膽脂瘤上皮細胞中Caspase-9及Smac的表達變化及其相關性
2016-03-23 07:39:38劉東亮馬秀嵐
山東醫藥 2016年1期
劉東亮,馬秀嵐
(中國醫科大學附屬盛京醫院,沈陽110000)
?
膽脂瘤上皮細胞中Caspase-9及Smac的表達變化及其相關性
劉東亮1,馬秀嵐2
(中國醫科大學附屬盛京醫院,沈陽110000)
摘要:目的探討膽脂瘤上皮細胞中Caspase-9和Smac的表達變化及其相關性。方法 選取16例膽脂瘤樣本及10例正常對照樣本。應用免疫組化法及Western blotting法檢測Caspase-9和Smac的表達。采用Spearman檢驗分析Caspase-9和Smac的相關性。結果膽脂瘤上皮細胞中Caspase-9及Smac蛋白表達較正常對照明顯降低(P均<0.01),Smac及Caspase-9的表達水平呈正相關(r=0.744,P<0.01)。結論 膽脂瘤上皮細胞中Smac和Caspase-9的表達降低,Smac及Caspase-9的表達呈正相關。
關鍵詞:膽脂瘤;Caspase-9;Smac;信號傳導通路
膽脂瘤是以上皮細胞增殖分化形成的角化上皮堆積為特點的慢性中耳疾病,如不積極治療,可進行性破壞中耳及周圍重要結構(如面神經、顱底結構等),導致聽力下降、面癱等,甚至是嚴重性的顱內并發癥。膽脂瘤雖具有腫瘤的某些特性,但它并非真正的腫瘤。無凋亡抑制是其與惡性腫瘤區別的一個重要因素。Caspase-9是內源性凋亡途徑的起始因子,而Smac是內源性凋亡時由線粒體釋放的促凋亡因子。我們檢測膽脂瘤上皮細胞中Caspase-9和Smac的表達,并分析了其相關性,來初步探討內源性凋亡在膽脂瘤中的作用。
1資料與方法
1.1臨床資料取2012~2014年于中國醫科大學附屬耳鼻咽喉頭頸外科診斷并行手術治療的16例膽脂瘤患者的上皮組織標本(膽脂瘤組),其中患者男6例、女10例,年齡10~60(30.75±17.92)歲,均有1個月至40年的慢性耳流膿病史,標本均經病理證實為膽脂瘤;10例對照組標本來自于同期因耳后或耳廓腫物接受手術治療的患者安全緣皮膚,病理證實無腫瘤細胞。
1.2上皮細胞中Caspase-9和Smac蛋白的檢測 ①蛋白定性表達:采用免疫組化法。取新鮮膽脂瘤組織及安全緣皮膚,一部分送病理證實,余浸入甲醛溶液中保存,擇期包埋,制作石蠟標本。按照常規SABC法進行檢測,設空白對照。上述步驟中用己知陽性切片作為陽性對照,用PBS緩沖液代替一抗作為陰性對照,以排除非特異性著色。結果判定:細胞核或胞質內出現棕黃色顆粒者為陽性細胞。在陽性細胞標準基礎上,綜合染色強度及陽性細胞數兩個方面進行分析。先按染色強度計分:O分為沒有染色,1分為微弱棕黃色,2分為中等強度棕黃色,3分為深棕色;然后每張400倍顯微鏡下隨機選取5個不重疊視野進行陽性細胞計數,每個視野計數200個細胞,計算5個視野的陽性細胞的平均百分比,按照百分比計分: 0~5%為0分,6%~25%為1分,26%~50%為2分,51%~75%為3分,>75%為4分。每張切片隨機取3個400倍視野,每個視野均進行染色強度計分與陽性細胞百分比計分,計算表達積分(染色強度計分與陽性細胞百分比計分乘積),并規定此積分:0分為陰性(-),1~4分為弱陽性(+),5~8分為中度陽性(++),9~12分為強陽性(+++)。②蛋白定量表達:采用Western blotting法。