釀酒酵母胞外蛋白在衰老過程中的變化
2015-12-25 01:58:10杜維樸永哲周彥品王小瑜趙長新
食品與發酵工業 2015年8期
杜維,樸永哲,周彥品,王小瑜,趙長新
1(大連工業大學,遼寧 大連,116034)2(大連民族大學,遼寧 大連,116600)
分泌蛋白(secretory protein)是指細胞或組織分泌的蛋白,同胞內蛋白一樣在參與細胞增殖、分化以及細胞間的信號轉導等重要生命過程中發揮作用[1]。深入了解分泌蛋白有助于弄清不同生長發育時期、不同培養狀態下的細胞狀態信息,細胞異常時的病理病因以及認識整個生命過程具有重要的意義[2]。Zwickl等[3]提供了一種代謝標簽摻入的方法研究胞外蛋白,方法對HepG細胞和人肝組織切片進行的研究結果證實對預防和治療肝癌其有廣泛的臨床應用價值。相比于原核細胞,真核細胞具有大量結構復雜的隔性間隔,為運輸與分泌帶來比較大的困難。公認的蛋白分泌途徑主要有2種[4],一種是常規分泌途徑的信號假說 (signal hypothesis);另一種為非常規的蛋白質分泌途徑,如白介素-1b、纖維原細胞生長因子等都是通過不依賴信號肽的非常規蛋白質分泌途徑分泌的[5]。目前已對蛋白質的轉運與分泌進行了廣泛深入的研究,但至今對分泌途徑種類未有一個確切的定論。
衰老是自然界生物體的普遍規律,是作為生命載體-細胞的最終歸宿。衰老過程伴隨著復制生殖能力退化,細胞代謝不可逆減緩,細胞周期停滯,細胞生理及形態學同時也發生改變等。衰老包括復制性衰老和時序型衰老[6],前者是指能復制出子代的個數,這種衰老與遺傳物質的損傷有關;時序型衰老是指在特定培養條件下菌體存活的時間。目前衰老體系已經作為一種模式應用到絲狀細菌(Podospora anserine)、釀酒酵母(Saccharomyces cerevisiae)、線蟲(Caenorhabditis elegans)、果蠅(Drosophila melanogaster)和小鼠(Musmusculus)等模式生物中來研究復雜的衰老現象[7]。本文以釀酒酵母為載體,探究在釀酒酵母時序型衰老的狀態下胞外蛋白與胞內酶活性之間的關系,探討衰老狀態下胞外蛋白來源的變化情況。
1 材料和方法
1.1 材料
1.1.1 菌種
Saccharomyces cerevisiae FFC 2144,大連工業大學菌種保藏中心。
1.1.2 儀器
湘儀高速冷凍離心機H2050R-1;MAPADA分光光度計UV6100;北京六一雙向電泳槽DYCZ-26B;北京博醫康冷凍干燥機VFD2000;上海智城恒溫搖床ZWYR-2101C;大龍三維脫色搖床 SK-D3309-Pro;Congent超濾儀;超聲波細胞破碎儀XO-1000;Thermo Scientific臺式四極桿-軌道阱高分辨質譜儀等。
1.1.3 藥品
General Electric Company載體兩性電解質pH 4-6.5,載體兩性電解質 pH 3-10。過硫酸銨、丙酮、苯酚、尿素(科密歐,優級純),愈創木酚,TCA,甲醇(科密歐,色譜純);ASB-14硫脲、CHAPS、色氨酸、組氨酸、YNB、精氨酸、丙烯酰胺、SDS(阿拉丁)。
1.1.4 培養基
采用改良無蛋白質的全合成YNB培養基[8],葡萄糖 2%,(NH4)2SO41%,YNB 0.67%,DL-蘋果酸0.6%,酒石酸0.2%,色氨酸0.5‰,組氨酸0.5‰,精氨酸0.5‰。
1.2 實驗方法
1.2.1 菌落計數法測定菌體濃度
將菌液用含有0.05%瓊脂的無菌生理鹽水稀釋成不同的濃度梯度,取20 μL用九格法涂到YPD平板上。每個平板3個梯度,每個梯度3個平行。涂布后的平板放入28℃下培養24 h后進行菌落計數,并換算出實際活菌濃度。活菌濃度定義為每毫升溶液中能自身分裂產生子代的活菌數,單位CFU/mL。
1.2.2 老化培養
