大鼠PAX6基因真核表達載體構建及鑒定*
2016-04-25 05:13:47嚴會文廖文萍閆麗麗
貴州醫科大學學報 2016年3期
嚴會文, 蘇 敏, 高 杰, 廖文萍, 閆麗麗, 黃 悅*
(貴州醫科大學 組織學與胚胎學教研室, 貴州 貴陽 550004)
?
大鼠PAX6基因真核表達載體構建及鑒定*
嚴會文**, 蘇敏, 高杰, 廖文萍, 閆麗麗, 黃悅***
(貴州醫科大學 組織學與胚胎學教研室, 貴州 貴陽550004)
[摘要]目的: 構建攜帶綠色熒光蛋白(GFP)報告基因及PAX6基因的重組真核表達載體,觀察其在人胚腎細胞(293FT)細胞中的表達。方法: 采用聚合酶鏈式反應(PCR)從大鼠腦組織中獲取PAX6基因,經連接T載體測序驗證正確后,與真核表達載體pEF1α-IRES-AcGFP經SalI/BamHI雙酶切后;經T4 DNA連接酶連接,獲得重組質粒pEF1α-PAX6-IRES-AcGFP,用菌液PCR、SalI/BamHI雙酶切及測序鑒定正確后,用脂質體法轉染293FT細胞,采用倒置熒光顯微鏡觀察GFP的表達、蛋白印跡(Western blot)法檢測PAX6蛋白表達。結果: 重組質粒pEF1α-PAX6-IRES-AcGFP經RT-PCR和雙酶切均得到大小為1 269 bp的目的條帶,測序鑒定該序列與GenBank中大鼠PAX6基因序列的同源性達100%,插入基因的大小和方向正確;重組質粒轉染293FT細胞后可見GFP綠色熒光,Western blot顯示PAX6蛋白表達。結論: 成功構建了PAX6真核表達重組質粒pEF1α-PAX6-IRES-AcGFP,能在293FT細胞中表達PAX6蛋白。
[關鍵詞]綠色熒光蛋白; PAX6基因; 表達載體,真核; 脂質體; 轉染
角膜盲是最重要的致盲眼病之一,全球約有超過1 000萬的角膜盲患者,我國現有角膜盲患者約300多萬,每年新發的感染性角膜病致盲患者約10萬人[1]。然而,角膜供體的極度匱乏,是造成角膜盲得不到及時治療的主要原因之一。位于角鞏膜交界處的角膜緣干細胞(limbal stem cells, LSCs)是角膜上皮細胞自我更新、修復及再生的基礎,能夠修復并替代衰老死亡的角膜上皮細胞,在角膜上皮的更新和角膜疾病的治療中起著不可替代的作用[1-2]。有學者發現人類與嚙齒類動物的PAX6基因整個編碼區域相似性為100%, WNT7A和PAX6是LSCs分化的關鍵因子,p63和PAX6協同作用可促進角膜緣干細胞的定向分化[3]。為LSCs移植應用于治療角膜盲等角膜疾病提供了研究方向。本課題通過構建真核表達載體pEF1α- PAX6-IRES-AcGFP的方法,觀察PAX6在人胚腎細胞293FT中的表達,為PAX6在誘導鼠胚胎干細胞分化的作用機制奠定實驗基礎。
1材料與方法
1.1實驗動物、試劑與儀器
