棘孢木霉對黃花蒿葉的光合特性和產量影響
2016-03-03 04:22:59楊興堂呂曼曼劉志華朱國棟馬德志張榮沭
貴州農業科學 2016年1期
楊興堂, 呂曼曼, 劉志華, 朱國棟, 王 慧, 馬德志, 張榮沭*
(1.東北林業大學 園林學院, 黑龍江 哈爾濱 150040; 2.東北林業大學 林學院, 黑龍江 哈爾濱 150040; 3.齊齊哈爾醫學院, 黑龍江 齊齊哈爾 161006)
棘孢木霉對黃花蒿葉的光合特性和產量影響
楊興堂1, 呂曼曼1, 劉志華2, 朱國棟1, 王 慧1, 馬德志3, 張榮沭1*
(1.東北林業大學 園林學院, 黑龍江 哈爾濱 150040; 2.東北林業大學 林學院, 黑龍江 哈爾濱 150040; 3.齊齊哈爾醫學院, 黑龍江 齊齊哈爾 161006)
為了對黃花蒿的增產和促進木霉免疫誘導劑類生物肥料在植物生長中的應用提供理論依據,在大田條件下進行根施1×105cfu/mL(T1)、1×106cfu/mL(T2)和1×107cfu/mL(T3)3個水平的棘孢木霉ACCC30536分生孢子對黃花蒿葉的光合特性和產量的影響試驗。結果表明:T3水平(200 mL/株)的木霉菌對黃花蒿的誘導效果最佳;根施木霉孢子量與青蒿葉片凈光合速率(Pn)呈正相關,光合“午休”現象有一定程度的減緩;T3處理黃花蒿的光合-光響應曲線參數最大凈光合速率、表觀量子效率、暗呼吸速率及光飽和點、光補償點均高于CK和其他處理。并且,木霉菌誘導黃花蒿60 d后T3組黃花蒿葉的產量較CK提高最大。說明,棘孢木霉ACCC30536能夠改善黃花蒿的光合能力,促進干物質的積累,從而提高其葉的產量。
棘孢木霉; 黃花蒿; 光合特性; 光響應; 葉產量
國際上應用非常普遍的木霉(Trichoderma)生防真菌能定殖在植物根際,具有增產、防治病害和改良土壤的作用[1-2]。木霉菌能通過提高植株的光合作用及增加土壤養分溶解促進植物生長,提高其產量。同時,發育健壯的植物抗病能力提升[3-4]。植物的生長是通過光合作用固定有機物積累干物質來實現的,所以提高植物的光合作用有助于其生物量的積累。黃花蒿(ArtemisiaannuaL.)為菊科一年生草本植物,是我國的傳統中藥,具清熱涼血、退虛熱,解暑的功效,其提取物青蒿素有顯著的抗瘧、解熱鎮痛作用,目前已替代了傳統的奎寧類抗瘧藥物,被廣泛應用于臨床治療各型瘧疾[5-6]。迄今為止,青蒿素的生產仍主要依靠從人工種植的黃花蒿葉中提取[7]。隨著青蒿素市場需求量的不斷增大,如何提高栽培黃花蒿的產量已成為人們研究的熱點。棘孢木霉ACCC30536能顯著改善山新楊的光合特性[8]。但采用木霉菌誘導黃花蒿提高其產量的研究報道極少。因此,筆者采用棘孢木霉ACCC30536分生孢子根施黃花蒿,分析木霉菌對黃花蒿葉的光合特性和產量的影響,為木霉免疫誘導劑類生物肥料在促進黃花蒿生長,提高其葉產量的應用提供理論依據。
