核桃炭疽病病原菌鑒定及生物學特性
2016-03-31 07:07:54王清海劉幸紅段春華牛贍光吳小芹1南京林業大學南方現代林業協同創新中心南京林業大學林學院江蘇南京1007山東省林業科學研究院山東濟南50014山東省果樹研究所山東泰安71018
山東農業大學學報(自然科學版) 2016年1期
關鍵詞:生物學
王清海,劉幸紅,范 昆,段春華,牛贍光,吳小芹1.南京林業大學南方現代林業協同創新中心,南京林業大學林學院,江蘇南京1007.山東省林業科學研究院,山東濟南50014.山東省果樹研究所,山東泰安71018
?
核桃炭疽病病原菌鑒定及生物學特性
王清海1,2,劉幸紅2,范昆3,段春華2,牛贍光2,吳小芹1*
1.南京林業大學南方現代林業協同創新中心,南京林業大學林學院,江蘇南京210037
2.山東省林業科學研究院,山東濟南250014
3.山東省果樹研究所,山東泰安271018
摘要:本文從表現炭疽病癥狀的核桃果實上分離到一株具有強致病性菌株(TS-09),并進行了鑒定和生物學特性研究。經形態特征觀察、培養性狀以及rDNA-ITS、ACT、β-tub2、GPDH四種保守基因序列分析,TS-09菌株為膠孢炭疽菌(Colletotrichum gloeosporioides)。生物學特性測定結果表明:TS-09菌株在PDA培養基上生長最好;25~30℃之間適宜菌株生長及產孢,28℃是菌株最適宜的生長及產孢溫度;弱酸性環境(pH4.0)有利于產孢,中性環境(pH7.0)適宜菌絲生長;長時間光照抑制菌絲生長,黑暗條件有利于菌絲生長;菌絲的抗逆性較強,60℃處理15 min、62℃處理10 min,菌絲死亡。研究結果將為核桃炭疽病的預測及有效控制提供理論依據。
關鍵詞:核桃炭疽病;鑒定;生物學
核桃(Juglans regia)是一種世界性分布的重要“木本糧油”生態樹種。核桃仁含有多種適宜于人體健康的營養成分,被譽為“21世紀的超級食品”。核桃木是可以用于制作高級家具和軍事用材。核桃種植具有良好的經濟價值和社會效益。近年來,我國核桃種植業發展迅速。據FAO統計,2013年中國核桃樹收果面積4.25×105hm2,產量4.0×104Hg/hm2,總產量1.70×106t,山東省核桃種植面積已達9.13×104hm2,產量9.3×104t。核桃種植面積的迅猛增加,集約化程度的不斷提高,但栽植品種以早食核桃為主,抗病性較差,加之密植的栽培模式致使核桃病害發生日趨嚴重。炭疽病可危害果實、葉片,潛伏期長、發病時間短、爆發性強,嚴重時病果率高達80%以上,落果率達50%以上,嚴重影響核桃產量和品質,造成嚴重的經濟損失,是核桃生產中的一類災難性病害[1,2]。