體質量100~120 g的健康清潔級SD大鼠(貴州醫科大學實驗動物中心提供)。人胚腎細胞293FT細胞(貴州醫科大學干細胞中心細胞庫)、真核表達載體pEF1α-IRES-AcGFP(美國康涅狄格大學Lai Laijun教授惠贈), pUCm-T Vector(上海生工生物工程有限公司);Trizol?Reagent、脂質體Lipofectamine?2000(Invitrogen公司,美國);限制性內切酶SalI 和BamHI(New England Biolabs公司,美國),割膠回收試劑盒、質粒小抽試劑盒(天根生化科技有限公司,中國北京),T4 DNA Ligase(Takala,日本),氨芐霉素、卡那霉素、SDS-PAGE試劑盒、BCA法蛋白定量試劑盒(北京索萊寶生物科技有限公司,中國北京),高效感受態細胞制備試劑盒、內參β-actin(上海生工生物工程有限公司,中國上海),DMEM培養液、0.25%胰蛋白酶(Hyclone,美國),胎牛血清(杭州四季青生物制品有限公司,中國杭州),兔多克隆PAX6抗體、驢抗兔二抗(abcam,美國);其他試劑為國產分析純。CO2培養箱(Thermo Fisher,美國),高速冷凍離心機、全自動數碼凝膠成像分析系統、電泳儀(Eppendorf,德國),PCR儀(ABI,美國);熒光倒置相差顯微鏡(Nikon,日本)。
1.2方法
1.2.1PAX6基因的擴增和驗證根據GenBank提供的已知序列(GeneID:18508),對PAX6基因全CDS區序列設計引物:上游引物5′-ATCAGAGTCGACATGCAGAACAGTCACAGCGGAG-3′,下游引物5′-ATCTGAGGATCCTTACTGTAATCGAGGCCAGTACTG-3′,引物由上海生工生物工程有限公司合成。取健康清潔級SD大鼠5只,無菌條件下由枕骨或第1頸椎處入路,用止血鉗將顱頂向上撬開,暴露腦組織,將腦組織冰上勻漿,參照Trizol Total RNA說明書,采用一步法提取SD大鼠腦組織中的總RNA后再逆轉錄為cDNA,以cDNA為模板,進行PAX6基因PCR擴增。擴增體系為:2×Master Mix 10 μL,上游引物1 μL,下游引物1 μL,cDNA 2 μL,補雙蒸餾水(ddH2O)至20 μL。循環條件為: 95 ℃預變性5 min,95℃變性30 s、64 ℃退火30 s、72 ℃延伸1 min(35個循環),最后72 ℃延伸7 min。反應結束后取PCR產物,采用0.7%瓊脂凝膠電泳進行鑒定,并割膠回收目的片段。參照T載體試劑盒說明,將回收純化的PCR產物連接于pUCm-T Vector中,取5 μL連接反應液轉化DH5α感受態細菌,將轉化的菌液涂在含100 mg/L氨芐青霉素的LB平板上培養16 h。待LB平板長出菌落,挑取單菌落接種到5 mL LB的液體培養基中,37 ℃振蕩培養約12 h至對數生長后期,用質粒小量回收試劑盒抽提質粒DNA,經菌液PCR鑒定和限制性內切酶SalI 和BamHI進行雙酶切鑒定正確后,將正確的細菌陽性克隆送上海生工生物工程有限公司測序并驗證。
