浦東白豬Nramp1基因第6內含子獨特突變位點分析
2015-12-27 02:36:17周文兵俞向前葉承榮朱建國
豬業科學 2015年7期
關鍵詞:研究
周文兵,張 艷,俞向前,葉承榮,張 峻,朱建國
(1.上海交通大學農業與生物學院,上海市獸醫生物技術重點實驗室,上海 200240;2.上海市浦東新區動物疫病預防控制中心,上海 201299;3.閔行區動物疫病預防控制中心,上海 201109)
浦東白豬Nramp1基因第6內含子獨特突變位點分析
周文兵1,張 艷1,俞向前2,葉承榮2,張 峻3,朱建國2
(1.上海交通大學農業與生物學院,上海市獸醫生物技術重點實驗室,上海 200240;2.上海市浦東新區動物疫病預防控制中心,上海 201299;3.閔行區動物疫病預防控制中心,上海 201109)
Nramp1是學術界近年研究的豬重要抗病基因之一,其第6內含子作為Nramp1的分型重點片段,國內外的諸多豬種均有清晰的測序分析結果。本研究通過提取浦東白豬血液組織DNA,使用PCR技術分離和克隆Nramp1基因的第6內含子基因片段,測序后與以往文獻中的其他豬種進行比較。 結果發現,浦東白豬在該段基因中有3個特異的突變點普遍存在,分別是259 bp的A→G突變,331 bp的G→A突變和29 bp后1~2個T堿基的插入。這些突變的發現可以為浦東白豬的育種和Nramp1的抗病性研究提供遺傳數據。
浦東白豬;Nramp1基因;第6內含子;PCR;基因突變
浦東白豬作為上海市特有的地方豬種,具有繁殖率高,性成熟早,閹割后,安靜少動,適合圈飼,肉質鮮美等特點[1],是我國寶貴的地方豬種資源中獨具特色的一個品種。隨著我國養豬業的快速發展,為了防治傳染性疾病所導致的藥物殘留問題日益嚴重。這個問題既影響人們群眾的健康也增加了生產成本。養殖不同品種具有特異性抗病能力的豬種可以有效地降低傳染病在不同豬群間的傳播能力,最終將會降低豬的養殖成本和提高豬肉產品的質量。Nramp(natural resistance associated macrophage protein)基因即天然抗性相關巨噬蛋白基因家族。該家族基因的初次發現是在20世紀70年代,Bradley[2]等人在小鼠上發現了一個導致杜氏利什曼蟲(Leishmania donovani)易感性的等位基因,被命名為Lsh。后來其他科學家[3-4]又先后在小鼠身上找到了對鼠傷寒沙門氏菌(Salmonella typhimurium)和牛結核分枝桿菌(Mycobacterium bovis)易感的等位基因,分別被命名為Ity和Bcg。接著在1982年,這3個等位基因被開始證明在1號染色體上的同一位點[5-6]。借助基因技術的發展,在20世紀90年代Nramp最終被發現和命名[7]后,Lsh/Ity/Bcg均被歸入該家族。Nramp1基因也于次年第1個被完整克隆并定位[8]。Nramp1基因在網狀內皮細胞器官的巨噬細胞(血液外周白細胞、脾臟、肺等)中表達,和動物體對多種胞內病原微生物的抵抗力相關[9]。由于Nramp1基因較為保守,成為近年受研究較為普遍的抗病基因之一。豬的Nramp1基因,已經在國內的諸多豬種上得到研究,而且研究的方向主要集中在第6內含子的Nde I位點分型,國內地方豬種中僅有貴州的香豬、糯谷豬和宗地花豬[10]以及黑龍江的民豬[11]被進行過Nramp1基因第6內含子的基因測序研究。
本研究采用PCR擴增片段后基因測序的研究方法,對浦東白豬的Nramp1基因第6內含子的核苷酸序列進行研究分析,尋找在該片段浦東白豬相對其他豬種的遺傳特異性。本研究的結果將為未來浦東白豬抵抗胞內病原微生物的能力研究提供遺傳學理論基礎,對Nramp1基因的抗病能力研究也會產生積極的意義。
1??材料和方法
1.1 實驗材料
實驗中采用從上海浦匯浦東白豬繁育有限公司豬場采集的浦東白豬血液樣本,使用購自北京百泰克生物技術有限公司的全血基因組DNA快速提取試劑盒(溶液型),貨號:DP1101,提取DNA樣本,用DNA溶解液水化后保存于-20 ℃。
1.2 引物和試劑
參照文獻[12]中的引物設計,引物由生工生物工程(上海)股份有限公司合成。引物設計如下:
F: 5’ GCCAGCTTCCACAGTCT CCAG 3’
R: 5’ GGGGGTACAAAGGGGAA GAAG 3’
引物用去離子水配置成100μM的貯存液,預期擴增片段為483 bp。
1.3 全血基因組DNA快速提取
參照試劑盒說明書進行,簡述如下:
1)將血液加入紅細胞裂解液中,使血液中的紅細胞完全裂解;