引起炭疽病的炭疽菌屬(Colletotrichum Corda.)真菌是全球公認的第八類重要的植物病原菌[3],對全球農林業生產構成了嚴重威脅。其中多個種如Colletotrichum acutatum可造成草莓、藍莓、芒果、枇杷、橄欖、獼猴桃、鱷梨等高檔水果品質降低,貨架期縮短[4-6],Colletotrichum kahawae侵染咖啡,引起非洲咖啡毀滅性的災害[7,8],Colletotrichumgloeosporioides引起柿、棗、蘋果、葡萄、梨、石榴等鮮食水果炭疽病[9,10],Colletotrichumgraminicola嚴重影響玉米、甘蔗以及高粱等谷物產量[11]。隨著炭疽菌研究的深入,越來越多的研究結果表明同種寄主植物上會存在多種炭疽菌。C. acutatum和C. gloeosporioides可以引起橄欖炭疽病[6]。劉威等人研究發現C. gloeosporioides和Colletotrichumfructicola均可以引起茶樹炭疽病[12]。2012年山東省核桃炭疽病大爆發,是否存在多種炭疽菌引起核桃炭疽病,尚未明確。筆者從山東省核桃產區表現炭疽病癥狀的果實上分離獲得的一株強致病菌。由于該屬真菌形態特征、生物學特性以及遺傳變異較大,本文采用形態學、分子生物學方法對該菌株進行了種類鑒定,并對生物學特性進行了研究,旨在為該病害的防治提供科學依據。
1 材料與方法
1.1供試材料
核桃感病果實,從山東濟南、泰安、臨沂、棗莊等核桃主產區核桃園中采集。
1.2核桃病果菌株分離培養
將采集的病果,帶回實驗室,先用70%酒精對病果表面消毒,然后置于無菌的燒杯中,底部鋪3層滅菌濾紙,加入少量無菌水,保持濕度,用保鮮膜密閉燒杯,置于28℃培養箱中,培養1~2 d,誘導病果上病菌產孢。挑取果實表面粉紅色分生孢子堆,在PDA平板劃線,培養。挑取單個菌落,進行純化。
1.3致病性測定
在實驗室進行離體果實接種試驗,品種為香玲。選擇無病無蟲傷的核桃果實,自來水沖洗,晾干后,用75%酒精表面消毒,采用注射接種法,將分離的菌株用無菌水配制成105個·mL-1的孢子懸浮液,每個果實在3個不同位置接種,每個接種點接種20 μL孢子懸浮液,以無菌水為對照,每個處理5個果實,每個處理平行測定4次,試驗重復3次。處理后將核桃置于滅菌的燒杯中,用保鮮膜密閉燒杯,保持一定濕度,置于28℃的恒溫培養箱中,待發病后,取發病核桃,按1.2進行分離。
1.4病原菌鑒定
1.4.1形態特征觀察將TS-09菌株接種于PDA平板,28℃培養5 d,觀察菌落特征,顯微觀察分生孢子形態、大小以及附著胞形態。
