Lactobacillus casei Zhang傳1 000-2 000代過程中的實驗室進化特性*
2015-12-25 01:59:08馬瑞芬惠文彥白梅張文羿孫天松
食品與發酵工業 2015年4期
關鍵詞:研究
馬瑞芬,惠文彥,白梅,張文羿,孫天松
(內蒙古農業大學乳品生物技術與工程教育部重點實驗室,內蒙古呼和浩特,010018)
益生菌是指通過攝入適當量從而對宿主產生有益作用的活性微生物,其通常可來源于發酵食品、有益共生物和環境[1]。這些微生物和宿主間的相互作用機理多種多樣,例如:影響共生菌群的結構組成和功能、改變宿主上皮細胞結構或者免疫應答[2]。近些年的研究成果顯示,益生菌的攝入在特定條件下可改變特定人群的腸道菌群結構,包括過敏性皮炎、結腸炎和腸應激綜合征,但一直缺乏有力的支撐證據[3-5]。
微生物的實驗室進化研究(adaptive laboratory evolution,ALE)是在人為因素的作用下通過控制環境來研究微生物的進化。早在1983年,細菌學家Russell就列舉了在進化生物學領域研究中細菌的一些“明顯生長優勢”,其中就提出嚴格控制細菌生長的變化的環境,并在適當時間和空間內為其更換新鮮培養基來研究細菌[6]。直到20世紀末實驗室進化理論才以一個獨立的學科問世[7]。最初的實驗室進化是對恒化器中適應性進行準確測量以揭示自然突變與工程突變之間的微妙變化與聯系[8]以及代謝與適應性間的關系[9]。之后的研究是長期連續傳代實驗。伴隨著分子技術的進步,實驗室進化已經延伸到基因水平的研究。通過全基因組序列的測定,可進一步揭示進化的動力學問題[10]。
Lactobacillus casei Zhang(L.casei Zhang)是從內蒙古地區蒙古族家庭制作的16份酸馬奶樣品中分離得到的1株益生乳桿菌[11]。具有以下特點:優良的益生特性,包括酸和膽鹽耐受性[12]、降膽固醇、抗氧化、抗菌、免疫調節以及調節腸道菌群結構等[13-16]。本文在已有工作基礎之上,檢測了L.casei Zhang在普通MRS培養基中連續傳代1 000~2 000代期間3個譜系的進化特性變化情況。
1 材料與方法
1.1 材料
1.1.1 菌株來源
益生菌L.casei Zhang培養1 000代后的衍生菌株是由內蒙古農業大學“乳品生物技術與工程”教育部重點實驗室乳酸菌菌種庫(LABCC)提供,LABCC編號IMAU10048。
1.1.2 試劑與儀器
試劑:法國梅里埃合成無糖MRS培養基,紐西蘭超特級脫脂乳粉,葡萄糖、HCl、NaOH、NaCl、瓊脂粉等常用試劑均為國產分析純試劑。
儀器:潔凈工作臺(SW-CJ-1F型),蘇凈集團蘇州安泰空氣技術有限公司;恒溫培養箱(LRH-500F型),上海一恒科學儀器有限公司;臺式高速冷凍離心機(Eppendorf,5810R 型),德國;pH 計(SJ-3F 型),上海精密科學儀器有限公司雷磁儀器廠;超純水系統(MUL-9000,昆山總馨企業有限公司;紫外可見分光光度計(UV-1700 PharmaSpec型,SHIMADZU);全自動高壓滅菌鍋(MLS-3751L型,PANASONIC)等。
1.2 實驗方法
1.2.1 L.casei Zhang連續傳代培養及培養代數計算
將1 000代益生菌L.casei Zhang三株平行菌株L.casei Zhang-1、L.casei Zhang-2、L.casei Zhang-3 按體積分數1%接種量進行長期連續傳代培養至2 000代。在每個傳代周期(24 h)內,菌體菌數約增長100倍,生長代數約為6.64 代[17]。
1.2.2 L.casei Zhang細胞形態觀察