1.2.2真核表達質粒pEF1α-PAX6-IRES-AcGFP的構建將連有PAX6基因并測序正確的pUCm-PAX6-T Vector和真核質粒pEF1α-IRES-AcGFP分別用限制性內切酶SalI和BamHI對其進行雙酶切,回收1 269 bp的純化基因片段和酶切后線性化pEF1α-IRES-AcGFP質粒DNA,用T4 DNA連接酶將目的基因片段PAX6定向插入帶綠色熒光蛋白(GFP)報告基因的哺乳動物真核表達載體pEF1α-IRES-AcGFP中。將5 μL連接產物轉化至自制大腸桿菌DH5α感受態細胞,冰上放置30 min,42 ℃熱沖擊90 s,冰上放置5 min,加入200 μL無抗性LB液體培養基,37 ℃,250 r/min振蕩培養45 min,取菌液200 μL涂布于含50 mg/L的卡那霉素的LB固體培養基中,37 ℃倒置培養過夜。菌液PCR篩選陽性克隆,限制性內切酶SalI /BamHI雙酶切鑒定,將初步篩選的陽性克隆送上海生工生物工程有限公司測序。
1.2.3293FT細胞培養和重組質粒pEF1α-PAX6-IRES-AcGFP轉染293FT細胞于37 ℃,5% CO2條件下,用含10%胎牛血清的高糖DMEM培養基培養,細胞長滿傳代,轉染前24 h,以5×105個/孔的細胞密度接種于6孔板;待細胞融合>95%時,按Lipofectamine?2000說明書要求進行轉染,設立未轉染組和轉染空質粒pEF1α-IRES-AcGFP的293FT細胞做陰性對照。轉染4 h后更換普通培養基,常規培養24 h后于倒置熒光顯微鏡觀察綠色熒光蛋白(GFP)表達(綠色熒光)并拍照。以上實驗重復3次。
1.2.4PAX6蛋白表達采用Western blot 法,收集轉染后24 h的各組293FT細胞,提取細胞蛋白并用BCA蛋白定量試劑盒進行定量;取蛋白20 μg,經SDS-PAGE凝膠電泳后,采用轉移電泳裝置,于500 mA恒流條件下電轉60 min,將蛋白轉移到PVDF膜上;用封閉液(含5%脫脂牛奶的TBST溶液)封閉PVDF膜4 ℃過夜,TBST洗膜3次, 10 min/次;用封閉液稀釋的兔多克隆抗體PAX6(1∶1 000)與封閉好的PVDF膜室溫孵育2 h;TBST洗膜3次,10 min/次。室溫下用封閉液稀釋的二抗抗體(1∶5 000)與封閉好的PVDF膜室溫孵育2 h;TBST洗膜3次,10 min/次,用ECL試劑盒曝光獲得顯示條帶的膠片,掃描圖像保存,用β-actin(1∶8 000)作內參照驗證蛋白含量。以上實驗重復3次。
2結果
2.1大鼠PAX6基因cDNA的克隆
大鼠PAX6基因擴增產物經瓊脂糖凝膠電泳在1 269 bp處可見特異性條帶,結果見圖1。目的片段克隆入pUCm-T Vector和測序結果經NCBI/BLAST進行序列比對,符合率為100%(見圖2)。
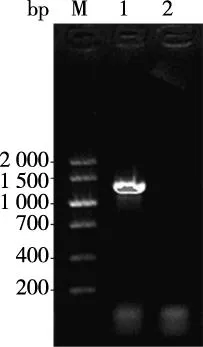
注:M為DNA Marker V,泳道1為PAX6基因片段,泳道2為陰性對照圖1 大鼠PAX6基因擴增產物Fig.1 Agrose gel electrophoresis detection of PCR products of PAX6 gene
2.2重組質粒pEF1α-PAX6-IRES-AcGFP的鑒定