1.4.2TS-09菌株部分保守基因序列分析及系統發育樹構建DNA的提取采用改進的CTAB法,試驗所用引物見表1[13-16],反應條件:25 μL反應體系,包括Taq酶mix 12.5 μL,上游引物下游引物各1 μL,ddH2O 8.5 μL,DNA模板2 μL。具體反應程序如下:95℃預變性5 min,94℃變性0.5 min,59℃退火0.5 min(ITS退火溫度58℃,GPDH退火溫度56℃),72℃延伸1.5 min,共36個循環;最后72℃延伸7 min,4℃保存。
PCR產物由上海生物工程技術服務有限公司進行雙向測序。利用NCBI的BLAST程序,將測序結果與GenBank(http://www.ncbi.nlm.nih.gov)數據庫中相關序列進行同源性比較分析。利用DNAMAN軟件對各基因序列進行比對,通過手工校正使序列排序獲得優化,將校正后各基因按首尾相連的方法合并,利用MEGA5.05軟件鄰近(Neighbor-Joining,NJ)法構建多基因系統發育樹,以自展法(Bootstrap)進行檢測,共循環1000次。

表1 引物序列Table1 Sequence of the primers
1.5核桃炭疽菌生物學特性測定
1.5.1不同培養基對菌絲生長和產孢影響測定取生長一致的核桃炭疽菌菌餅(Φ9 mm),分別接種于PDA培養基、PSA培養基、PMA培養基、燕麥片培養基(OA)、玉米粉瓊脂培養基(CMA)、馬丁氏培養基(Martin)、沙氏培養基(SDA)等培養基平板上,28℃恒溫培養5 d,采用十字交叉法測量菌落直徑,觀察菌落生長狀況,并于第10 d測產孢量[17,18]。每個處理平行測定4次,3次重復。
1.5.2溫度對菌絲生長和產孢影響測定將菌餅(Φ9 mm)置于PDA平板上,分別置于5℃、15℃、20℃、25℃、28℃、30℃、35℃的恒溫培養箱中培養,每日17時測量菌落直徑,第10 d測產孢量,處理、重復、測定方法同1.5.1。
1.5.3pH值對菌絲生長和產孢影響測定分別配置pH為2、3、4、5、6、7、8、9、10的PDA平板。配置低pH值的培養基時先配好PDA液體培養基,并調整到相應的pH值,然后用其他的三腳瓶盛放相應量瓊脂粉。同時滅菌,溫度降低到59℃時把液體瓊脂快速倒入PDA液體培養基中,制作平板。將菌餅(Φ9 mm)置于不同pH值的培養基上,28℃下培養5 d。處理、重復、測定方法同1.5.1。
1.5.4光照條件對菌絲生長影響測定將菌餅(Φ9 mm)置于PDA平板上,設12 h日光燈交替照射、日光燈24 h持續照射、連續黑暗3個處理,于28℃下培養,處理、重復、測定方法同1.5.1。