1 材料與方法
1.1 供試材料
黃花蒿種子采自重慶酉陽,按張榮沭[9]的方法進行溫室紙筒育苗。棘孢木霉ACCC30536(T.asperellumACCC30536)菌株由東北林業大學林學院森保學科提供。
1.2 試驗設計
木霉分生孢子培養采用PDA固體培養基(主要成分為馬鈴薯、葡萄糖和瓊脂粉,pH自然),將活化的木霉分生孢子在無菌條件下接種到培養皿中,在28℃條件下培養6 d獲得大量分生孢子。用自來水配制成高濃度的孢子懸液,顯微鏡下計數,測得其濃度,用自來水將其稀釋為1×105cfu/mL、1×106cfu/mL和1×107cfu/mL 3個水平的木霉孢子懸液。
2014年5月10日黃花蒿幼苗從溫室移栽至哈爾濱市郊大田,株間距30 cm,行間距60 cm。小區面積18 m2(6 m×3 m)。7月10日將1×105cfu/mL(T1)、1×106cfu/mL(T2)和1×107cfu/mL(T3)的木霉孢子懸液根施黃花蒿,200 mL/株,對照(CK)施用等體積的自來水,3次重復。試驗期間管理措施相同,黃花蒿生長期間保持土壤水分充足,定期除草、松土。
1.3 測定項目與方法
1.3.1 光合作用日變化的測定 8月中旬,選擇晴朗無風的天氣,各試驗組中選3株長勢一致的黃花蒿,在其近頂端選取3片健壯的成熟葉片測定凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Cond)和胞間CO2濃度(Ci)的日變化。測定儀器為便攜式光合作用分析儀(Li-6400XT,美國LI-COR公司),采用開放式氣路,CO2濃度約為350 μL/L左右,自然光照,8:30-16:30每隔2 h測定1次。
PLL電路是一個相位負反饋環路,由鑒相器、環路濾波器和壓控振蕩器(VCO)組成,它將輸入信號與VCO輸出信號的相位進行比較,控制兩個信號使其保持同相位。若在PLL輸入信號中加上由晶振等產生穩定的頻率信號,在VCO輸出與鑒相器輸入之間接入分頻器,通過切換分頻器的頻率,便得到VCO的輸出與輸入頻率同樣精度的分頻信號[7]。這就是PLL合成頻率源的工作原理,如圖1所示。
1.3.2 光響應曲線的測定和參數分析 8月中旬選擇3個晴天,在9:00-12:00期間,隨機選取各試驗組的黃花蒿近頂部無病蟲害、朝向一致的健壯成熟葉片,用內置6400-02 LED紅藍光源和6400-01CO2注入系統的Li-6400XT便攜式光合測量系統,進行連體葉片光合速率測量。