重組質粒pEF1α-PAX6-IRES-AcGFP克隆后的菌液PCR產物經瓊脂糖凝膠電泳成像結果顯示在1 269 bp處見目的條帶(圖3);提取純化質粒,用限制性內切酶SalI 和BamHI對其進行雙酶切鑒定,瓊脂糖凝膠電泳成像結果顯示在1 269 bp和6 000 bp處見目的條帶和載體條帶(圖4)。
2.3重組質粒pEF1α- PAX6-IRES-AcGFP轉染293FT細胞
將轉染24 h后的293FT細胞置于熒光顯微鏡下觀察,細胞呈上皮樣成片生長,貼壁疏松,轉染前后細胞形態學無明顯變化,轉染空質粒pEF1α-IRES-AcGFP和重組質粒pEF1α-PAX6-IRES-AcGFP組的293FT細胞胞質內可見明顯的特異性綠色熒光,細胞未轉染組未見綠色熒光表達(圖5)。Western blot蛋白結果顯示,未轉染組和轉染空質粒組均未見PAX6表達,轉染重組質粒組在50.6 kDa處有特異性條帶(圖6)。
3討論
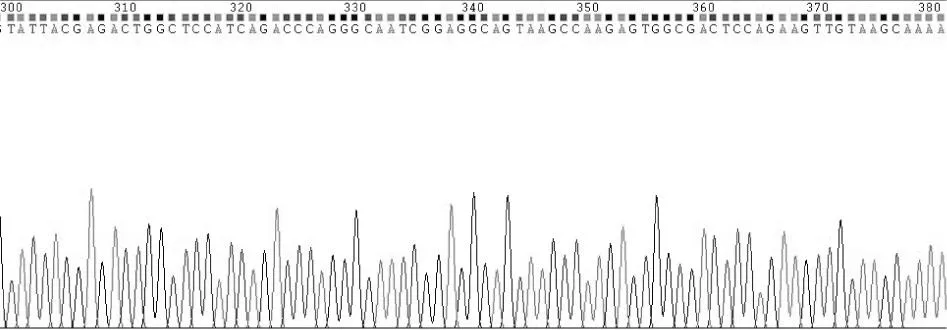
圖2 質粒pEF1α-PAX6-IRES-AcGFP部分測序結果Fig.2 A part of sequencing map of pEF1α-PAX6-IRES-AcGFP plasmid
外源蛋白在哺乳動物細胞中的表達量一般是由載體在染色體上的整合位置、啟動子的強度、轉錄產物的穩定性、mRNA的翻譯效率、目的蛋白的折疊效率以及目的蛋白的穩定性等因素決定,其中前4個因素均與表達載體的選擇有關[13]。所以表達載體的選擇是決定外源蛋白在哺乳動物細胞中表達量的關鍵,而啟動子的強度對于基因的表達起著決定性的作用。常用的啟動子有巨細胞病毒即早期啟動子 (PCMV- IE) 、 人延伸因子1α亞基啟動子 (PEF-1α)和 Rous肉瘤長末端重復序列等。本實驗選用含有人延伸因子1α亞基啟動子(PEF1α)的哺乳動物表達載體pEF1α- IRES-AcGFP用于目的基因PAX6的表達。啟動子PEF1α具有不受細胞周期影響,啟動子強度高等優點,可使PEF1α啟動子下游的蛋白實現高表達,也適用于難轉的細胞,有利于轉染和外源蛋白的大量擴增[12]。且該載體的多克隆位點中有一個IRES元件,可在PEF1α啟動子驅動下轉錄為雙順反子,實現目的基因與綠色熒光報告基因(AcGFP)獨立翻譯[13]。此外,該載體能夠在流式細胞儀中高效穩定檢測或瞬時轉染哺乳動物細胞表達目的基因與AcGFP,且可以用G418篩選穩定轉染細胞株,有利于后續轉染胚胎干細胞。
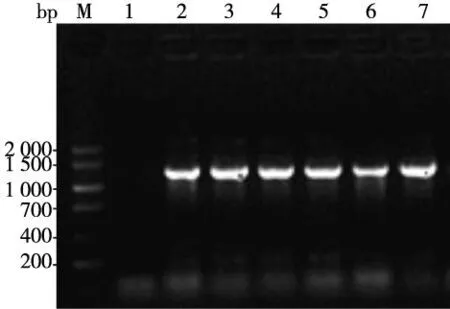
注:M為DNA Marker V,泳道1為陰性對照,泳道2~7為PAX6基因片段圖3 菌液PCR鑒定重組質粒pEF1α-PAX6-IRES-AcGFP Fig.3 The electrophoresis of bacterial colony PCR product of plasmid pEF1α-PAX6-IRES-AcGFP
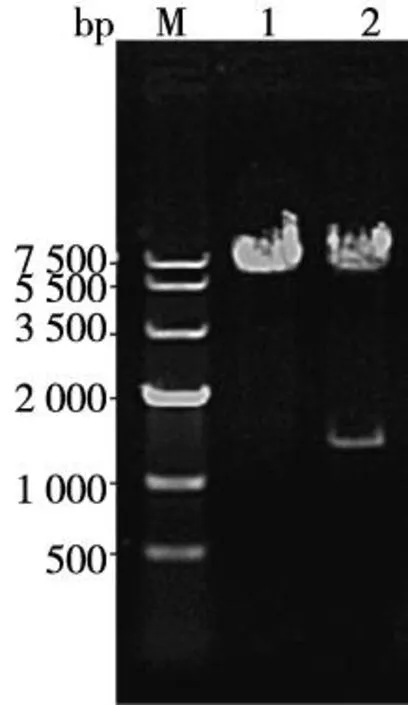