1.5.5菌株致死溫度測定將菌餅置于滅菌試管中,每試管放10枚菌片,分別在45℃、50℃、55℃、60℃、65℃的恒溫水浴鍋中分別處理10 min、15 min。隨機從每個試管中取出5枚菌餅置于PDA平板,28℃恒溫培養2 d后,以菌絲能夠生長的最高溫度為基礎,增加1℃設為1個處理,處理中又設10、15 min 2個時間,最后確定致死溫度及相應時間,每個處理平行測定4次,重復3次。
1.6數據處理與分析
數據處理采用Spss20.0軟件,采用Duncan多重比較法進行差異顯著性分析。
2 結果與分析
2.1核桃病果菌株分離及致病性
誘導病果發病后,均分離到真菌菌株,單孢純化后獲得一株優勢菌株(編號為TS-09)。將TS-09菌株孢子懸浮液接種于健康的核桃果實,發病率為100%,對照未發病。病斑初期褐色,逐漸變黑色,近圓形至不規則形,中央稍凹陷,病部有黑色小點產生,潮濕條件下,黑點上溢出粉紅色粘質孢子堆。癥狀與田間發病一致,且從這些病斑上都能夠再分離到與原接種菌株相同的菌株。這表明分離到的TS-09菌株為該病的致病菌。
2.2核桃炭疽病菌TS-09菌株鑒定
TS-09菌株在PDA培養基平板上菌落棉絮狀,邊緣完整,初期灰白色,隨后在平板中央變為深灰色,后期有粉紅色分生孢子堆出現。分生孢子盤褐色至黑褐色;分生孢子單胞、無色、長橢圓形,大小為10.15±0.21×3.37±0.09 μm,附著胞圓形,淡褐色(圖1)。利用ACT、rDNA-ITS、GPDH、β-tub2 等4種保守基因構建的多系統發育樹表明,TS-09菌株與不同來源地膠孢炭疽菌模式菌株聚在一起,與其它炭疽菌形成明顯分支。結合TS-09菌株的形態特征、培養性狀,鑒定TS-09菌株為膠孢炭疽菌C. gloeosporioides。
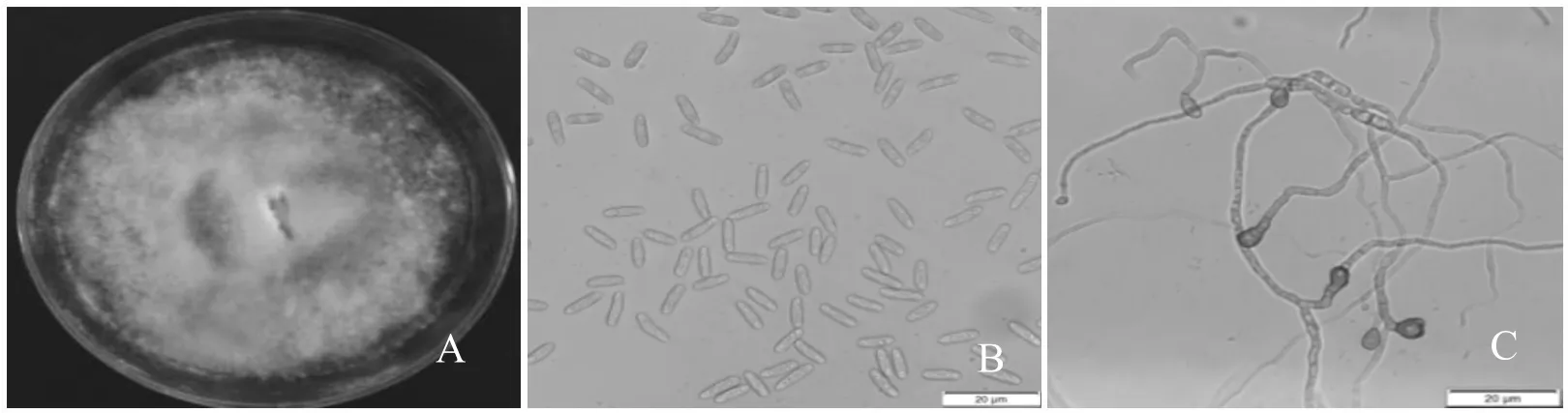
圖1 TS-09菌株形態特征A.菌落培養性狀;B.分生孢子;C.附著胞Fig.1 The culture characters of strain TS-09A. Colony characteristics on PDAfor15d;B.Conidia;C. Appressoria
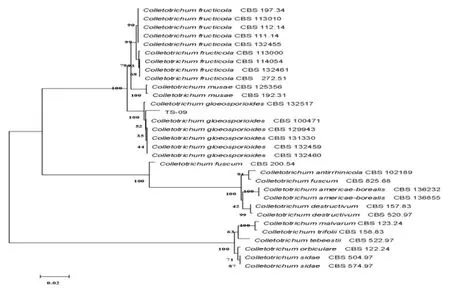
圖2 基于ACT, rDNA-ITS, GPDH和β-tub2序列的TS-09菌株系統發育樹Fig.2 Phylogenetic tree of strain TS-09 based on ACT, ITS, GPDH and β-tub2 genes sequences data
