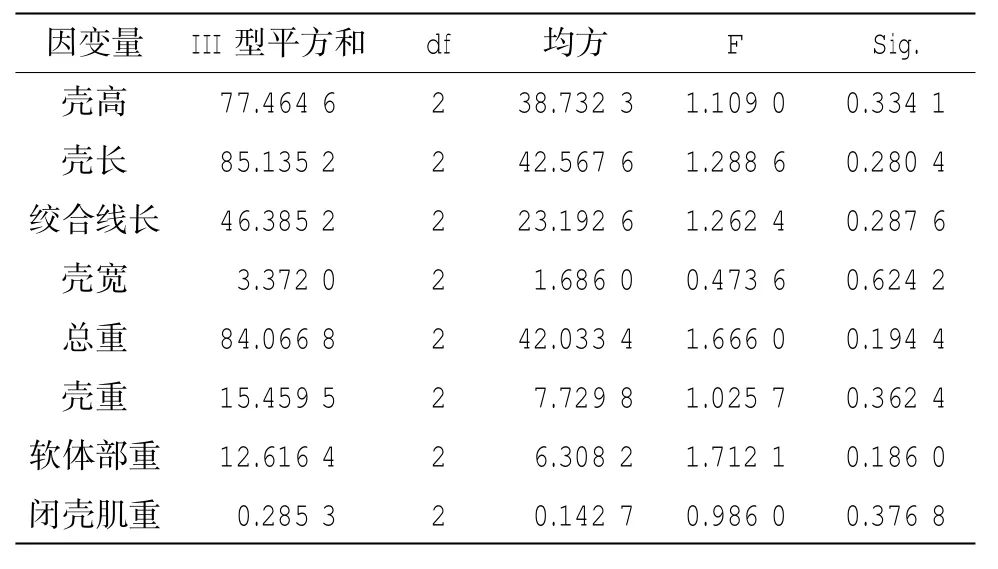
表7 g.3154A>G位點和g.4379A>G位點交互作用分析
3討論
有研究表明SNP位點在不同群體中,分布情況具有差異性。Morales-Colli'o等(2014)在野生群體和養殖群體中對紫扇貝(Argopecten purpuratus)基因上的三個SNP位點進行分型,結果發現,一個位于5’-UTR的SNP和一個位于編碼區的SNP均只出現在野生群體中,另一個位于3'-UTR 的SNP卻只出現在養殖群體中。Sauvage等(2007)在太平洋牡蠣(Crassostrea gigas)中發現編碼區每60個堿基有一個SNP位點,而非編碼區每40個堿基就有一個,表明非編碼區還存在大量SNP位點。而在本實驗中,我們用于分型的群體僅來自于一個混合選擇群體,屬于人工選育群體;且SNP位點全部來自外顯子區域,這些可能是導致部分SNP位點基因型未被全部檢出的主要原因。
在馬氏珠母貝MSTN的外顯子中,檢測出21個多態SNP位點,明顯高于Guo等(2011)在海灣扇貝(Argopecten irradians)MSTN基因外顯子上確定的兩個SNP位點,而Nú?ez-Acu?a等(2014)則在貽貝(Mytilus chilensis)中檢測到37個SNP位點,說明該基因的多態性水平存在物種間差異。將這21個SNPs的遺傳參數,如觀測雜合度(Ho)、期望雜合度(He),與Huang等(2014)獲得的馬氏珠母貝SNPs結果比較,發現其雜合度和期望雜合度較其高,說明馬氏珠母貝MSTN基因外顯子堿基變異水平較高。
為了探索這些SNP與性狀之間是否存在某些關聯性,對SNP基因型和表型進行了統計學分析。GLM程序分析結果顯示21個SNP位點中有兩個位點與某些生長性狀顯著關聯。從統計結果上看,g.3154A>G位點GG型個體的殼高、殼長、總重、殼重、軟體部重及閉殼肌重大于AG型基因型個體;g.4379A>G位點GG型個體的殼高、殼長、絞合線長、總重、軟體部重及閉殼肌重大于AG型基因型個體。對于這兩個SNP位點,GG型都為優異基因型。同時,這兩個位點與馬氏珠母貝的軟體部重及閉殼肌重顯著關聯,因而這兩個位點可能與myostatin基因的表達密切相關。根據前面的分析,GG型為優異基因型,我們推測GG型個體myostatin基因的表達較其他基因型個體較低。類似的研究在扇貝中也有報道,如W ang等(2010)對櫛孔扇貝(Chlamys farreri)基因編碼區的SNP進行鑒定,并將其基因型與生長性狀進行關聯分析,發現Cf-MSTN存在兩個SNPs,分別位于外顯子2 和3上,外顯子2上的SNP的GG基因型個體在體重、軟體部重、閉殼肌重、殼長、殼高各項生長指標上也均優于基因型AA和AG的個體,同時還發現1個SNP位點會引起氨基酸變化。在本研究中,g.3154A>G位點和g.4379A>G位點雖位于Pf-MSTN cDNA的開放閱讀框內,但不會引起氨基酸的改變,屬于同義突變。然而,近年來很多研究表明,同義突變也能調控基因的轉錄和翻譯,Greenwood等(2003)發現啟動子和內含子同義突變均能夠影響許多基因的轉錄效率,Kimchi-Sarfaty等(2007)發現MDR1基因中的一個同義突變可以改變蛋白的空間結構;Capon等(2004)發現角膜鎖鏈蛋白基因中的同義突變能提高mRNA分子的穩定性。因此,將來對這類SNP繼續進行深入研究很有必要,這有助于闡明myostatin基因變異對馬氏珠母貝生長性狀的影響。
優異基因的聚合能得到理想的累加效應,基因聚合最早由Yadav等(1990)研究改良芥菜(Brassica juncea)抗逆和抗病性狀時提出。目前,基因聚合效應已經廣泛應用在研究農作物和禽畜的經濟性狀中,而在水產育種方面報道較少。束婧婷(2007)統計不同品種雞(Gallus domesticus)ADSL和ARS-AIRS-GART基因的多態性,并分析不同基因型組合對胸肌IMP(肌苷酸)含量的影響,結果表明,TTTT組合基因型個體的IMP含量顯著高于CCCC型個體(P<0.05)。陳磊(2013)研究了大黑口鱸(Micropterus salmoide)8個分子標記的基因聚合效應,發現隨著優異基因型數量的增加,平均體重同步遞增,而位于MSTN上的優異基因型對體重的貢獻率最大。基因型的聚合效應并不是各基因型效應所造成的表型的簡單相加,而且純合基因型互相組合具有顯著的累加效應,能夠穩定的遺傳給子代。在本研究中,g.3154A>G位點和g.4379A>G位點并不存在交互作用,GGGG型個體在軟體部重和閉殼肌重具有最大均值,但較其它基因型組合差異并不顯著,因而這兩個優異基因型不存在明顯聚合效應。
