LeftyA基因重組慢病毒表達載體的構建及鑒定
2016-01-07 06:47:56邱宇安,陳文學,靳文劍等
實用癌癥雜志 2015年12期
·基礎研究·
LeftyA基因重組慢病毒表達載體的構建及鑒定
邱宇安陳文學靳文劍陳火國鐘寧余潔麗
作者單位:330029 江西省腫瘤醫院
【摘要】目的想構建LeftyA基因的慢病毒表達載體,鑒定其在293T細胞中的表達。方法首先擴增LeftyA基因序列,將其與慢病毒骨架載體連接后,鑒定慢病毒表達載體構建成功。將重組慢病毒表達載體質粒及包裝質粒共轉染293T細胞,鑒定慢病毒轉染并包裝成功。結果LeftyA基因成功轉導至以GV287為骨架質粒的慢病毒系統中,包裝后測定病毒滴度達2.0×108 TU/ml,免疫印記法檢測到LeftyA蛋白在293T細胞中的表達。結論成功構建了攜帶人LeftyA基因的慢病毒表達系統。
【關鍵詞】LeftyA;慢病毒載體
基金項目:江西省青年科學基金資助項目(編號:20132BAB215018)
DOI:10.3969/j.issn.1001-5930.2015.12.001
中圖分類號:R73-36文獻標識碼:A
收稿日期(2015-10-12修回日期 2015-10-21)
Construction and Indentification of LeftyA Recombinant Lentiviral Expression Vector
QIUYu'an,CHENWenxue,JINWenjian,etal.JiangxiCancerHospital,Nanchang,330029
Abstract【】ObjectiveTo construct LeftyA gene lentiviral vector system and determine its expression in 293Tcells.MethodsFull-length sequence of LeftyA was amplified by PCR,and was connected with lentiviral vector.Gene sequencing was used to identify that recombinant lentiviral vector was constructed successfully.The plasmids were transfected into 293Tcells.Indentification by fluorescent microscopy confirmed lentiviral transfection and packaging successfully.ResultsLeftyA were subcloned into GV287 as skeleton plasmid for a lentivirus packing system.The virus titer was 2.0×108 TU/ml in the supernatant liquid.The proteins of LeftyA were detected by western blot.ConclusionThe LeftyA gene recombinant lentiviral expression system is constructed successfully.
【Key words】LeftyA;Lentiviral vector
(ThePracticalJournalofCancer,2015,30:1755~1757)
我國是胃癌高發區,其發病率及死亡率均明顯高于世界平均水平[1]。本課題前期研究提示體外培養的胃癌細胞株LeftyA基因表達豐度低,LeftyA基因可能對胃癌的發生和發展起著抑制作用。因此,有必要進一步研究LeftyA基因對胃癌細胞功能的影響。我們構建了LeftyA基因重組慢病毒載體,為進一步的功能學研究打下基礎。
1材料與方法
1.1 主要試劑
限制性內切酶(NEB公司),In-FusionTMPCRCloningKit(clontech公司),Taq polymerase (SinoBio公司),dNTP(Takara 公司),Plasmid抽提Kit(Promega 公司),DMEM(GIBCO公司),Lipofectamine2000(Invitrogen公司)。
1.2 主要儀器