2.3不同培養基對核桃炭疽菌培養性狀及菌絲生長的影響
TS-09菌株在不同培養基上培養性狀稍有差異(圖3)。在營養豐富的培養基上如PDA、PMA、PSA和SDA培養基上,菌絲稠密,灰白色至淺灰色,顏色均一,培養基背面淡黃色,顏色由里至外逐漸變淺。在營養相對缺乏的培養基如CMA培養基和OA培養基上菌絲稀疏,白色,棉絮狀,培養基背面與培養基顏色基本一致。在SDA、CMA、OA和Martin培養基上易產生分生孢子堆。
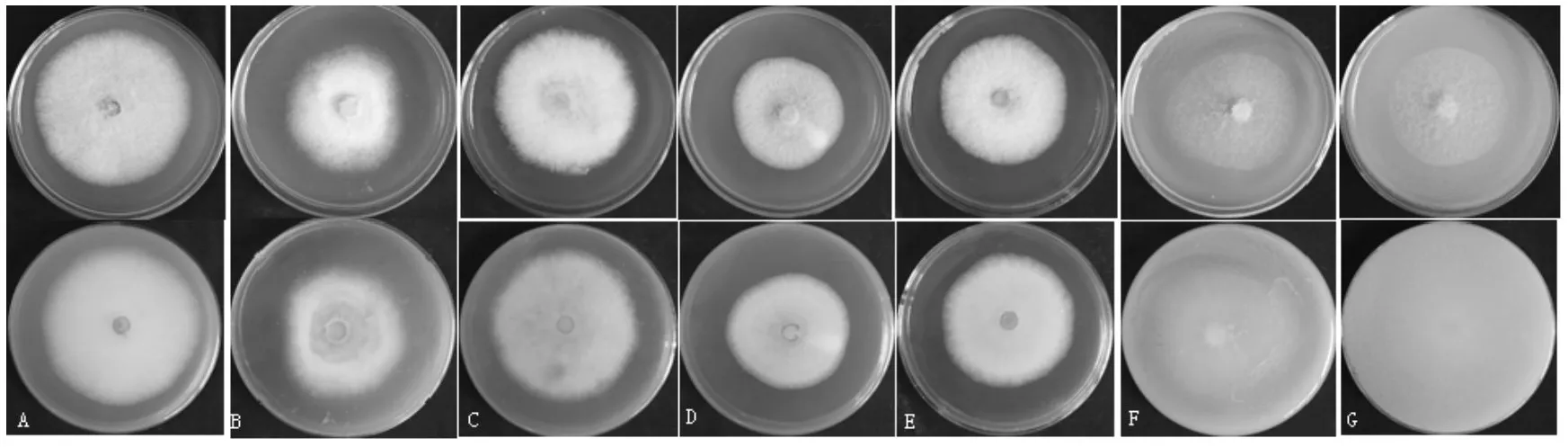
圖3 TS-09菌株在不同培養基上的形態特征Fig.3 The morphological characteristics of strain TS-09 on various culture media for 5 d growthA.PDA,B.PMA,C. PSA,D. Martin,E.SDA,F. OA,G.CMA

由圖4可以看出,TS-09菌株在PDA培養基上生長最好,菌落直徑達68.58±1.18 mm,與其他處理相比,在P=0.01水平上達到極顯著水平。其次為PSA培養基。在Martin培養基上生長最差,菌落直徑僅為46.67±0.74 mm。
2.4溫度對核桃炭疽菌TS-09菌株菌絲生長和產孢的影響
圖5結果表明,溫度對TS-09菌株菌絲生長和產孢的影響趨勢一致,低于5℃,TS-09菌株停止生長及產孢,高于35℃,菌絲生長緩慢,不能產孢。在25~30℃之間適宜菌株生長及產孢,28℃,菌絲生長及產孢達到最大,是菌株的最適宜的生長及產孢溫度。每年的7月下旬,雨量大,雨水勤,溫度高,適于核桃炭疽病的大面積發生,高于35℃,低于20℃,病害發生程度有所減輕。
2.5pH對核桃炭疽菌TS-09菌株菌絲生長和產孢的影響
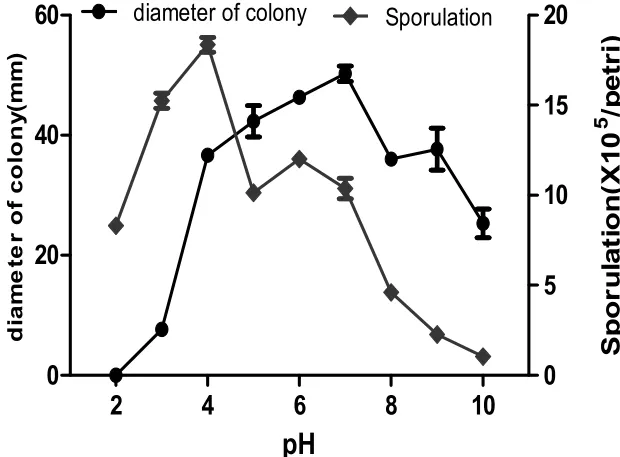
Fig.6 Effect of different pH on mycelial growth and sporulation of strain TS-09.
適宜TS-09菌株菌絲生長及產孢的pH值范圍較廣,在pH3.0~10的范圍內菌絲均能正常生長。pH7.0,菌絲生長最好。弱酸性環境易于產孢,pH4.0時產孢量最大(圖6)。
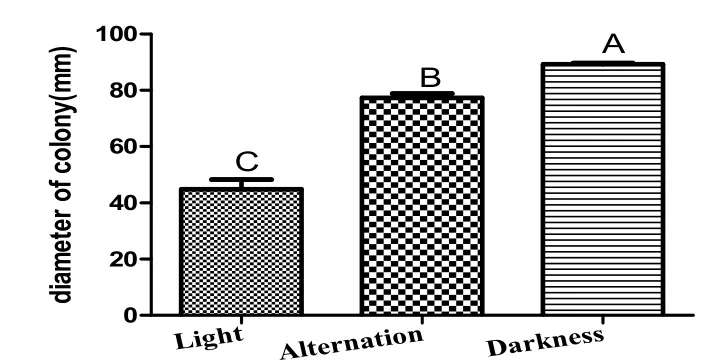
Fig.7 Effects of different light on colony diameter of strain TS-09.