光合有效輻射設定的梯度為12個:2 000 μmol/(m2·s),1 800 μmol/(m2·s),1 500 μmol/(m2·s),1 200 μmol/(m2·s),1 000 μmol/(m2·s),800 μmol/(m2·s),600 μmol/(m2·s),400 μmol/(m2·s),200 μmol/(m2·s),100 μmol/(m2·s),50 μmol/(m2·s),20 μmol/(m2·s),以不照射為對照。CO2濃度為350 μL/L,葉面溫度由Li-6400溫度控制器控制在(30±1)℃。采用非直角雙曲線模型[10-11]按照公式(1)利用SPSS統計軟件對測定數據結果進行擬合,從而計算最大凈光合速率(Pn)、表觀量子效率(AQY)和暗呼吸速率(Rd)等光合-光響應曲線參數。光飽和點(LSP)和光補償點(LCP)的計算方法是將0~200 μmol/(m2·s)低光強下的光響應曲線進行線性回歸分析。直線與X軸(PAR)的交點就是LCP;該直線與Y=Pnmax直線相交,交點所對應X軸的數值即為LSP。
式中,Pn為凈光合速率,E為光照強度(PAR),Pnmax為最大凈光合速率,Q為表觀量子效率(AQY),k為曲角,Rd為暗呼吸速率。
1.3.3 黃花蒿葉產量的測定 木霉誘導60 d后,測量黃花蒿葉的產量。將各試驗組黃花蒿的葉片從其枝條上分離稱量鮮重;稱重后,將葉片按照文獻[12]的方法烘干,再分別稱量干重。
1.4 數據分析
利用Microsoft Office Excel 2007處理數據和繪制圖表。用SPSS18.0和Minitab 16.0軟件包進行統計分析及在P=0.05下比較差異顯著性。
2 結果與分析
2.1 黃花蒿光合生理因子的日變化
2.1.1 凈光合速率(Pn) 3個水平木霉菌誘導后黃花蒿與CK的凈光合速率變化趨勢基本一致,均呈雙峰型曲線,峰值分別出現在10:30和14:30,說明黃花蒿存在午休現象(圖1)。并且Pn在14:30的峰值基本上大于10:30的峰值。T3處理的光合速率在8:30-16:30明顯高于其他處理和CK,且在12:30處的T3>T2>T1>CK,說明,根施木霉菌既能提高黃花蒿的光合作用又能緩解光合午休現象(圖1),提升了楊樹苗對光的生態適應能力。并且木霉菌處理提高黃花蒿光合能力與其施用的木霉分生孢子的劑量有關,最佳施用劑量為1×107cfu/mL(200 mL/株)。
2.1.2 氣孔導度(Cond) 從圖1看出,3個水平木霉菌誘導后黃花蒿與CK的氣孔導度日變化規律基本一致,并且在不同程度上增強了黃花蒿的氣孔導度。其中,T3和T2處理對黃花蒿氣孔導度的影響較大。說明,木霉菌對黃花蒿氣孔導度的影響也與施用的木霉劑量相關。在12:30由于氣溫升高,蒸騰速率增大,木霉處理組的氣孔導度增量較其他時間點減小。因為植物是通過改變氣孔數目和開閉程度來調節葉片的蒸騰速率和水勢,并且光合作用和干物質積累所需要的二氧化碳也是通過氣孔而獲得,所以氣孔在調控水分丟失和光合作用之間總是處于一種折中狀態。說明根施適量的木霉菌能夠使黃花蒿提高自身與環境對水分和CO2的交換能力,有利于黃花蒿調整水分以適應環境溫度和攝入CO2,提高其對干物質的積累。