選取細胞兩種壽命中的時序型壽命作為考察條件,在菌體生長到對數期末時轉入到無菌生理鹽水中培養,通過長時間培養使酵母細胞老化,觀察細胞形態和酶活性等情況。
1.2.3 胞內粗酶干粉的提取
將發酵液10 000×g,10 min離心棄掉上清得到酵母菌體,菌體沉淀用預冷PBS清洗3次,將清洗后的菌體放入研缽中液氮研磨。收集研磨后的菌體粉末,凍干保存。
1.2.4 胞外蛋白提取
用凍干/平衡酚-丙酮沉淀法提取胞外蛋白:將過0.22 μm膜后的發酵液直接凍干濃縮至80 mL,加入等體積的Tris-平衡酚(1 mol/L,pH 8.0)冰浴攪拌30 min后離心棄上清,保留酚相及中間層。加入3倍體積的濃度為100 mmol/L的乙酸銨-甲醇后于-20℃過夜沉淀,次日離心后的沉淀先用含有0.1%(w/v)DTT的冷丙酮清洗,再用冷丙酮清洗2次,每次10 min。清洗后的蛋白進行真空干燥除去殘余丙酮,-80℃保存。
1.2.5 胞內酶活性測定
精確稱取0.5 g粗酶干粉溶于10 mL PBS緩沖液中,漩渦振蕩促進溶解后離心,上清液即為酶溶液。整個過程盡量低溫短時間內完成以減少酵母胞內自身酶系的相互作用。測定酶溶液中超氧化物歧化酶(superoxide dismutase,SOD)、過氧化物酶(peroxidase,POD)、蛋白酶的酶活性。在測定SOD、POD酶活性時加入終濃度為0.5 mmol/L的PMSF以抑制蛋白酶對測定結果的影響。其中SOD酶用鄰苯三酚自氧化法[9],將4.6 mL Tris-HCl 2×緩沖液與4.4 mL蒸餾水混合后加入100 μL粗酶液25℃水浴加熱20 min,在紫外波長325 nm處每隔30 s,測3 min內吸光值的變化與鄰苯三酚自氧化速率作對比。酶活力定義為每毫升酶液中SOD抑制率達50%時,所對應的SOD量為1個SOD活力單位(U);用愈創木酚法[10]測定POD酶活性,將0.2 mol/L,pH 6.0的磷酸鹽緩沖液2.9 mL、2.0%的 H2021.0 mL、0.05 mol/L愈創木酚1.0 mL、粗酶液0.1 mL搖勻,在37℃水浴中保溫15 min,立即加入10%的TCA 2 mL終止反應,在470 nm處測量1 min吸光值的變化的平均值。以每分鐘吸光值變化0.01為1個酶活力單位(U)。蛋白酶活性測定采用國標法,酶活力定義為在40℃下,將每分鐘水解酪蛋白產生1 μg酪氨酸定義為1個活力單位。
1.2.6 胞外蛋白的雙向電泳
1.2.6.1 蛋白樣品的溶解及定量
取保存的蛋白質干粉溶于適量的裂解液中(7 mol/L 尿素,2 mol/L 硫脲,2%CHAPS,2%ASB-14,20 μL 3-10兩性電解質,0.1%DTT)4℃過夜并用Bradford法[11]測定蛋白質含量,根據裂解液體積估算蛋白質提取率,并將裂解液稀釋至上樣所需濃度。
1.2.6.2 等電聚焦
IEF 膠條的制作:取540 μL H2O,200 μL 30%丙烯酰胺,0.6 g尿素,4~6載體兩性電解質 4.8 μL,3~10 載體兩性電解質 24 μL,5 μL 10% 過硫酸銨,4 μL TEMED混勻并緩緩注入到玻璃管中(12 cm),室溫聚合。等電聚焦電壓如下:300 V,30 min;500 V,1 h;800 V,1 h;1 600 V,4 h。
1.2.6.3 膠條平衡
首先將等電聚焦后的膠條由玻璃管的一端擠出放入含有0.2 g/mL DTT的平衡液中平衡15 min,然后再置于0.2 g/mL碘乙酰胺中平衡17 min。平衡液:6 mol/L尿素,體積分數為20%甘油,2%SDS,50 mmol/L Tris-HCl(pH 8.8,1.5 mol/L)。
1.2.6.4 SDS-PAGE
將平衡后的膠條轉移到12%濃度的二維膠上進行恒流電泳,參數為20 mA/gel 15 min,40 mA/gel直至溴酚藍移動到膠條的最低端。