2.6光照條件對TS-09菌株菌絲生長的影響
圖7結果表明,光照對TS-09菌株菌絲生長具有較強的影響。TS-09菌株適宜在黑暗條件下生長,與其它兩個處理在P=0.01水平上具有顯著性差異。與12 h日光燈交替照射相比,持續光照則抑制菌絲生長。
2.7核桃炭疽菌TS-09菌株菌絲致死溫度
TS-09菌株菌絲在65℃處理10 min、15 min均死亡,60℃處理10 min,菌絲能夠存活,處理15 min死亡,以60℃為起點,再次增加61℃、62℃、63℃、64℃等4個處理,繼續測定菌株的致死溫度。結果表明TS-09菌株菌絲的致死溫度為60℃處理15 min,62℃處理10 min(表2)。
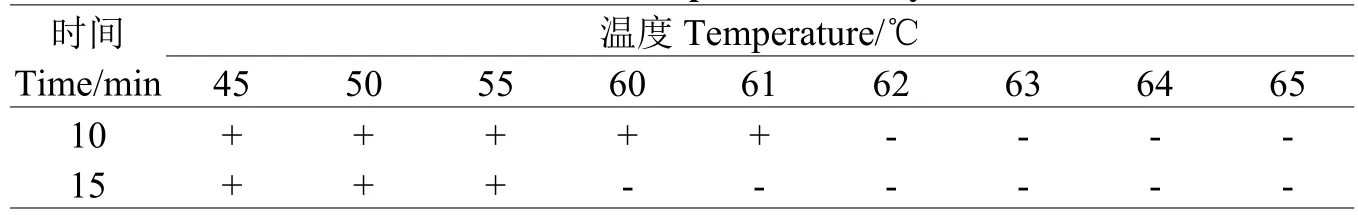
表2 菌絲的致死溫度Table 2 The lethal temperature of mycelium
3 結論與討論
炭疽菌屬真菌1790年首次發現,直到1831年Corda才首次建立炭疽菌屬。截止目前,炭疽菌屬真菌的分類、鑒定工作雖然取得了長足的進步,但其分類還是依然比較混亂,仍有一些爭議的問題有待進一步考證。炭疽菌屬真菌具有豐富的種群多樣性,形態特征、培養性狀、致病力以及遺傳變異較大,有些近似種之間差異微小,而且有時會有多種炭疽菌同時存在同一寄主的現象。所以只有依據形態學、致病性、系統發育學以及多基因的系統遺傳學才能夠提供較準確的結果。近年山東省核桃產區炭疽病發生危害程度日益嚴重。為了有效控制核桃炭疽病的暴發流行,明確病原種類是十分必要的。本實驗從山東省核桃產區核桃病果上的分離獲得了一株強致病力菌株TS-09,通過對菌株的ACT、rDNA-ITS、GPDH、β-tub24種保守基因的擴增序列分析,結合形態特征、培養性狀,將TS-09菌株鑒定為膠孢炭疽菌C. gloeosporioides。實驗結果表明膠孢炭疽菌是引起近幾年山東省核桃產區炭疽病暴發流行的主要致病菌。TS-09菌株在PDA培養基平板上菌落棉絮狀,初期灰白色,隨后在平板中央變為深灰色,后期有粉紅色分生孢子堆。附著胞圓形。分生孢子單胞、無色、長橢圓形,大小為10.15±0.21×3.37±0.09 μm,這與秦巴山區核桃炭疽菌的分生孢子形態、大小基本一致[19]。
一種炭疽菌可以侵染多種寄主植物,也可以多種炭疽菌侵染同一寄主植物。Embaby等人研究發現C. acutaum和C. gloeosporioides均可以引起草莓炭疽病[20];危害辣椒,引起辣椒炭疽病的有C. acutatum、C. truncatum、C. fructicola和C. siamense等4種炭疽菌[21-23]。本研究僅對從病果上分離到的一株強致病性菌株進行了鑒定,至于核桃上是否存在其它種的炭疽菌還會進行后續研究。
明確核桃炭疽菌的生物學特性是重要的研究基礎。本實驗對核桃炭疽菌TS-09菌株的生物學特性進行了研究。結果表明,核桃炭疽菌TS-09菌株在不同培養基上培養性狀略有不同。在營養相對豐富的培養基上,菌絲生長迅速;營養相對缺乏的培養基不利于菌絲生長,適于產生大量分生孢子。這表明菌株在逆性環境中,易通過產生分生孢子來度過不良環境。菌株生長及產孢的適宜溫度為28℃。TS-09菌株適宜在中性環境中生長,偏酸性環境中易于產孢,這與侵染草莓的膠孢炭疽菌生物學特性不同。侵染草莓的膠孢炭疽菌更適合在酸性條件生長,中性條件產孢[24]。光照對菌絲生長具有較強的影響,24 h光照,不適宜菌絲生長,黑暗條件利于菌絲生長,這可能也是核桃園內栽植密度過大時病害發生較重的原因之一。病原菌的生物學特性與病害田間的發生、消長規律密切相關,山東省每年的7月份雨季來臨,溫度高,濕度大,非常適宜核桃炭疽菌的生長、產孢和侵染,田間核桃炭疽病發生嚴重,實驗結果與田間發生規律相吻合。本研究結果明確了引起山東省核桃主產區炭疽病的強致病性病原種類及其生物學特性,將為預測病害發生和有效控制提供一定的理論依據。
田間觀察發現,核桃炭疽病菌每年4月份開始侵染,7月中下旬暴發,潛伏期長。該病菌的潛伏侵染與核桃葉片果實自身代謝產物的關系以及影響該病短時間暴發的作用因子還有待深入研究。
參考文獻
[1] An HS,Yang KQ. Resistance gene analogs in walnut(Juglans regia)conferring resistance to Colletotrichum gloeosporioides[J]. Euphytica,2014,197(2):175-90
[2]曲文文,楊克強,劉會香,等.山東省核桃主要病害及其綜合防治[J].植物保護,2011,37(2):136-140
[3] Dean R,Van Kan JAL,Pretorius ZA,et al. The Top 10 fungal pathogens in molecular plant pathology[J]. Molecular Plant Pathology,2012,13(4):414-4130
[4] Urdaneta L,Sanabria ME,Rodríguez D,et al. Antracnosis caused by Colletotrichum acutatum Simmonds in strawberry fruit in Lara and Trujillo states,Venezuela[J]. Revista de la Facultad de Agronomia,2013,30(4):504-28
[5] Miles TD,Gillett JM,Jarosz AM,et al. The effect of environmental factors on infection of blueberry fruit by Colletotrichum acutatum[J]. Plant Pathology,2013,62(6):1238-1247