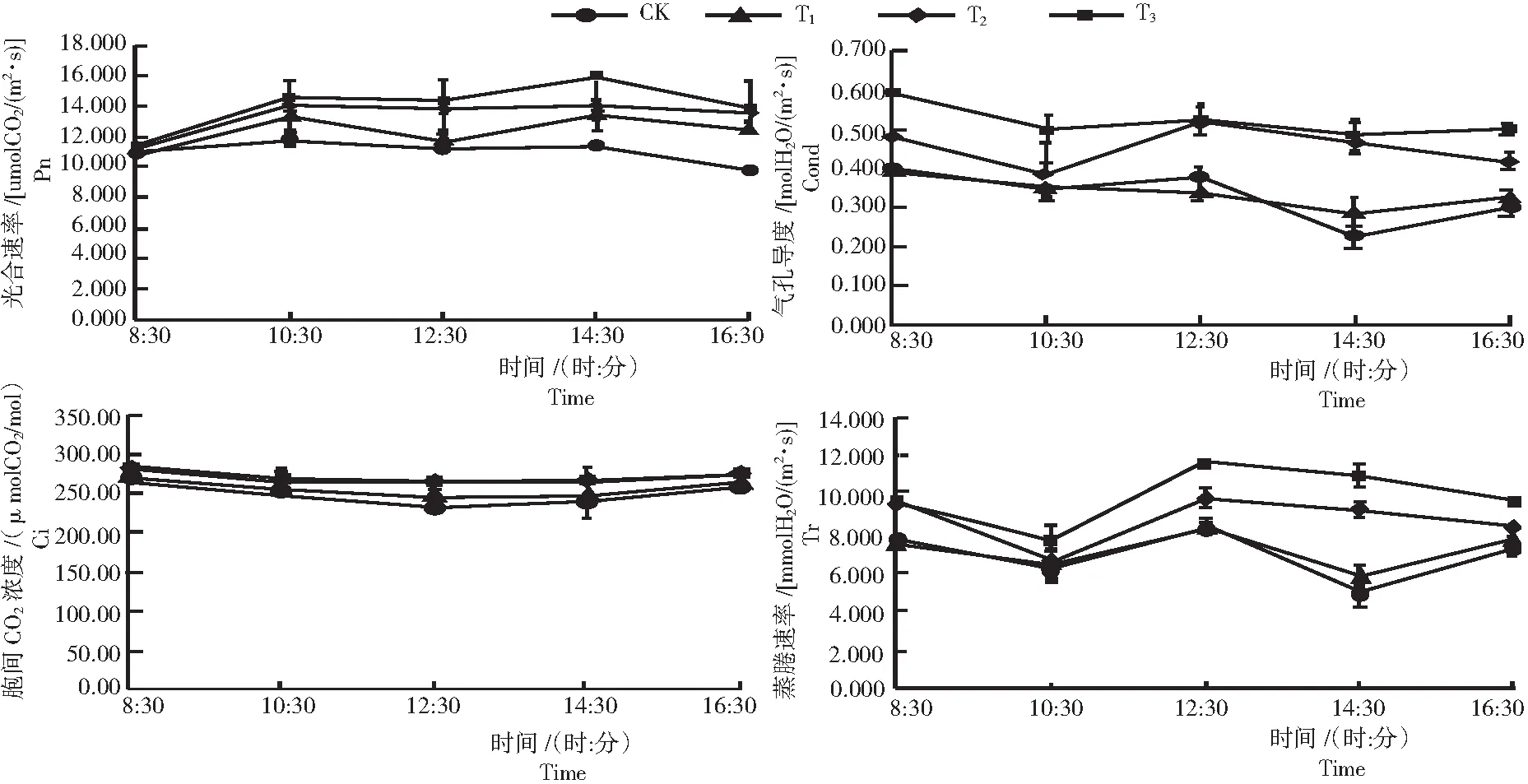
圖1 不同處理黃花蒿葉光合生理因子的日變化
2.1.3 胞間CO2濃度(Ci) 從圖1表明,3個水平木霉菌誘后黃花蒿和CK的胞間CO2濃度日變化規律基本一致。檢測初期的胞間CO2濃度較高,之后一直下降,到12:30降到最低點,再一直升高。木霉菌處理組黃花蒿的Ci均比CK高,并且T3和T2水平的木霉誘導效果相近,在10:30-14:30Ci變化不大。說明,木霉菌能夠提高胞間CO2濃度,有利于黃花蒿光合作用的進行,增加干物質的積累,從而提高葉的產量。
2.2 黃花蒿的光響應曲線參數
根據圖2、圖3和表的結果:
1) 木霉菌可以提高黃花蒿葉片最大凈光合速率,并且黃花蒿葉片最大凈光合速率提高與施用的木霉分生孢子量的大小呈正相關,即最大凈光合速率T3>T2> T1>CK,差異顯著(P<0.05)。最大凈光合速率較高有利于干物質的積累,能為提高植物產量奠定良好的生理基礎。
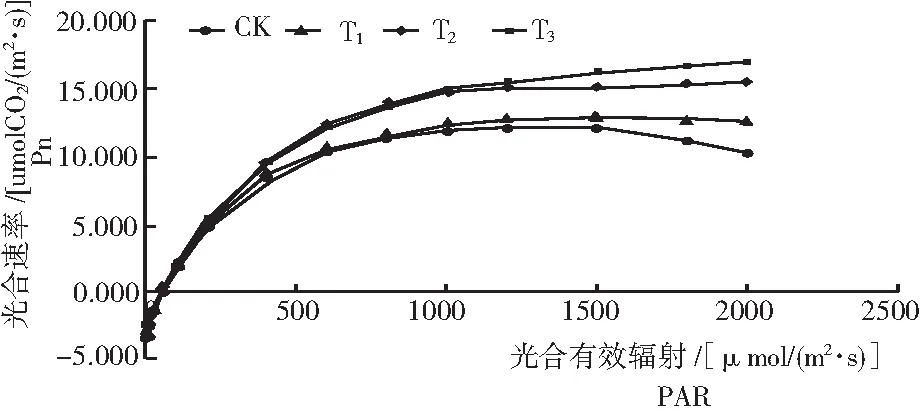
圖2 黃花蒿葉的光合速率的光響應
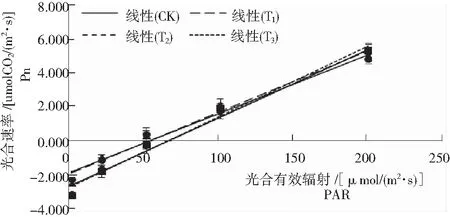
圖3 低光強下黃花蒿的Pn-PAR擬合直線[PAR﹤200/μmol(m2·s)]
Fig.3 Pn-PAR Fitted lines ofA.annualeaf under low light intensity
2) 表觀量子效率(AQY)是光合作用中光能轉化效率的指標之一,是凈光合速率與相應光量子通量密度的比值。T1和T2水平木霉菌誘導后黃花蒿葉片的AQY值差異不大,但T3處理顯著高于其他試驗組,為0.055 μmol/mol。說明,適量的木霉孢子誘導可以提高黃花蒿對光能的轉化效率。
3) 暗呼吸速率(Rd)反映植物在黑暗條件下的
呼吸速率。采用不同水平木霉分生孢子處理的黃花蒿,其葉片的暗呼吸速率隨著施用木霉分生孢子水平的增加而增大。然而,暗呼吸速率小才有利于植物在低Pn條件下保持碳平衡。

表 黃花蒿葉片的光合光響應曲線模擬參數
注:每列中不相同字母表示在P< 0.05水平上顯著性差異。
Note: Different letters in the same column indicate significance of difference atP< 0.05 level.