在菌株L.casei Zhang連續傳代過程中,培養至2000代時對菌株進行革蘭氏染色、油鏡觀察、照相,觀察培養周期末菌株細胞形態。
1.2.3 L.casei Zhang培養周期末活菌數變化
在菌株傳代過程中,每培養200代測定各個菌株在培養周期末(24 h)的活菌數。樣品經倍比稀釋到合適梯度后,采用MRS固體培養基平皿傾注法,37℃厭氧培養72 h后計菌落總數,選取菌落數在30~300之間的平板作為活菌總數測定的指標。
1.2.4 L.casei Zhang培養周期末濁度變化
在菌株傳代過程中,每培養200代測定連續3天的培養周期末的培養液濁度。傳代培養液用未接菌培養基適當稀釋,采用比濁法于600 nm處測定其濁度。
1.2.5 L.casei Zhang產酸活力測定
在菌株傳代過程中,每培養200代測定菌株的產酸活力。各菌株以1×107/mL接種于10 mL滅菌脫脂乳中,37℃恒溫培養24 h,取出后立即放入冰浴,每樣加30 mL蒸餾水稀釋后,以酚酞作指示劑,以0.1 mol/L的NaOH滴定至微紅色30 s不退色為止,記錄消耗NaOH的體積以計算出樣品中的乳酸濃度,再換算為活力單位。
發酵活力:以每107個菌體細胞在此條件下發酵1 mL脫脂乳產生1 μg乳酸計為1個活力單位(U)[18]。
U=0.1 V×M
式中:U為活力單位;V為消耗NaOH的體積;M為乳酸的摩爾質量。
1.2.6 數據處理
采用DPS軟件對數據進行方差分析;采用Origin8.6繪圖軟件進行繪圖。各項指標均做3個平行。
2 結果與分析
2.1 L.casei Zhang細胞形態觀察
圖1所示為L.casei Zhang分別在1 000代和2 000代時顯微鏡照相的照片。由圖1可知,L.casei Zhang 1 000代的3個譜系的細胞均為短桿狀、成對或呈鏈桿狀排列。與1 000代3個譜系細胞形態相比,在MRS培養基中連續傳代至2 000代,3個譜系的細胞形態與之前相同,細胞體積也沒有發生顯著變化。這說明,L.casei Zhang細胞未發生明顯的變異情況。當然,這一結論只能作為評價該譜系群體進化規律的部分參考,關于其細胞內部成分組成及其比例是否發生變化仍然需要更進一步的研究。

圖1 L.casei Zhang傳1 000~2 000代期間細胞形態對比Fig.1 Cell appearance of Lactobacillus casei Zhang during subculturing for 1 000~2 000 generations
2.2 L.casei Zhang生長周期末活菌數
L.casei Zhang在連續培養1 000~2 000代期間,不同代數3個譜系所形成的群體在培養周期末的活菌數變化情況見圖2。
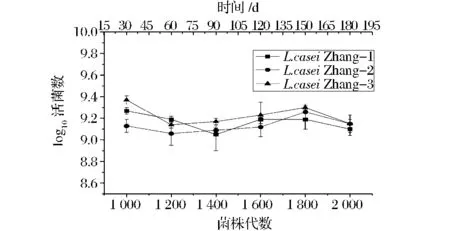
圖2 L.casei Zhang在不同培養代數的活菌數變化Fig.2 Viable counts of L.casei Zhang of different generations
由圖2可以看出,在MRS培養基中,3個譜系活菌數在傳代過程中變化不明顯;縱向比較3個平行培養的菌落發現,在最初開始培養時活菌數就有差異,但這樣的差異并沒有隨培養代數的增加而持續。總體來看,L.casei Zhang在傳代1 000~2 000代期間,3個譜系間活菌數的進化趨勢基本平行,菌株的平均適應性也相對穩定。側面反映了該菌株具有穩定的表型特性。
2.3 L.casei Zhang周期末濁度