將少量組織塊(100 mg左右)置于 1~2 mL 勻漿器中球狀部位,加400 μL 單去污劑裂解液(含PMSF)于勻漿器進行勻漿,裂解30 min 后,即可用移液器將裂解液移至1.5 mL離心管中,然后在4 ℃、1 2000 r/min離心5 min,取上清分裝于0.5 mL離心管中并置于-20 ℃保存。根據目的蛋白的分子量,配制10% SDS-PAGE膠垂直電泳,根據蛋白濃度調整上樣量,保持每孔40 μg蛋白上樣。電泳結束后轉移到PVDF膜上。用5%脫脂奶粉常溫下封閉搖床l h,PBS液洗膜3次,每次15 min,然后加入按說明書要求稀釋的Caspase-9和Smac和內參GAPDH抗體,于4 ℃孵育過夜。TBST液洗膜3次,每次15 min,加入二抗,室溫下搖床l h。TBST液洗膜3次,每次15 min,用ECL顯色液顯色,定影液終止顯色。各實驗重復3次。
2結果
2.1各組Caspase-9、Smac的表達對照組Smac表達局限于基底部增生區細胞胞質中,越靠近角質層其表達越弱或呈陰性表達。膽脂瘤上皮無或弱陽性(-~+)表達,主要集中在顆粒細胞層。對照組Caspase-9局限于基底部增生區細胞胞質中,越靠近角質層其表達越弱或呈陰性表達。膽脂瘤上皮無或弱陽性(-~+)表達,主要集中在顆粒細胞層和透明細胞層。膽脂瘤組Smac的表達為-、+、++、+++分別為0、6、4、0例,對照組分別為5、11、0、0例。膽脂瘤上皮的Smac表達明顯弱于對照組(χ2=4.975,P<0.05)。膽脂瘤組Caspase-9的表達為-、+、++、+++分別為0、6、4、0例,對照組分別為6、10、0、0例。膽脂瘤組Caspase-9表達明顯弱于對照組(χ2=10.156,P<0.01)。膽脂瘤組、對照組Caspase-9表達分別為0.47±0.11、0.87±0.14,Smac表達分別為0.23±0.12、0.71±0.13。膽脂瘤組Caspase-9、Smac的表達均低于對照組(P均<0.05)。見圖1。
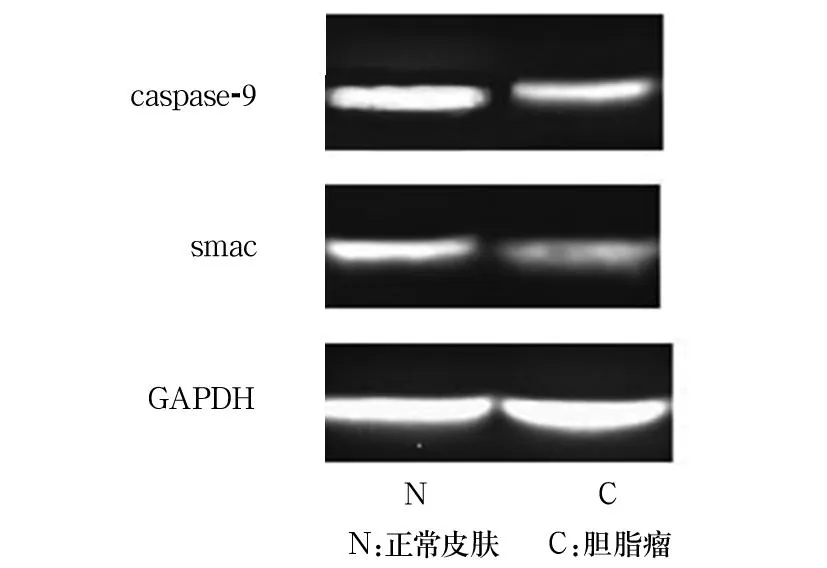
圖1 Western blotting法檢測Caspase-9及Smac的表達
2.2Smac及Caspase-9表達的相關性Smac與Caspase-9的表達呈明顯正相關(r=0.744,P<0.01)。
3討論
中耳膽脂瘤為囊袋狀結構,囊內壁為復層鱗狀上皮,后天原發或繼發于中耳的膽脂瘤上皮多來源于鼓膜上皮層,形成的囊內充滿脫落上皮、角化物質和膽固醇結晶。人們通過研究膽脂瘤上皮細胞中增殖標志物,試圖證明膽脂瘤囊內壁復層鱗狀上皮高度增生的特性。這些標志物包括細胞角蛋白[1]、增殖細胞核抗原[2]、表皮生長因子[3]、Ki67[4]、HMGB-1[5]等。最后這些結果都支持膽脂瘤上皮細胞具有高度增殖的行為,且這種增殖主要以基底層為主。
在一個多細胞機體中,細胞分裂增殖與凋亡是對立統一的,共同協調著組織和器官形態發生的發展方向,使組織細胞數處于動態平衡。在中耳膽脂瘤中,其鱗狀上皮具有高度增殖現象已經被公認,這與腫瘤相似,但是中耳膽脂瘤上皮細胞仍具有凋亡能力,從而使中耳膽脂瘤上皮的細胞處于相對的動態平衡,高速產生大量的角化碎片。