4) 光飽和點(LSP)是植物利用強光能力大小的指標。T2和T3的光飽和點(LSP)分別是CK的1.07倍和1.05倍,均顯著大于CK和T1(P<0.05)。說明,采用T2、T3水平的木霉分生孢子誘導黃花蒿,提高了植物對光的生態適應能力。
5) 光補償點(LCP)是植物利用弱光能力大小的重要指標,該值越小表明利用弱光的能力越強。具有高LSP與低LCP的植物,對光的生態適應能力強,受到強光刺激時不易發生抑制。雖然本試驗中黃花蒿的LCP值表現出與Rd值一致的變化規律,但施用不同水平木霉誘導后,T2和T3組黃花蒿LSP與LCP的差值相近,分別為1 368.09和1 346.62;T1和CK組黃花蒿LSP與LCP的差值相近,分別為1 267.13和1 286.05。說明,T2和T3木霉分生孢子誘導黃花蒿,能顯著提高黃花蒿對光的生態適應能力。
2.3 黃花蒿葉的產量
試驗結果表明,不同濃度的棘孢木霉分生孢子誘導黃花蒿60 d后,T1、T2、T3和對照葉片的干重分別為(68.34±8.25)g/株、(73.96±8.52)g/株、(83.56±9.81)g/株和(66.38±8.92)g/株。說明,隨著木霉分生孢子濃度的增加,黃花蒿生物量產量也呈上升趨勢。T3和T2處理單株黃花蒿葉的平均干重與CK和T1相比存在顯著差異(P<0.05),分別是CK的1.26和1.11倍。表明,施用適量棘孢木霉分生孢子具有促進黃花蒿的生長的作用。
3 結論與討論
1) 黃花蒿除了具有抗瘧疾、殺蟲、抗癌、清熱解毒等藥理作用外,還能用于調節植物生長。黃花蒿的有效藥用成分為葉組織,因此,提高葉組織的產量是栽培黃花蒿獲取大的經濟效益追求的目標之一。在以往對影響黃花蒿產量因素的研究中,多以施用化肥改變土壤中氮磷鉀等大量元素含量、鋅錳硼等微量元素含量[13-14],以及采取不同的栽培措施[15]、在不同生長時期改變土壤水份[16]等方法來建立增產的最佳管理辦法。而通過施用木霉菌來提高黃花蒿有效生物量的研究極少。木霉菌(Trichoderma)是一種國際上農林業生產實踐中應用越來越廣泛的生防因子和生物肥料,不僅對植物病原菌具有競爭作用、重寄生作用及抗菌作用,而且還有溶解作用使植物提高對營養物質的吸收和促進植物生長。本試驗采用不同水平的木霉分生孢子溶液根施黃花蒿,能使其葉產量提高1.26倍。說明木霉能顯著提高黃花蒿葉的產量。研究發現,植物益生菌能改善土壤環境[2],然而,施入土壤的有益微生物也會影響其原有微生物環境的生態平衡。大量施入土壤中的木霉菌,會打破土壤原有的菌群互生的平衡關系,改變植物賴以生存土壤的微生態環境。因此,木霉的施用劑量對土壤的改良效果不同。本研究發現,施用濃度為1×107cfu/mL (200 mL/株)的木霉分生孢子,能獲得最高的黃花蒿葉產量。
2) 植物是通過光合作用獲取能量來合成有機物,可見高效的光合作用對植物積累干物質、增加產量至關重要。研究發現,哈茨木霉能顯著提高番茄植株葉片的光合作用[17]。陸寧海[18]等施用哈茨木霉制劑通過增加葉片的葉綠素含量來提高光合作用能力,使小麥和玉米幼苗生長發育增快。王慧[8]等用5×103cfu/cm3土的棘孢木霉分生孢子誘導山新楊,顯著提高了其光合-光響應能力。說明木霉菌不僅對草本植物而且對木本植物的光合作用均具有積極的影響。本實驗結果表明,施用3個水平木霉菌的黃花蒿最大凈光合速率明顯高于對照,并且對黃花蒿光合作用的影響與施入木霉的劑量相關。T3處理的最大凈光合速率,光飽和點和表觀初始量子效率均有顯著提高。并且T3和T2的LSP與LCP的差值顯著大于T1和Con組,改善了黃花蒿對光的生態適應能力,說明根施木霉后的黃花蒿提高了對光能的利用,有利于其產量的增加。