1.2.6.5 蛋白染色
Neuhoff考染法[12]
1.2.7 蛋白點鑒定
1.2.7.1 前期酶解
用去尖的槍頭從膠上切下蛋白點后清洗,加入50%乙腈/0.2 mol/L NH4HCO3(pH 8.0)進行脫色,用100%ACN進行脫水,凍干后的膠加入胰蛋白酶溶液進行酶解。
1.2.7.2 質譜樣品制備
向吸漲后的膠塊加入不含胰蛋白酶的50 mmol/L NH4HCO3緩沖液,放置在37℃培養箱反應過夜。次日用掌式離心機離心,取出上清轉移至加入了60%CH3CN/5%TFA的新管中;在沉淀中加入超純水,孵育10 min,離心移出上清與前一管上清液混合;再向沉淀中加入60%ACN/0.1%TFA超聲15 min,吸出溶液,并入前次上清,反復抽提2次,將所得上清用真空離心濃縮儀干燥5 min,放入-80℃冰箱中過夜。次日凍干后-80℃保存。
1.2.7.3 質譜分析
色譜條件:富集柱:C18(100 μm ×2 cm),分析柱C18(75 μm ×15 cm,2 μm),流動相 a:98%H2O+2%CH3CN+0.1%TFA,流動相 b:20%H2O+80%CH3CN+0.1%TFA,柱溫 35 ℃,流速 0.25 μL/min。質譜條件:噴霧電壓2.5 kV,離子源溫度300℃,掃描范圍:m/z=300~2000。利用Xcalibue軟件和NCBI等相關數據庫搜索比對。
2 結果與討論
2.1 衰老過程中培養基內活細胞數與胞外蛋白測定結果
將處于對數末期的酵母細胞轉入無菌生理鹽水培養,每2天用菌落計數法測定活細胞數量以及胞外蛋白含量,結果如圖1所示。
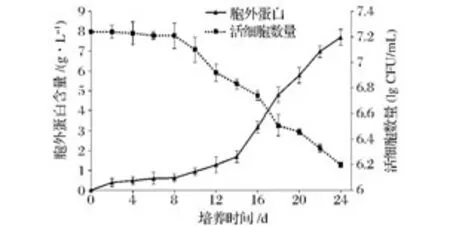
圖1 衰老過程中胞外蛋白與活細胞數量關系圖Fig.1 The number of viable cells and extracellular protein content in the culture
圖1可知在無菌生理鹽水中培養的酵母細胞在8 d內細胞數量基本維持穩定,測定結果來看細胞數目峰值為107.25個/mL,到第8天時細胞數目為107.21個/mL。可復制出子代的活性細胞占峰值的92%以上,8%細胞中包括休眠細胞、死亡裂解細胞以及誤差等。圖1同時顯示8 d內胞外蛋白含量沒有明顯升高,通過以上的數據進行下一步實驗方案的確定,盡量將細胞裂解對胞外蛋白組成的干擾降到最低,故第8天為胞外蛋白的最佳提取時間。從第8天開始活細胞數量顯著下降,胞外蛋白含量明顯增加。24天時細胞數量下降約1個單位左右,胞外蛋白濃度達到7.2 g/L。由此可知在無菌生理鹽水培養4 d內細胞由于處在無外源C源的脅迫下,酵母產生了一種應答機制[13]。這種應答機制將利用胞內儲存的能量合成并向外環境分泌蛋白酶和信號肽等胞外蛋白。隨著培養的繼續細胞內能量耗盡,一部分細胞出現老化自溶,一部分細胞進入休眠狀態,這兩類細胞在菌落計數中不會被體現。有活性的菌體數每下降0.7個單位說明有一半的細胞老化嚴重不能在充足營養下短時間內復制出子代,在此過程中胞外蛋白含量急速上升,說明有小部分細胞自溶亦或是酵母胞內蛋白在向外排出。
2.2 衰老過程中胞內酶活性測定結果
根據圖1得到的數據酵母在無營養培養基中老化衰老,在第8天開始逐漸轉為休眠狀態亦或是老化凋亡。故8 d內酵母形態及數目基本保持完整,有利于測定胞內酶系變化。測定9 d內酵母老化過程中胞內SOD酶、POD酶、蛋白酶的變化,結果如圖2所示。
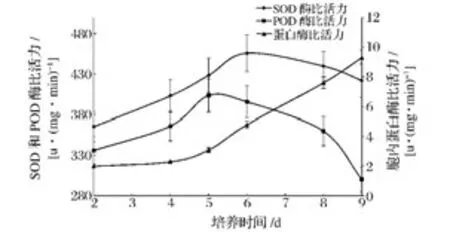
圖2 衰老培養過程中SOD酶、POD酶、蛋白酶活性Fig.2 The activities of SOD,POD and protease during long term fermentation in saline