目前認為細胞凋亡的發生主要集中在三條途徑:外源性凋亡途徑(死亡受體途徑)、內源性凋亡途徑(線粒體凋亡途徑)及內質網應激途徑[6~8]。Caspase 家族成員大多都是凋亡的啟動子或效應子, 在細胞凋亡過程中發揮重要作用[9]。目前已經發現的有14 種Caspase蛋白,其中Caspase-3 起著關鍵作用,而且也是目前為止在Caspase家族中研究比較透徹的一個,它是最主要的效應因子,一旦被激活,即發生下游的級聯反應,則凋亡便不可避免,于是Caspase-3 也被稱為“死亡蛋白酶”[10]。正常情況下,胞質中的Caspase-3 以無活性的酶原形存在, 細胞凋亡信號的出現可導致Caspase-3 在多種蛋白水解酶的作用下,發生裂解而活化[11~13]。
凋亡的執行過程是一系列Caspase蛋白級聯切割的過程。不同蛋白酶分別切割并激活Caspase-3 酶原。活化的Caspase-3 又進一步切割不同的底物,導致蛋白酶級聯切割放大,最終使細胞走向死亡。在內源性凋亡途徑中,由于生長激素撤退、射線輻射等凋亡因素刺激后,引起BH3-Only蛋白中促凋亡因子較抗凋亡因子成分增加,線粒體膜改變(如離子通道開放、膜電位下降及通透性增高等),線粒體膜間隙的細胞色素C等釋放,釋放到胞質中的細胞色素C與Apaf-1及dATP結合,形成的復合物募集pro-Caspase-9,形成凋亡復合體,pro-Caspase-9自動活化,Caspase-9為線粒體凋亡的起始因子,活化的Caspase-9剪切并將執行Caspase(如Caspase-3, -6 and -7)活化,從而誘導細胞凋亡[14~17]。凋亡抑制蛋白(IAPs)是Caspase家族的抑制因子,該家族成員均含有1~3個桿狀病毒IAP重復序列,利用此結構區與Caspase-8,7,9結合而發揮抑制凋亡的作用[16]。Smac蛋白正常時也位于線粒體膜間隙,在凋亡誘導因子的作用下,同細胞色素C一起釋放人胞質,與凋亡IAPs結合并解除其對Caspases的抑制作用,從而使Caspase活性增強,促進細胞凋亡發生[17]。
本實驗結果中我們發現Smac與Caspase-9在膽脂瘤上皮細胞中的表達均弱于對照組,說明以Caspase-9為起始因子的內源性凋亡途徑在膽脂瘤上皮細胞凋亡的過程中是被抑制的,這可能與膽脂瘤的病因及刺激因素有關,因為內源性凋亡的刺激因素主要為生長因子撤退、癌基因激活、DNA損傷、化療和射線等。本病例中患者均有慢性中耳炎病史,而在炎癥反應過程中炎細胞釋放的白介素、腫瘤壞死因子等恰恰是誘導外源性凋亡的重要細胞因子;本實驗中發現對照組上皮細胞的Smac及Caspase-9均有陽性表達,且均以基底層為主,說明皮膚正常細胞確實存在增殖與凋亡的負反饋作用,使基底細胞在迅速增殖的過程中凋亡相關因子亦增加,阻止基底細胞的過度增殖。無論在對照組亦或是膽脂瘤組,Caspase-9與Smac的表達水平呈正相關,證實了Smac通過結合IAP,阻斷IAP對Caspase-9的抑制作用,增強了Caspase-9的促凋亡作用。但是由于IAP不僅能結合Caspase-9,也能結合其他凋亡相關的Caspases,所以Smac結合IAP后,亦能增強其他Caspases的凋亡活性。
總之,本研究結果顯示,與正常上皮比較,內源性凋亡在膽脂瘤中是明顯受到抑制的,通過解除這種抑制從而達到對膽脂瘤的藥物治療效果,可能將會是今后努力的方向。
參考文獻:
[1] Olszewska E, Sudhoff H. Comparative cytokeratin distribution patterns in cholesteatoma epithelium[J]. Histol Histopathol, 2007,22(1):37-42.