在連續培養1 000~2 000代過程中,L.casei Zhang在培養周期末的OD600值變化情況見圖3。由圖3可以看出L.casei Zhang三個譜系的OD600值在1 400代之前均呈現增長的趨勢,到1 600代時達到最低點又整體下降,之后又有所增加。通過對比活菌數發現菌株在1 600代時適應性與之前相比無明顯差異,因此推斷3個譜系在連續進化1 000~2 000代期間OD600值產生的不規律的變化可能是最初接菌時產生的誤差所致。3個譜系之間周期末濁度沒有發生明顯變異.且處于平行進化的趨勢。

圖3 L.casei Zhang在不同培養代數的濁度變化Fig.3 The OD600value of L.casei Zhang of different generations
2.4 L.casei Zhang產酸活力變化
實驗研究了連續傳代培養1 000~2 000代過程中,每200代測定菌株的產酸活力,測定結果如表1所示。由表1可以看出,菌株在1 400代時3個譜系的產酸活力均有微小下降,但變化不顯著。
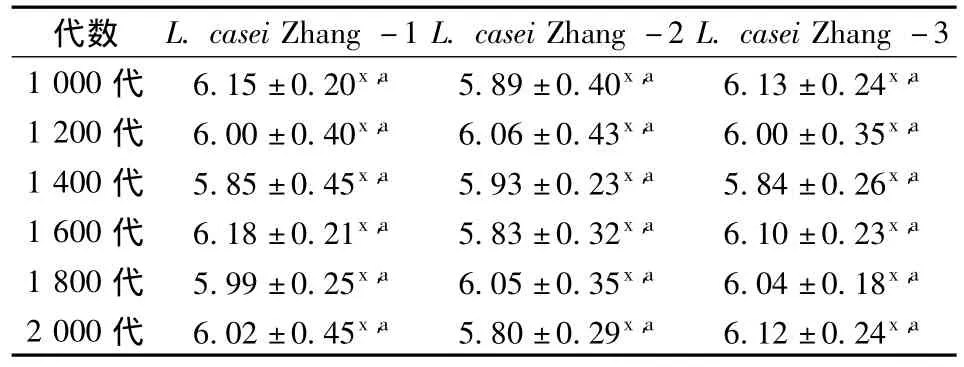
表1 L.casei Zhang在不同培養代數的菌體產酸活力變化(U)Table 1 Acid-producing ability of L.casei Zhang of different generations
3 討論
近年來,關于微生物實驗室進化特性的研究越來越多,早期有對單細胞藻類的研究[21],還有利用分子生物學技術對酵母進化特性進行研究[22]。一般來說,在微生物進化過程中,必然的會發生菌株變異,從而導致菌株部分特性的改變,這也是進化的必然趨勢。例如,Merry等人對土壤細菌實驗室進化1 000代進化特性的觀察發現群體細胞長度有增加的現象[23]。此外,對大腸桿菌連續傳代40 000~50 000代進化特性研究發現中,其平均適應性呈現遞增趨勢,其中增加最多達3%[24]。除了菌株適應性增長之外,還有一些顯著的表性特征的改變,如細胞的體積隨著傳代的進行呈現增加趨勢。或者是菌落形態比原始菌株形狀更圓等[25]。這些表觀特性的變化可能是細菌為了更好地適應環境而做出的“自我改變”。
與其他人研究結果不同的是,在對益生菌L.casei Zhang在普通MRS培養基中傳代1 000~2 000代期間實驗室進化特性的研究發現,該菌株的表觀特性包括細胞形態、活菌數、濁度和菌株活力等指標均未發生顯著變異,而且保持著幾乎平行的進化趨勢。由此可以側面反映出L.casei Zhang是一株相對穩定的益生菌菌株。但是,由于實驗所測指標仍然存在一定局限性,該結果并不能夠全面評價該菌株的進化特性。
4 結論
研究結果表明,益生菌L.casei Zhang在正常培養條件下3個譜系間表性特征進化趨勢相對平行,然而,這些進化特性的結果仍存在局限性。一方面,細菌由于世代時間較短,繁殖迅速,即使出現某些突變,也可能會在短期內消失,這就給研究帶來一定困難,另一方面,本實驗傳代時間較短,而菌株的一些特性需要突變不斷積累才會表現出來,這也需要更充足的數據來驗證。總體來說,這些數據雖不能全面說明L.casei Zhang在生理學特性方面是否發生了微妙的變化,但該實驗結果仍然可以作為評價該菌株進化特性的有力依據,也可為該菌株的產業化生產提供很好的保障。
[1] Sanders M E,Guarner F,Guerrant R,et al.An update on the use and investigation of probiotics in health and disease[J].Gut,2013,62(5):787-796.