注:泳道M為DNA marker IV,泳道1為pEF1α-IRES-AcGFP雙酶切結果,泳道2為重組質粒pEF1α-PAX6-IRES-AcGFP雙酶切圖4 SalI/BamHI雙酶切鑒定真核質粒pEF1α-PAX6-IRES-AcGFPFig.4 The electrophoresis of plasmid pEF1α-IRES-PAX6-AcGFP digested product by SalI and BamHI
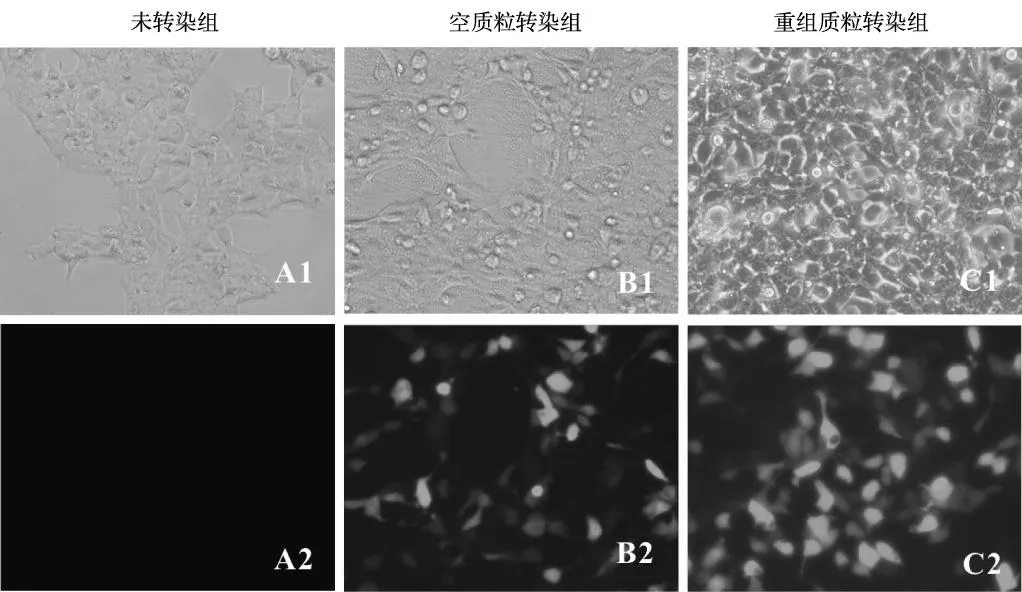
注:A1、B1和C1為顯微鏡(×200)明場下的293FT細胞;A2 、B2和C2為顯微鏡(×200)488 nm波長激發光下的293FT細胞圖5 真核質粒pEF1α-PAX6-IRES-AcGFP轉染293FT細胞24 h后鏡下觀察(200×)Fig.5 Transfected ppEF1α-PAX6-IRES-AcGFP into 293FT cells
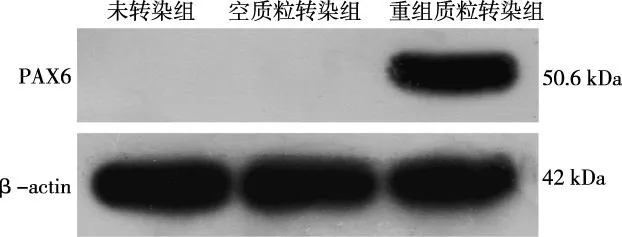
圖6 轉染后293FT細胞中PAX6蛋白表達Fig.6 The protein expression after PAX6 transfection in 293FT cell by western blotting analysis
在克隆目的基因PAX6時,本研究先將PAX6連接于T載體,測序正確后,再經酶切定向插入到真核載體。T載體是TA克隆載體的簡稱,是一種能夠直接克隆PCR產物的工具,本研究采用的pUCm-T載體是一種已經線性化的載體,載體每條鏈的3’端帶有一個突出的T(脫氧胸苷),載體的兩端可以和PCR產物的兩端進行正確的AT配對,在連接酶的催化下,可以把PCR產物連接到pUCm-T載體中,形成含有目的片斷的重組載體,能夠滿足基因保存和測序的需要[14-15],且能克服常規方法中載體自連、過程復雜、克隆效率低等不足,提高了連接效率和陽性率。293FT細胞是一種改裝過的人胚腎細胞,由293細胞派生,FT的含義就是帶有能復制(F)的SV40大T抗原,含有SV40復制起始點與啟動子區的質粒可以復制,比較容易轉染,被廣泛應用于瞬時轉染以過表達各種目標蛋白。脂質體轉染法是利用脂質體脂質雙分子層的融合及內吞作用使外源基因PAX6進入293FT細胞。
PAX6基因是PAX基因家族成員,是控制著眼球形態發生的一個重要調節基因[4-6],在眼、神經系統、鼻、胰腺和內分泌等組織器官的發育中起著重要作用,廣泛參與細胞增殖遷移分化和黏附等生命活動[7-8]。在眼睛發育過程中,PAX6從胚胎第8天表達,時間早于任何形態學分化[9],表面外胚層中, PAX6表達于眼睛發育最原始的階段,基板形成前PAX6就已經表達,一直持續表達到晶狀體分化,最終存在于成人晶狀體和角膜上皮,是眼發育的主控基因[3, 11]。Ouyang[3]的研究發現,將PAX6轉到皮膚上皮干細胞(Skin epithelial stem cells, SECs)中時,發現其可以充分地將SECs轉變為表達K5/K14的樣細胞,再將此細胞移植應用于角膜上皮缺損的兔子模型中發現,移植的細胞最初位于角膜緣的地區,然后逐步向中央角膜與相應的區域移動,3個月后,成功修復缺損的角膜并維持正常的角膜透明度。