3) 隨著醫藥市場對黃花蒿的藥效成分青蒿素需求量的增加,黃花蒿的需求量也隨之提高[19]。本試驗結果表明:施加棘孢木霉ACCC30536能顯著影響黃花蒿光合特性、提高其藥用生物量產量,在大面積種植的前提下,施用木霉菌肥增加的收入將非常可觀。由于青蒿素產量是由黃花蒿生物量產量和青蒿素含量所共同決定的,提高黃花蒿生物量產量只是增加青蒿素產量的一種途徑,木霉菌對黃花蒿中青蒿素含量的影響也極為重要,相關內容還需作進一步研究。
[1] Martinez-Medina A,Alguacil M D,Pascual J A,et al.Phytohormone profiles induced by trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants[J].Journal Of Chemical Ecology,2014,40(7):804-815.
[2] Yadav R L,Shukla S K,Suman A, et al.Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions[J].Biology Fertility of Soils,2009,45(5):461-468.
[3] Siddiqui Y,Meon S,Ismail R,et al.Bio-efficiency compost extracts on the wet rot incidence,morphological and physiological growth of okra(Abelmoschus esculentus[(L.)Moench])[J].Scientia Horticul Turae,2008,117(1),9-14.
[4] Tripathi P,Singh P C,Mishra A,et al.Trichoderma:a potential bioremediator for environmental clean up[J].Clean Technologies and Environmental Policy,2013,15(4):541-550.
[5] Nicholas S,Huahong W,Werner RM,et al.Artemisinin biosynthesis in growing plants of Artemisia annua.A13CO2study[J].Phytochemistry,2010,17(2-3):179-187.
[6] Patrick S C,Keat H T,Devin R P,et al.Functional genomics and the biosynthesis of artemisinin[J].Phytochemistry, 2007,68(14):1864-1871.
[7] Han J L,Wang H,Ye H C, et al.High efficiency of genetic transformation and regeneration ofArtemisiaannuaL.via Agrobacterium tumefaciens-mediated procedure[J].Plant Science,2005,168(1):73-80.
[8] 王 慧,劉志華,呂曼曼,等.棘孢木霉菌對山新楊移栽苗葉片光合特性的影響[J].北方園藝,2013,23(1):78-82.
[9] 張榮沭.大緯度引種青蒿藥效、產量影響因子及藥效分析方法學研究[D].哈爾濱:東北林業大學,2008.
[10] 譚念童,林 琪,李玲燕,等.限量補灌對旱地冬小麥灌漿期旗葉光響應及產量的影響[J].華北農學報,2010,25(4):145-151.
[11] 許 賢,王貴啟,樊翠芹,等.外來入侵植物黃頂菊生長及光合特性[J].華北農學報,2010,25(Z):133-138.