雖然未檢測到聚合效應,但在本研究中篩選到的顯著關聯的SNP位點仍可作為馬氏珠母貝基因型選擇育種的潛在分子標記,這些標記的育種效應將在后續的研究中加以驗證。
參考文獻
Capon F,Allen M H,Ameen M,et al,2004.A synonymous SNP of the corneodesmosin gene leads to increasedmRNA stability and demonstratesassociation with psoriasisacross diverse ethnic groups.Human MolecularGenetics,13 (20):2361-2368.
Greenwood T A,Kelsoe JR,2003.Promoter and intronic variants affect the transcriptional regulation of the human dopamine transporter gene.Genomics,82 (5):511-520.
Guo L,Li L,Zhang S,etal,2011.Novel polymorphisms in themyostatin geneand theirassociationwith growth traits in a variety ofbay scallop,Argopecten irradians.AnimalGenetics,42 (3):339-340.
Guo L,Li L,Zhang S,etal,2012.Molecular cloning and characterization of the myostatin gene in a cultivated variety of bay scallop,Argopecten irradians.Aquaculture,350-353:192-199.
Hu X,Guo H,He Y,et al,2010.Molecular characterization ofmyostatin gene from Zhikong scallop Chlamys farreri(Jones et Preston 1904).Genes& Genetic Systems,85 (3):207-218.
Huang X D,W u SZ,Guan Y Y,etal,2014.Identification ofsixteen single-nucleotide polymorphism markers in the pearl oyster,Pinctada fucata,for population genetic structureanalysis.JournalofGenetics,93 (1):e1-e4.
Kimchi-Sarfaty C,Oh JM,Kim IW,et al,2007.A“silent”polymorphism in theMDR1 gene changessubstrate specificity.Science,315 (5811):525-528.
LiQ,Li Y G,Huang X D,et al,2014.Molecular cloning and single nucleotide polymorphisms identification of myostatin cDNA in the pearl oyster,Pinctada fucata.Journal of theW orld Aquaculture Society,45 (6):638-651.
LiR,Liu D H,Cao C N,etal,2014.Single nucleotide polymorphisms of myostatin gene in Chinese domestic horses.Gene,538 (1) :150-154.
McFarland D C,Velleman S G,Pesall JE,et al,2007.The role ofmyostatin in chicken (Gallusdomesticus)myogenic satellite cellproliferation and differentiation.Generaland Comparative Endocrinology,151 (3):351-357.