2)離心得到白細胞團,利用細胞核裂解液釋放出白細胞的基因組DNA;
3)加入蛋白質沉淀液,離心去除蛋白質雜質;
4)異丙醇沉淀DNA;
5)用70%乙醇漂洗數次;
6)晾干后加入DNA溶解液水化。
1.4 PCR擴增和克隆測序
PCR反應總體系25 μl,組分為:Mix,12.5 μl; Dna模板,3 μl;上下游引物,各1 μl;去離子水,7.5 μl。
PCR運行程序:94 ℃預變性3 min;94 ℃ 30 S,64 ℃ 45 S,72 ℃45 S,30個循環;最后72 ℃延伸5 min。
PCR產物用1%的瓊脂凝膠電泳檢測,目的條帶純化回收,將回收的產物經E.coliDH5α Competent細胞質粒轉化后送鉑尚生物技術(上海)有限公司測序。PCR產物電泳,純化回收和質粒轉化中的主要試劑均購自Takara公司。
2??結果
2.1 PCR擴增
PCR產物經1%瓊脂糖凝膠電泳檢測,結果顯示擴增出的產物均為483 bp左右,與預期片段相符。
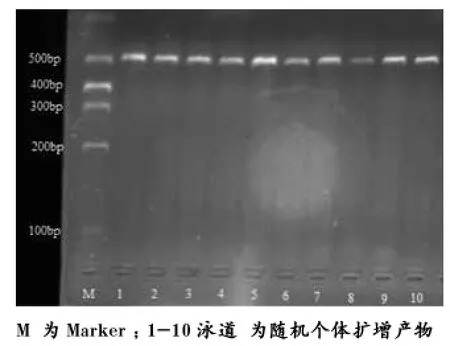
圖1 浦東白豬Nramp1基因第6內含子PCR擴增產物
2.2 擴增結果片段基因測序
選取10個條帶清晰的目的條帶片段,從瓊脂凝膠中回收,經質粒轉化后進行全序列基因測序。基因測序的結果與Gen Bank Nramp1(登陸號:AY368473)的基因序列進行比對,結果發現3個與 AY368473 不同的突變點。突變結果剪輯如圖2、3、4所示,其中AY368473.1為從Genbank選取的基因序列,1-10為選取的PCR擴增產物基因序列。浦東白豬的Nramp1基因第6內含子在331位點出現G→A突變;在259位點出現A→G突變;29位點之后插入了1~2個T堿基。
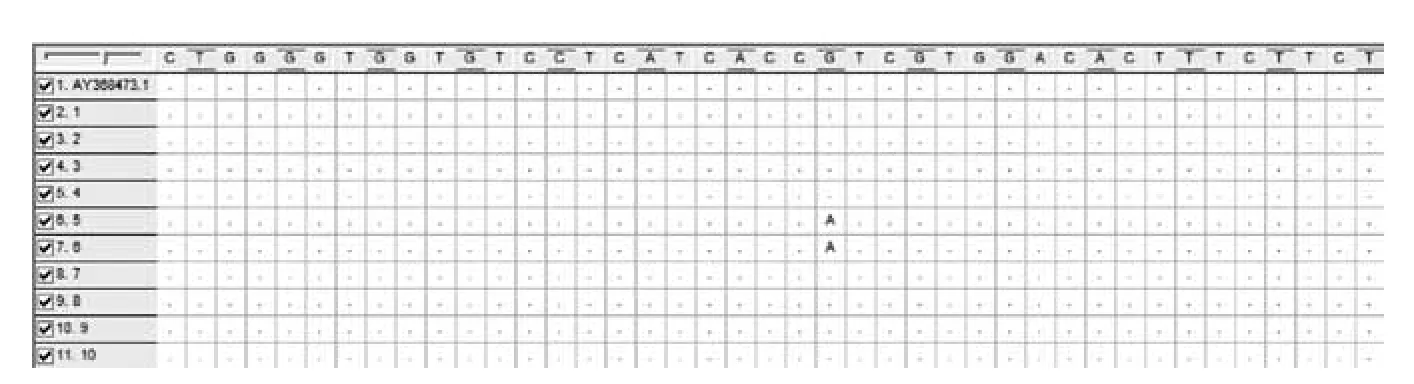
圖2 Nramp1第6內含子基因序列比較結果1
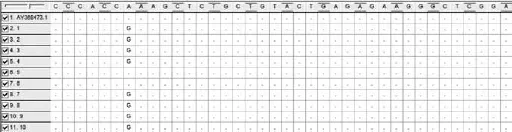
圖3 Nramp1第6內含子基因序列比較結果2

圖4 Nramp1第6內含子基因序列比較結果3
2??討論
近年國內外豬種的Nramp1基因多態性的研究大多集中在第6內含子,尤其圍繞在111 bp位點的C→A突變。自吳宏梅等[12]首次發現該突變在大白豬和松遼黑豬2個豬種上有相反的抗病能力表現后,楊軍等[13]、趙生國等[14]以及劉艷冬[10]分別在不同的中外豬種發現了該突變,并證實了該突變可導致抗病力的差異表現。Nramp1基因作為抗病基因,的確具有重要的研究價值。可惜該突變位點發生作用的機理,至今尚未有較為確切的合理解釋。我國具有廣大的地域,地理上的分割促使了很多地方豬種的獨立培育。對這些地方豬種的第6內含子基因序列進行更廣泛的基因學研究,或許可以找到更多的可致Nramp1基因抗病性改變的基因突變,這將有助于對該現象的解釋,也將對Nramp1基因的抗病機理研究頗有裨益。
本實驗采用PCR方法分離和克隆浦東白豬的Nramp1基因第6內含子片段,在采用基因測序方法來檢測完該片段后,尋找浦東白豬在該片段中特有的突變位點。結果顯示浦東白豬的該片段與國內外已經得到研究的諸多豬種相比具有很高的品種特異性。研究中發現的3個突變位點,既不同于迄今唯一被發現可以產生抗病力差異的111 bp位點突變,與羅文華[15]從4種中外豬種發現的5個突變位點和李娟娟[16]等人從合作豬發現的10個突變位點也沒有重復。這個結果說明了浦東白豬在Nramp1抗病基因的遺傳獨特性,浦東白豬在抗病基因的研究中將可能具有非常重要的作用。如果未來對浦東白豬繼續進行相關基因抗病性的基因學和免疫學研究,會對抗病豬種的培育產生難以預料的幫助。
近20年來隨著國外豬種的大量引入,國內地方豬種的分布范圍嚴重減小,種群數量急劇下降,部分種群中一些低頻率的基因突變位點可能因此而喪失。特別在一些群體基數異常小的豬群,該現象會更加突出。這一方面將使很多地方豬種的遺傳特異性更加突出,另一方面也可能會導致該豬種群體中特有的部分遺傳特性永遠喪失,成為遺傳資源的重大損失。利用基因技術對現存地方豬種進行廣泛的抗病基因學研究,將能夠為保護地方豬種遺傳資源產生重要的作用。
[1] 朱鴻伯.川沙縣志[M].上海:上海人民出版社,1990:199.
[2] Bradley D J. Letter:Genetic control of natural resistance to Leishmania donovani[J]. Nature,1974,250(464):353-354.