[6] Talhinhas P,Gon?alves E,Sreenivasaprasad S,et al. Virulence diversity of anthracnose pathogens(Colletotrichum acutatum and C. gloeosporioides species complexes)on eight olive cultivars commonly grown in Portugal[J]. European Journal of Plant Pathology,2015,142(1):73-83
[7] Abera A,Lemessa F,Muleta D. The antifungal activity of some medicinal plants against coffee berry disease caused by Colletotrichum kahawae[J]. International Journal of Agricultural Research,2011,6(3):268-79
[8] Loureiro A,Guerra-Guimar?es L,Lidon FC,et al. Isoenzymatic characterization of Colletotrichum kahawae isolates with different levels of aggressiveness[J]. Tropical Plant Pathology,2011,36(5):287-93
[9] Rahimlou S,Babaeizad V,Sayari M. First report of fruit spot of pomegranate caused by Colletotrichum gloeosporioides in Iran[J]. Journal of Plant Pathology,2014,96(3):605
[10] Palou L,Montesinos-Herrero C,Tarazona I,et al. Postharvest anthracnose of persimmon fruit caused by Colletotrichum gloeosporioides first reported in Spain[J]. Plant Disease,2013,97(5):691
[11] Sukno SA,Sanz-Martín JM,González-Fuente M,et al. First report of anthracnose stalk rot of maize caused by Colletotrichum graminicola in Switzerland[J]. Plant Disease,2014,98(5):694
[12]劉威,葉乃興,陳玉森,等.茶樹炭疽菌Colletotrichum fructicola的鑒定及系統發育分析[J].茶葉科學,2014,34(1):95-104
[13] O'Donnell KA. Multigene phylogeny of the Gibberella fujikuroi species complex:Detection of additional phylogenetically distinct species[J]. Mycoscience,2000,41(1):61-78
[14] Glass NL,Donaldson GC. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes[J].Applied and Environmental Microbiology,1995,61(4):1323-30
[15] Carbone I,Kohn LM. A method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia,1999,91(3):553-556
[16] O'Donnell K,Cigelnik E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous[J]. Molecular Phylogenetics and Evolution,1997,7(1):103-16
[17] Kleemann J,Rincon-Rivera LJ,Takahara H,et al. Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum[J]. PLoS Pathogens,2012,8(4): e1002643
[18]范昆,張雪丹,余賢美,等.無花果炭疽菌的生物學特性及8種殺菌劑對其抑制作用[J].植物病理學報,2013,43(1):75-81
[19]肖玉貴,周建華,肖銀波.秦巴山核桃炭疽病病原菌生物學特性及防治技術的研究[J].四川林業科技,2010,31(1):54-57
[20] Embaby EM,Ragab ME,Dougdoug KA,et al. Identification of Colletotrichum acutatum and C. gloeosporioides on strawberry in Egypt[A].Acta Horticulturae,2012:657-60
[21] Diao Y,Zhang C,Xu J,et al. Genetic differentiation and recombination among geographic populations of the fungal pathogen Colletotrichum truncatum from chili peppers in China[J]. Evolutionary Applications,2015,8(1):108-118