[12] Zhang F G,Yuan J,Yang X M,et al.Putative Trichoderma harzianum mutant promotes cucumber growth by enhanced production of indole acetic acid and plant colonization[J].Plant and Soil,2013,368(1-2):433-444.
[13] 韋美麗,黃天衛,陳中堅,等.不同氮磷鉀用量對黃花蒿產量的影響[J].現代中藥研究與實踐,2009,23(1).9-10.
[14] 吳葉寬,李隆云,馬 鵬,等.鋅錳硼對黃花蒿產量和青蒿素含量的影響[J].中國中藥雜志,2010,35(3):275-278.
[15] 韋 霄,李 鋒,許成瓊,等.不同栽培措施對黃花蒿產量和青蒿素含量的影響[J].廣西科學院學報,1999,15(3):37-41.
[16] 孫年喜,李隆云,鐘國躍,等.不同生長期土壤水分處理對黃花蒿生理特性及產量的影響[J].中國中藥雜志,2009,34(4):386-389.
[17] Alexandru M,Lazar D,Ene M,et all.Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes[J].Romanian Biotechnological Letters,2013,18(4):8499-8510.
[18] 陸寧海,徐瑞富,房鎮宏,等.哈茨木霉對小麥和玉米幼苗生長的影響[J].江蘇農業學報,2005,21(3):238-240.
[19] 陳思安,韓 頌,趙 敏.產青蒿素作物黃花蒿的研究現狀及展望[J].黑龍江醫藥,2010,23(6):891-894.
(責任編輯: 聶克艷)
Effects ofTrichodermaasperellumon Leaf Photosynthetic Characteristics and Yield ofArtemisiaannua
YANG Xingtang1, LU Manman1, LIU Zhihua2, ZHU Guodong1,WANG Hui1, MA Dezhi3, ZHANG Rongshu1*
(1.CollegeofLandscapeArchitecture,NortheastForestryUniversity,Harbin,Heilongjiang150040; 2.SchoolofForestry,NortheastForestryUniversity,Harbin,Heilongjiang150040; 3.QiqiharMedicalCollege,Qiqihar,Heilongjiang161006,China)
The photosynthetic characteristics and leaf yield ofArtemisiaannuarespectively applying with 1 105cfu/mL(T1), 1×106cfu/mL(T2)and 1×107cfu/mL(T3)T.asperellumACCC30536 conidium by root application in field were studied to provide the theoretical basis for increasingArtemisiaannuayield and promoting application ofTrichodermaas an immune-inducing biofertilizer in plant growth. Results: The inductive effect of T3 treatment onArtemisiaannuais the best among three treatments. The application amount ofTrichodermaasperellumspores is positively related toPnofArtemisiaannualeaves. ApplyingTrichodermaasperellumcan retard the photosynthetic “noon break” phenomenon to some extent. The maximum net photosynthetic rate, apparent quantum yield, dark respiratory, light saturation point and light compensation point ofArtemisiaannuatreated with 1 107cfu/mL(T3)T.asperellumACCC30536 conidium all are higher than CK, T1 and T2 treatment. TheArtemisiaannualeaf yield of T3 treatment increases significantly compared with CK after 60 d. In conclusion, applyingT.asperellumACCC30536 can improve the photosynthetic capacity ofArtemisiaannuaand promote dry matter accumulation, which results in yield increase ofArtemisiaannualeaf.
Trichodermaasperellum;Artemisiaannua; photosynthesis; photoresponse; leaf yield
2015-06-02; 2016-01-07修回
國家自然科學基金資助項目“木霉誘導下楊樹ARF轉錄因子對其生長及抗病的分子調控機制”(31370642);國家自然科學基金資助項目“棘孢木霉的刺激植物響應蛋白Epl1誘導楊樹系統抗病性機制”(31170601);黑龍江省自然科學基金資助項目“木霉菌促進山新楊生長的Auxin信號轉導機制”(C201216)
楊興堂(1990-),男,在讀碩士,研究方向:園林植物種質資源。E-mail:1270314027@qq.com
*通訊作者:張榮沭(1965-),女,副教授,碩士生導師,從事園林植物病害生物防治研究。E-mail:zrs6504@sina.com
1001-3601(2016)01-0035-0132-05
S567.21+9
A
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