而將敲除PAX6基因的兔LSCs移植到此模型并不能修復角膜損傷,提示PAX6在修復角膜損傷中起到決定性的作用。由于LSCs獲得的局限性,本課題構建攜帶眼發育關鍵基因PAX6的真核表達載體,初步探究其在真核細胞的表達,再進一步探討PAX6轉染胚胎干細胞并進一步誘導胚胎干細胞定向分化為LSCs,以期建立穩定細胞株,為角膜損傷提供種子細胞。
本課題成功構建重組真核表達載體pEF1α-PAX6-IRES-AcGFP,并通過脂質體轉染法將重組載體轉入293FT細胞,轉染24 h后熒光顯微鏡下觀察到綠色熒光,Western blot結果顯示轉染重組質粒組PAX6蛋白高表達,未轉染組和轉染空質粒組沒有PAX6蛋白表達,證明過表達質粒成功轉染了293FT細胞并建立了PAX6過表達的293FT細胞瞬時轉染模型,為后續研究中PAX6在真核細胞中的調節作用及PAX6轉染胚胎干細胞誘導分化為角膜緣干細胞實驗奠定了基礎。
4參考文獻
[1] Ordonez P, Girolamo ND. Limbal epithelial stem cells: role of the niche microenvironment[J]. Stem cells, 2012(2):100-107.
[2] Yoon JJ, Ismail S, Sherwin T. Limbal stem cells: Central concepts of corneal epithelial homeostasis[J]. World journal of stem cells, 2014(4):391.
[3] Ouyang H, Yuanchao X, Ying L, et al. WNT7A and PAX6 define corneal epithelium homeostasis and pathogenesis[J]. Nature, 2014(2):358-361.
[4] Shaham O, Menuchin Y, Farhy C, et al. Pax6: A multi-level regulator of ocular development[J]. Progress in Retinal and Eye Research, 2012(5):351-376.
[5] Ashery-Padan R ,Gruss P. Pax6 lights-up the way for eye development[J]. Current Opinion in Cell Biology, 2001(6):706-714.
[6] Xu S, Sunferland ME, Coles BLK, et al. The proliferation and expansion of retinal stem cells require functional Pax6[J]. Developmental Biology, 2007(2):713-721.