[2] Raffa S, Leone L, Scrofani C, et al. Cholesteatoma-associated fibroblasts modulate epithelial growth and differentiation through KGF/FGF7 secretion[J]. Histochem Cell Biol, 2012,138(2):251-269.
[3] Thorup MB, Munk M, Poulsen SS, et al. Expression of the epidermal growth factor system in human middle ear cholesteatoma[J]. Acta Otolaryngol, 2014,134(2):124-134.
[4] Kuczkowski J, Pawelczyk T, Bakowska A, et al. Expression patterns of Ki-67 and telomerase activity in middle ear cholesteatoma[J]. Otol Neurotol, 2007,28(2):204-207.
[5] Chi Z, Wang Z, Liang Q, et al. Induction of cytokine production in cholesteatoma keratinocytes by extracellular high-mobility group box chromosomal protein 1 combined with DNA released by apoptotic cholesteatoma keratinocytes[J]. Mol Cell Biochem, 2015,400(1-2):189-200.
[6] Kiechle FL, Zhang X. Apoptosis: biochemical aspects and clinical implications[J]. Clin Chim Acta, 2002,326(1-2):27-45.
[7] Ashe PC, Berry MD. Apoptotic signaling cascades[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2003,27(2):199-214.
[8] Liu H, Baliga R. Endoplasmic reticulum stress-associated caspase 12 mediates cisplatin-induced LLC-PK1 cell apoptosis[J]. J Am Soc Nephrol, 2005,16(7):1985-1992.
[9] Stepień A, Izdebska M, Grzanka A. The types of cell death[J]. Postepy Hig Med Dosw, 2007,61:420-428.
[10] Poreba M, Strózyk A, Salvesen GS, et al. Caspase substrates and inhibitors[J]. Cold Spring Harb Perspect Biol, 2013,5(8):a008680.
[11] Galluzzi L, Kepp O, Kroemer G. Caspase-3 and prostaglandins signal for tumor regrowth in cancer therapy[J]. Oncogene, 2012, 31(23):2805-2808.
[12] Snigdha S, Smith ED, Prieto GA, et al. Caspase-3 activation as a bifurcation point between plasticity and cell death[J]. Neurosci Bull, 2012,28(1):14-24.
[13] Choudhary GS, Al-Harbi S, Almasan A. Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis[J]. Methods Mol Biol, 2015,1219:1-9.
[14] Shi L, Chen J, Yang J, et al. MiR-21 protected human glioblastoma U87MG cells from chemotherapeutic drug temozolomide induced apoptosis by decreasing Bax/Bcl-2 ratio and caspase-3 activity[J]. Brain Res, 2010,1352:255-264.
[15] Kuida K. Caspase-9[J]. Int J Biochem Cell Biol, 2000,32(2):121-124.
[16] Yang D, Song X, Zhang J, et al. Therapeutic potential of siRNA-mediated combined knockdown of the IAP genes (Livin, XIAP, and Survivin) on human bladder cancer T24 cells[J]. Acta Biochim Biophys Sin, 2010,42(2):137-144.
[17] Du C, Fang M, Li Y, et al. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition[J]. Cell, 2000,102(1):33-42.
《山東醫藥》第55卷第45期73頁“塵螨過敏哮喘患兒舌下含服粉塵螨滴劑后血清塵螨SlgE、SlgG4及IL-12、IL-13水平變化”一文應為上海市嘉定區衛生局青年科研基金立項項目(QNKYJJ 2012-12-02)。
本刊編輯部
(收稿日期:2015-03-09)
中圖分類號:R735.8
文獻標志碼:B
文章編號:1002-266X(2016)01-0049-03
doi:10.3969/j.issn.1002-266X.2016.01.018
通信作者:馬秀嵐(E-mail:subtlety@126.com)