SOD、POD酶是細胞抗氧化酶系中的關鍵酶,負責將對細胞有害的氧自由基(super oxygen anion,O2-)轉化成低害的H2O2,并最終轉化成無害的H2O,其活性的變化與細胞的氧化程度密切相關。故SOD、POD酶活性相對高低可以作為細胞老化程度的一個依據[14]。從圖上看,在持續饑餓脅迫下 SOD酶、POD酶的活性都呈先升高后降低的趨勢,在5~6天時達到峰值,而后迅速下降。在前5 d的衰老過程中SOD、POD酶活性上升以清除不斷增加的氧自由基,但5 d后由于細胞內能量消耗殆盡導致胞內抗氧化酶活性降低,不能及時清除不斷產生的自由基,自由基的積累會導致細胞組織機能紊亂,進而使細胞穩態失衡、老化加劇[15]。與之不同的是胞內蛋白酶活性一直處于升高狀態,且隨著培養天數的增加蛋白酶活性升高明顯。胞內蛋白酶主要存在于溶酶體中,在細胞內的消化器官、細胞自溶、防御以及對某些物質的利用等方面起作用。5~6天時SOD、POD酶活性的降低與蛋白酶活性的急劇升高相對應。胞外蛋白不斷增加的原因可能是細胞受到氧化性損傷,在老化狀態下細胞內部分修飾錯誤或老化失活的蛋白被降解從而排到細胞外。
2.3 雙向電泳圖譜比較
提取在YNB培養基培養3 d的酵母胞外蛋白以及在無菌生理鹽水中衰老培養8 d的酵母胞外蛋白做雙向電泳,結果如圖3所示。

圖3 酵母胞外蛋白雙向電泳圖譜Fig.3 Two-dimensional gel electrophoresis analysis of extracellular protein
從圖3來看兩張圖譜的特征相一致,在兩張圖的左側都出現并排的縱向條紋,在同一分子質量上都有并排出現的橫向拖尾,且一半以上的蛋白點都可以相互識別。與正常培養時相比,衰老狀態下酵母胞外蛋白表達量大、蛋白點多20% ~30%,在局部放大圖中可以清晰地看到衰老培養比正常的圖多出許多分子質量較低的隨機散亂的蛋白點。這說明在饑餓脅迫下酵母逐漸衰老的過程中,胞外蛋白的組成發生了較大變化。

圖4 酵母胞外蛋白雙向電泳局部放大圖Fig.4 Magnification of two-dimensional gel electrophoresis
2.4 質譜分析結果
質譜分析結果顯示,在9個鑒定結果中檢測到8個胞內蛋白、1個胞外蛋白、1個蛋白為匹配。匹配到的胞外蛋白覆蓋率較高達到87.63%,匹配到的胞內蛋白匹配率較低,基本都在30%以下。質譜結果表明在衰老培養條件下,從圖譜上看散亂的點為胞內蛋白。蛋白點與數據庫匹配率不高,蛋白點多為對比目標蛋白的碎片,而非完整蛋白。說明這些蛋白經過水解后排出到細胞外,這也與胞內蛋白酶活性升高相吻合。初步推論為衰老過程中的酵母細胞將細胞內的某些被破壞或失活的蛋白水解成碎片后排出到細胞外,而對于這些被排出的蛋白是否被進行統一的修飾標記以及這些蛋白的具體排出途徑有待于我們進一步的論證。

表1 質譜分析結果Table 1 The results of MS analysis
3 結論
本文在連續饑餓脅迫下對釀酒酵母進行衰老培養,提取衰老狀態下的釀酒酵母胞外蛋白并用雙向電泳進行圖譜分析。結合胞內SOD酶、POD酶、蛋白酶活性變化,及質譜的分析結果等得出在衰老狀態下SOD、POD酶活性先升高后降低,過氧化物酶活性的降低導致其不能及時清除衰老過程中產生的自由基,胞內氧活性自由基對細胞造成氧化損傷。同時蛋白酶活性的持續升高使胞外蛋白圖譜變化明顯,蛋白酶活性的升高將一部分老化組織降解成碎片,使此類的“垃圾蛋白”最終排出到胞外。
[1] 張海婧,劉輝,呂麗艷,等.分泌蛋白質組研究進展[J].分析化學,2007,35(6):912-918.
[2] 李明,鄧芳,王保龍.分泌蛋白質組學在腫瘤標志物研究中的進展[J].臨床輸血與檢驗,2013,15(4):407-409.