PCR儀(Applied Biosysems公司),穩壓DNA電泳儀(BioRad公司),凝膠成像儀(天能公司),細菌搖床(華利達實驗設備公司),高速離心機(日立公司),熒光顯微鏡(奧林巴斯),Plus-20離心超濾裝置(Millipore公司)。
1.3 質粒及細胞株
慢病毒系統包括骨架質粒GV287(pUbi-MCS-3FLAG-SV40-EGFP AgeI /AgeI 酶切),包裝質粒pHelper1.0、pHelper2.0和包裝細胞293T均由上海吉凱基因化學技術有限公司提供。
1.4 引物設計
使用Primer5軟件設計引物,LeftA引物:上游(P1)5'-GAGGATCCCCGGGTACCGGTCGCCACCATGTGGCCCCTGTGGCTCTG-3',下游(P2)5'-TCCTTGTAGTCCATACCTGGCTGGAGCCTCCTTGGCAC-3'。引物含交換配對堿基、酶切位點,并含有目的基因5'端部分序列用于PCR釣取目的基因。PCR產物大小:1142 bp。
1.5 LeftyA慢病毒表達載體的構建
1.5.1PCR擴增目的基因(LeftyA)片段反應條件:94 ℃預變性5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 2 min共30個循環,72 ℃ 10 min。PCR結果見圖1。
1.5.2載體酶切GV287載體原件順序Ubi-MCS-3FLAG-SV40-EGFP,大小10.4 kb,酶切反應溫度37 ℃,酶切反應時間2 h。酶切結果見圖2。
1.5.3重組質粒構建PCR產物交換入線性化載體,用氯化鈣制備新鮮的大腸桿菌感受態細胞,將交換后的連接產物轉化大腸桿菌,將已轉化的感受態細胞轉移到AMP抗性(100 μg/ml)的LB瓊脂培養基上。于37 ℃培養16 h。長出的克隆進行后續PCR鑒定。
1.6 LeftyA慢病毒表達載體的鑒定
引物 (P3)5'-CTGCTGCTACAGGTGTCGGT-3', (FLAG-R-2)5'-CCTTATAGTCCTTATCATCGTC-3'一個位于目的基因中,一個位于載體上,用于菌落PCR鑒定轉化子。陽性轉化子PCR產物大小:574 bp。陽性克隆測序,結果進行同源比對分析。鑒定結果見圖3。
轉染293T細胞:將3種質粒載體(重組病毒質粒及兩種輔助包裝原件載體質粒)分別進行高純度無內毒素抽提,按Lipofectamine 2000試劑盒使用說明進行操作,共轉染293T細胞,熒光顯微鏡下觀察細胞內熒光標記基因(GFP)的表達情況,Westernblot檢測目的基因的表達。測定病毒滴度。鑒定結果見圖4、5。
2結果
2.1 PCR擴增目的基因結果(圖1)
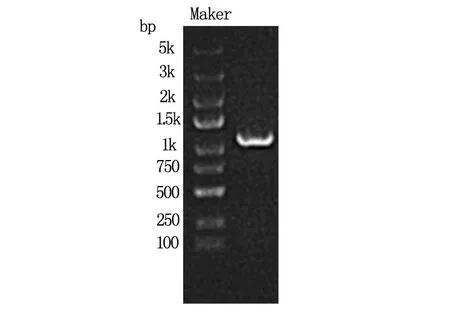
圖1 PCR擴增目的基因(LeftyA)片段
2.2 載體酶切結果(圖2)

1為10kb Marker;2為載體酶切產物,3為未酶切載體。
2.3 陽性轉化子電泳結果(圖3)

1為陰性對照(ddH2O);2為陰性對照(空載自連對照組);3為陽性對照(GAPDH);4為Marker;5~12為LEFTY 1~8號轉化子。
2.4 重組質粒測序結果
將陽性克隆作為模板擴增出目的基因進行測序,將測序結果與目標基因序列進行同源性分析對比。結果顯示重組質粒構建成功,同源性100%。
2.5 重組質粒轉染293T細胞結果(圖4、5)
取培養生長狀態良好的293T細胞,質粒轉染前一天將細胞分入24-well培養板培養,轉染24 h后熒光顯微鏡下觀察細胞內熒光標記基因(GFP)的表達情況,可見細胞內有熒光蛋白表達,提示質粒轉染293T細胞成功。轉染36 h后收集細胞進行Westernblot檢測。可以觀察到43 kD附近處有特征條帶,其大小和目的基因融合蛋白相吻合。
2.6 病毒滴度測定結果
轉染293T細胞后8 h更換為完全培養基,培養48 h后,收集富含慢病毒顆粒的細胞上清液,對其濃縮后得到高滴度的慢病毒濃縮液,在293T細胞中采用ELISA法+熒光法測定并標定病毒滴度,計算出包裝的病毒滴度約為2×108TU/ml。
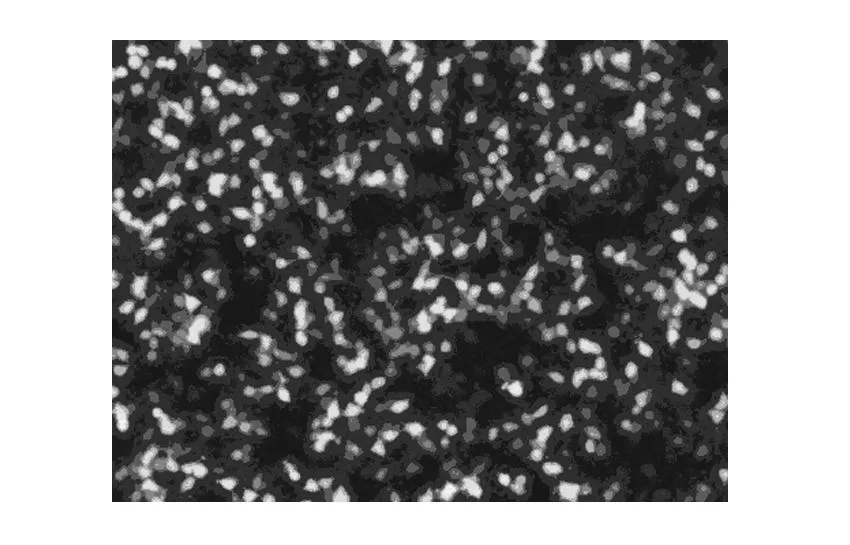
圖4 質粒轉染293T細胞(×100)

1為陽性對照(標準品SURVIVIN-3FLAG-GFP 分子大小
3討論