[7] Noll M. Evolution and role of Pax genes[J]. Current Opinion in Genetics & Development, 1993(4):595-605.
[9] Yao JG, Weasner BM, Wang LH, et al. Differential requirements for the Pax6(5a) genes eyegone and twin of eyegone during eye development in Drosophila[J]. Developmental Biology, 2008(2):535-551.
[10]Gehring WJ, Ikeo K. Pax6: mastering eye morphogenesis and eye evolution[J]. Trends in Genetics, 1999(9):371-377.
[11]Kozmik Z. The role of Pax genes in eye evolution[J]. Brain Research Bulletin, 2008(2-4):335-339.
[12]Novosylna O, Jurewicz E, Pydiura N, et al. Translation elongation factor eEF1A1 is a novel partner of a multifunctional protein Sgt1[J]. Biochimie, 2015(119):137-145.
[13]Abdul Mutalib NE, Matlsa N, Alitheen NB, et al. IRES-incorporated lactococcal bicistronic vector for target gene expression in a eukaryotic system[J]. Plasmid, 2014(73):26-33.
[14]Peng RH, Xiong AS, Liu J, et al. Adenosine added on the primer 5′ end improved TA cloning efficiency of polymerase chain reaction products[J]. Analytical Biochemistry, 2007(1):163-165.
[15]Adachi Y, Fukuhara C.TA strategy for rapid and efficient site-directed mutagenesis[J]. Analytical Biochemistry, 2012(1):66-68.
(2015-12-05收稿,2016-02-22修回)
中文編輯: 吳昌學; 英文編輯: 劉華
Construction and Authenticate of Eukaryotic Expression Vector with Rat PAX6
YAN Huiwen, SU Min, GAO Jie, LIAO Wenping, YAN Lili, HUANG Yue
(DepartmentofHistologyandEmbryology,GuizhouMedicalUniversity,Guiyang,Guizhou, 550004)
[Abstract]Objective: To construct a green fluorescence protein report gene (GFP) and an eukaryotic expression vector of rat PAX6 gene and, observe its expression in 293FT cells. Methods: The rat PAX6 gene was amplified by Polymerase Chain Reaction (PCR). After verified correctness of sequencing by connection with T vector, the recombinant products of the pEF1α-PAX6-IRES-AcGFP were gained by T4 ligase connecting pUCm-T-PAX6 and eukaryotic expression vector pEF1α-IRES-AcGFP with restriction enzymes SalⅠ/BamH. After identified by colony PCR and sequencing, pEF1α-PAX6-IRES-AcGFP was transfected into 293FT cells with Lipofectamine(?)2000 assay. The expression of PAX6 protein and GFP were detected by western blot and fluorescence microscope. Results: A specific band of 1269bp was detected from recombinant plasmid pEF1α-PAX6-IRES-AcGFP by digestion of SalⅠand BamHⅠand RT-PCR. Sequencing and identification showed that homology between this sequence and the rat PAX6 gene sequence from GenBank was 100%, and the size and the direction of the inserted gene were right. After transfection, 293FT cells showed green fluorescence under fluorescence microscope. A band of PAX6 protein from cells was detected by western blot assay. Conclusion: The eukaryotic expression vector of PAX6 is constructed successfully and can obtain high PAX6 protein expression in 293FT cells.
[Key words]green fluorescence protein; PAX6 gene; expression vector,eukaryotic; lipofectamine; transfection
[中圖分類號]R329.2
[文獻標識碼]A
[文章編號]1000-2707(2016)03-0288-06
*[基金項目]貴州省科技廳基金[黔科合J字(2015)2011號]; 貴州省優秀科技教育人才省長專項資金項目[黔省專合字(2012)41號]
**貴州醫科大學2013級碩士研究生
***通信作者 E-mail:huranggaigaiyu@sina.com
網絡出版時間:2016-03-17網絡出版地址:http://www.cnki.net/kcms/detail/52.5012.R.20160317.1106.058.html