[3] Plant J, Glynn A A. Genetics of resistance to infection with Salmonella typhimurium in mice[J]. J Infect Dis,1976 ,133(1):72-78.
[4] P Gros, E Skamene, A Forget. Genetic control of natural resistance to Mycobacterium bovis (BCG) in mice. The Journal of ImmunologyDecember 1, 1981,127(6): 2417-2421.
[5] Plant JE, Blackwell J M.O’Brien AD, Bradley DJ and Glynn AA (1982) Are the Lsh and Ity disease resistance genes at one locus on mouse chromosome 1?[J]. Nature (Lond), 297:510-511.
[6] Skamene E, Gros P, Forget A, et al. Charles C and Taylor BA (1982) Genetic regulation of resistance to intracellular pathogens[J]. Nature (Lond) ,297:506-510.
[7] Vidal S M, Malo D, Vogan K. Skamene E and Gros P (1993) Natural resistance to infection with intracellular parasites: Isolation of a candidate for Bcg[J].Cell,73:469-485.
[8] Cellier M, Govoni G, Vidal S,et al. Schurr E and Gros P (1994) Human natural resistance-associated macrophage protein: cDNA cloning,chromosomal mapping, genomic organization, and tissue-specific expression. J Exp Med 180:1741-1752.
[9] Jenefer M,Blackwell. Structure and function of the natural-resistanceassociated macrophage protein (Nramp1), a candidate protein for infectious and autoimmune disease susceptibility. Molecular Medicine Today Volume 2, Issue 5, May 1996, Pages 205-211.
[10] 劉艷冬.香豬NRAMP1 基因多態性及與仔豬腹瀉相關性研究[D].貴陽:貴州大學, 2009.
[11] 顧永娟.豬NRAMP1基因與仔豬腹瀉及其生產性狀關系的研究 [D]. 哈爾濱:東北農業大學,2006.
[12] 吳宏梅,王立賢,程篤學,等.豬Nramp1 基因多態性與免疫功能的相關性[J].中國農業科學,2008,41(1):215-220.
[13] 楊軍,陳紅頌,丁山河,等.3個外種豬Nramp1基因多態性研究[J].長江大學學報:自然科學版,農學卷,2009,6(4):40-43.
[14] 趙 生國,趙海,潘虹,等.4豬種Nramp1基因第6內含子多態性研究[J].中獸醫醫藥雜志,2012,31(5):34-36.
[15] 羅文華.豬Nramp1基因內含子測序及Nramp1和FUT1基因多態性研究[D].廣州:華南農業大學.2004.
[16] 李娟娟,馬小軍,王立賢,等.合作豬Nramp1基因第六內含子多態性分析[J].甘肅農業大學學報,2010,45(2):10-13.
2015-06-04)
上海市種業發展項目地方畜禽遺傳資源保護與開發關鍵技術研究和示范及上海市生豬現代農業產業技術體系項目
周文兵,男,江蘇省宿遷市人,碩士研究生,研究方向為預防獸醫學
朱建國,email: zhu_jg@sjtu.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