[22] Sharma G,Shenoy BD. Colletotrichum fructicola and C. siamense are involved in chilli anthracnose in India[J]. Archives of Phytopathology and Plant Protection,2014,47(10):1179-1194
[23] Vitale S,Infantino A. Presence of Colletotrichum acutatum causing anthracnose on hot pepper in central Italy[J]. Journal of Plant Pathology,2014,96(3):607
[24]張海英,張明會,劉志恒,等.草莓炭疽病病原鑒定及其生物學特性研究[J].沈陽農業大學學報,2007,38(3):317-321
Identification and Biological Characteristics of Pathogen from Colletotrichum gloeosporioides
WANG Qing-hai1,2,LIU Xing-hong2,FAN Kun3,DUAN Chun-hua2,NIU Shan-guang2,WU Xiao-qin1*
1. Co-innovationCenterforSustainableForestryinSouthernChina,CollegeofForestry/NanjingForestryUniversity,Nanjing210037,China
2. Shandong Provincial Academy of Forestry, Ji’nan 250014,China
3. Institute of Pomology, Tai’an 271018,China
Abstract:Walnut anthracnose disease is a kind of serious disease of walnut all over the world. In this paper,the identification and the biological characteristics of strain TS-09 isolated from the walnut fruits were studied. The results showed that the strain TS-09 was identified as Colletotrichum gloeosporioides based on its morphologic characters,cultural characteristics and four conserved genes(ribosomal DNA-ITS,ACT,β-tub2 and GPDH)sequences analysis and the pathogenicity was powerful. For the growth of strain TS-09,the best medium was PDA. The optimum temperature was 28℃and the optimum pH for mycelium growth was 7.0. The condition of weak acid was favorable for spore production(pH 4.0). During continuous illumination,the mycelium growth was inhibited while the condition of continuous darkness was better for mycelium growth. The mycelium had stronger resistance against temperature and the lethal temperature for mycelium was 60℃(15 min)or 62℃(10 min). Our research expected to offer theory support for prediction and effective control of walnut anthracnose.
Keywords:Colletotrichum gloeosporioides;identification;biology
*通訊作者:Author for correspondence. E-mail:xqwu@njfu.edu.cn
作者簡介:王清海(1978-),男,山東嘉祥人,博士,工程師,主要從事林木病害及生物防治研究. E-mail:wqhhai@126.com
基金項目:山東省科技發展計劃項目:核桃炭疽病新型健康管理模式構建;山東省農業重大應用技術創新課題:生物藥劑在冬棗、核桃生產中的選配與應用;江蘇高校優勢學科建設工程項目(PAPD)
收稿日期:2015-05-10修回日期:2015-08-23
中圖法分類號:S436.621.1
文獻標識碼:A
文章編號:1000-2324(2016)01-0009-06
猜你喜歡
陜西學前師范學院學報(2022年10期)2022-10-31 14:23:44
中國飼料(2021年17期)2021-11-02 08:15:10
現代畜牧科技(2021年9期)2021-10-13 06:38:54
現代畜牧科技(2021年4期)2021-07-21 06:13:00
生物學通報(2021年5期)2021-03-16 06:42:16
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
數學物理學報(2017年2期)2017-06-05 09:12:30
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:07
醫學研究雜志(2015年11期)2015-06-10 06:44:03