[2] Bengmark S.Gut microbiota,immune development and function[J].Pharmacological Research,2013,69(1):87-113.
[3] Arora T,Singh S,Sharma R K.Probiotics:Interaction with gut microbiome and antiobesity potential[J].Nutrition,2013,29(4):591-596.
[4] Barraud D,Bollaert P E,Gibot S.Impact of the administration of probiotics on mortality in critically ill adult patients probiotics in the ICUA Meta-analysis of randomized controlled trials[J].CHEST Journal,2013,143(3):646-655.
[5] De Greef E,Vandenplas Y,Hauser B,et al.Probiotics and IBD[J].Acta Gastro-enterologica Belgica,2013,76(1):15-19.
[6] Russell H L.Bacteriology in its general relations[J].American Naturalist,1893,27(322):847-859.
[7] Elena S F,Lenski R E.Evolution experiments with microorganisms:the dynamics and genetic bases of adaptation[J].Nature Reviews Genetics,2003,4(6):457-469.
[8] Dykhuizen D E,Hartl D L.Selection in chemostats[J].Microbiological Reviews,1983,47(2):150.
[9] Dykhuizen D E,Dean A M,Hartl D L.Metabolic flux and fitness[J].Genetics,1987,115(1):25-31.
[10] Hegreness M,Kishony R.Analysis of genetic systems using experimental evolution and whole-genome sequencing[J].Genome Biol,2007,8(1):201.
[11] 孟和畢力格,烏日娜,王立平,等.不同地區酸馬奶中乳桿菌的分離及其生物學特性的研究[J].中國乳品工業,2004,32(11):6.
[12] 張和平,孟和畢力格,王俊國,等.分離自內蒙古傳統發酵酸馬奶中L.casei Zhang潛在益生特性的研究[J].中國乳品工業,2006,34(4):4-10
[13] WU R,WANG W,YU D,et al.Proteomics analysis of Lactobacillus casei Zhang,a new probiotic bacterium isolated from traditional home-made koumiss in Inner Mongolia of China[J].Molecular& Cellular Proteomics,2009,8(10):2 321-2 338.
[14] WANG J C,ZHANG W Y,ZHONG Z,et al.Transcriptome analysis of probiotic Lactobacillus casei Zhang during fermentation in soymilk[J].Journal of Industrial Microbiology& Biotechnology,2012,39(1):191-206.
[15] WANG J,WU R,ZHANG W,et al.Proteomic comparison of the probiotic bacterium Lactobacillus casei Zhang cultivated in milk and soy milk[J].Journal of Dairy Science,2013,96(9):5 603-5 624.
[16] Lenski R E,Rose M R,Simpson S C,et al.Long-term experimental evolution in Escherichia coli.I.adaptation and divergence during 2,000 generations[J].American Naturalist,1991,138(6):1 315-1 341.
[17] WANG J,GUO Z,ZHANG Q,et al.Fermentation characteristics and transit tolerance of probiotic Lactobacillus casei Zhang in soymilk and bovine milk during storage[J].Journal of Dairy Science,2009,92(6):2 468-2 476.
[18] 李妍.益生菌Lactobacillus casei Zhang高密度培養技術及發酵過程中關鍵酶基因表達變化的研究[D].呼和浩特:內蒙古農業大學,2008.
[19] Colegrave N,Kaltz O,Bell G.The ecology and genetics of fitness in Chlamydomonas.VIII.The dynamics of adaptation to novel environments after a single episode of sex[J].Evolution,2002,56(1):14-21.
[20] Zeyl C.Experimental evolution with yeast[J].FEMS Yeast Research,2006,6(5):685-691.
[21] Riley M S,Cooper V S,Lenski R E,et al.Rapid phenotypic change and diversification of a soil bacterium during 1000 generations of experimental evolution[J].Microbiology,2001,147(4):995-1 006.
[22] Wiser M J,Ribeck N,Lenski R E.Long-term dynamics of adaptation in asexual populations[J].Science,2013,342(6164):1 364-1 367.
[23] Lenski R E,Mongold J A.Cell size,shape,and fitness in evolving populations of bacteria[C].Scaling in Biology.Oxford University Press,2000:221-235.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