慢病毒載體是以人類免疫缺陷型病毒為基礎發展起來的基因治療載體,它對分裂細胞和非分裂細胞均具有感染能力,并可以較長期穩定的表達目的基因,便于基因功能的研究。
Lefty蛋白是TGFβ大家族的配體,通過與Nodal以及Nodal的受體結合而抑制Nodal信號轉導[2]。Nodal是早期胚胎干細胞誘導的關鍵信號分子,參與中胚層和內胚層形成、前后體軸的位置確定和左-右體軸特化等關鍵事件[3]。Nodal還可以抑制人類胚胎干細胞的分化,維持人類胚胎干細胞多潛能干性[4]。
有報道指出,Nodal在多種惡性腫瘤(惡性黑色素瘤、乳腺癌、腦膠質瘤等)中能高水平表達,且表達水平與腫瘤浸潤和轉移等密切相關[5-7]。有關惡性黑色素瘤及乳腺癌的研究提示[8]:在轉移性腫瘤細胞中出現Nodal信號通路的抑制因子Lefty缺失,從而導致Nodal的過度表達;胚胎干細胞微環境(含Lefty)將明顯下調Nodal的表達,并抑制腫瘤的發生發展,促進腫瘤細胞凋亡。因此Nodal信號通路在胚胎發育和腫瘤轉移中的差異可能是:胚胎組織可以通過分泌Lefty等蛋白分子對發育過程進行調控,而腫瘤中出現Lefty等缺失。這提示腫瘤浸潤和轉移的發生與腫瘤組織缺乏類似胚胎發育時精細有效的調控機制密切相關。基于上述分析及前期研究結果,我們提出假設:胃癌細胞中Lefty等Nodal信號通路的抑制因子缺失,是導致Nodal信號失控的重要原因。
故我們成功進行了LeftyA基因重組慢病毒的構建與包裝,為下一步的功能學實驗打下基礎。
參考文獻
[1]Forman D,Burley VJ.Gastric cancer:global pattern of the disease and an overview of environmental risk factors〔J〕.Best Pract Res Clin Gastroenterol,2006,20(4):633-649.
[2]Cheng SK,Olale F,Brivanlou AH,et al.Lefty blocks a subset of TGFbeta signals by antagonizing EGF-CFC coreceptors〔J〕.PLoS Biol,2004,2(2):E30.
[3]Shen MM.Nodal signaling:developmental roles and regulation〔J〕.Development,2007,134(6):1023-1034.
[4]James D,Levine AJ,Besser D,et al.TGFbeta/activin/nodal signaling is necessary for the maintenance of pluripotency in human embryonic stem cells〔J〕.Development,2005,132(6):1273-1282.
[5]Topczewska JM,Pestovit LM,Margaxyan NV,et al.Embryonic and tumorigenic pathways converge via Nodal signaling:Role in melanoma aggressiveness〔J〕.Nat Med,2006,12(8):925-932.
[6]Strizzi L,Postovit LM,Margaryan NV,et al.Emerging roles of nodal and cripto-I:from embryogenesis to breast cancer progression〔J〕.Breast Disease,2008,29:91-103.
[7]Lee CC,Jan HJ,Lai JH,et al.Nodal promotes growth and invasion in human gliomas〔J〕.Oncogene,2010,29(21):3110-3123.
[8]Pestovit LM,Margaryan NV,Seftor EA,et al.Human embryonic stem cell microenvironment suppresses the tumorigenic phenotype of aggressive cancer cells〔J〕.Proc Natl Acad Sci,2008,105(11):4329-4334.
(編輯:吳小紅)