[3] Zwickl H,Traxler E,Staettnrt S,et al.A novel technique to specifically analyze the secretome of cells and tissues[J].Electrophoresis,2005,26(14):2 779-2 785.
[4] Lee M C,Miller E A,Goldberg J,et al.Bi-directional protein transport between the ER and Golgi[J].Annu Rev Cell Dev Biol,2004,20:87-123.
[5] Seelenmeyer C,Wegehingel S,Tews I.Cell surface counter receptors are essential components of the unconventional export machinery of galectin-1[J].Cell Biol,2005,171(2):373-381.
[6] Steinkraus K A,Kaeberlein M,Kennedy B K,et al.Replicative aging in yeast:the means to the end[J].Annu Rev Cell Dev Biol,2008,24:29-54.
[7] 楊婷,張沖,陳清軒.衰老機制研究進展[J].中國生物工程雜志,2005,25(3):6-11.
[8] CHEN Y P,Kirk N,Piper P W.Effects of medium compositionon MF-α1 promoter-directed secretion of a small protease inhibitor in Saccharomyces cerevisiae batch fermentation[J].Biotechnology Letters,1993,15(3):223-228.
[9] 顧含真,陸領倩,袁勤生,等.超氧化物歧化酶兩種測活方法的比較[J].藥物生物技術,2006,13(5):377-379.
[10] 黃智,戴思慧,馬凌珂,等.西瓜種子萌發過氧化物酶活性變化的研究[J].湖南農業科學,2010(9):43-45.
[11] 李海玲,彭書明,李凜,等.4種常用蛋白濃度測定方法的比較[J].中國生化藥物雜志,2008,29(4):277-278.
[12] Neuhoff V,Arold N,Taube D,et al.Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nano-gram sensitivity using Coomassie Brilliant Blue G-250 and R-250[J].Electrophoresis,1988,9(6):255-262.
[13] Bennett J Giardina,Bruce A Stanley,Hui-Ling Chiang.Glucose induces rapid changes in the secretome of Saccharomyces cerevisiae[J].Proteome Science,2014(12):9-18.
[14] 袁方,樸永哲,劉巨濤,等.外源肌醇、柚皮苷及姜黃素對釀酒酵母抗衰老性能影響[J].釀酒科技,2014,241(7):43-47.
[15] Letelier M,Lepe A,Faúndez M,et al.Possible mechanisms underlying copper-induced damage in biological membranes leading to cellular toxicity[J].Chemico-Biological Interactions,2005,151(2):71-82.