Morales-Collío K,Valenzuela-Mun~oz V,Gallardo-Escárate C,2014.The myostatin gene of Argopecten purpuratus (APMSTN) :transcript expression and single-nucleotide polymorphism differencesbetween wild and hatchery-bred populations.Journal of Molluscan Studies,80 (2):169-176.
Nún~ez-Acun~a G,Gallardo-Escárate C,2014.The myostatin gene of Mytilus chilensis evidences a high level of polymorphism and ubiquitous transcriptexpression.Gene,536 (1):207-212.
Sauvage C,Bierne N,Lapègue S,etal,2007.Single Nucleotide polymorphismsand their relationship to codon usagebias in the Pacific oyster Crassostreagigas.Gene,406 (1-2):13-22.
W ang L,Su JG,Yang C R,et al,2012.Genomic organization,promoter activityofgrass carp MDA5 and theassociation of itspolymorphisms with susceptibility/resistance to grass carp reovirus.Molecular Immunology,50,236-243.
W ang X,Meng X,Song B,etal,2010.SNPs in themyostatin gene of the mollusk Chlamys farreri:Associationwith growth traits.Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,155 (3):327-330.
Yadav R D,Singh SB,RaiM,etal,1990.Gene pyramiding and horizontal resistance to diara stress inmustards.Journal National Academy Science Letters,13 (9):325-327.
Yeh F C,1997.Population genetic analysis of codominant and dominant markersand quantitative traits.Belgian JournalofBotany,129,157.
陳磊,2013.大口黑鱸生長相關優勢基因型聚合效果研究.碩士學位論文.上海:上海海洋大學.
束婧婷,吉文林,包文斌,等,2007.雞ADSL基因和GARSAIRS-GART基因對雞肉肌酸(IMP)含量的影響.畜牧獸醫學報,38(8):786-791.
魏可鵬,俞菊華,李紅霞,等,2012.建鯉IGFBP1基因的克隆及與增重相關的SNP位點分析.華北農學報,27(3):75-80.
于凌云,白俊杰,樊佳佳,等,2010.大口黑鱸肌肉生長抑制素基因單核苷酸多態性位點的篩選及其與生長性狀關聯性分析.水產學報,34(6):845-851.
(本文編輯:袁澤軼)
Singlenucleotide polym orphism sofm yostatin geneand itsassociation w ith grow th traits in the pearloyster,Pinctada fucata
HUANG W en1,2,LIQin1,2,LIN Jian-shi1,HE Mao-xian1
(1.CASKey LaboratoryofTropicalMarine Bio-resourcesand Ecology,Guangdong ProvincialKey LaboratoryofApplied Marine Biology,South China Sea InstituteofOceanology,Chinese AcademyofSciences,Guangzhou 510301,China;2.UniversityofChineseAcademyof Sciences,Beijing100049,China)
Abstract:Myostatin (MSTN),known as themostpowerfulgrowth differentiation factor,playsa crucial role in the negative regulation ofmuscle growth and development.The existence ofa single genewith such amajor effecton themuscle growth makes ithave the potentialvalue in the genetic improvementofanimals.In thisstudy,we used 21 eSNPs from myostatin gene to divide the genotypes of 106 individuals from a breeding stock of pearl oyster Pinctada fucata and made the correlation analysiswith the eightgrowth traits.The results show that theminorallele frequency (MAF) and polymorphism information content(PIC) are in the range of0.028 3 to 0.490 4 and 0.093 6 to 0.375 0,and the observed and expected heterozygosities range from 0.028 3 to 1.000 and 0.098 9 to 0.502 7,respectively.Two SNPsare significantly associated with growth traits:for the g.3154A>G site,individualswith GG type are significantly greater than thosewith AG type in the shellheight,shell length,totalweight,shellweight,soft tissueweightand adductormuscleweight(P<0.05);for the g.4379A>G site,oneswith GG type are significantly greater than thosewith AG type in the shellheight,shell length,hinge line length,totalweight,soft tissue weight and adductormuscle weight(P<0.05) .There is no significant interaction between g.3154A>G site and g.book=89,ebook=924379A>G site.These two SNPs could be used as the potentialmolecularmarkers in the genotype-selective breeding for P.fucata.
Keywords:Pinctada fucata;myostatin;single-nucleotide polymorphism;growth traits;association analysis
通訊作者:何毛賢,研究員,電子郵箱:hmx@scsio.ac.cn。
作者簡介:黃文,男,碩士研究生,主要從事馬氏珠母貝分子遺傳育種研究,電子郵箱:94214460@qq.com。
基金項目:國家863計劃(2012AA10A410);廣東省科技計劃項目(2013B020308005)。
收稿日期:2015-01-30;
修訂日期:2015-03-19
Doi:10.11840/j.issn.1001-6392.2016.01.012
中圖分類號:S968.31
文獻標識碼:A
文章編號:1001-6932(2016)01-0088-08
