NH4+對(duì)合成氣乙醇發(fā)酵過(guò)程的影響及其酶學(xué)機(jī)理*
2015-12-25 02:00:58張炎達(dá)王風(fēng)芹彭一丁謝慧宋安東
食品與發(fā)酵工業(yè) 2015年12期
張炎達(dá),王風(fēng)芹,彭一丁,謝慧,宋安東
(河南農(nóng)業(yè)大學(xué)生命科學(xué)學(xué)院,農(nóng)業(yè)部農(nóng)業(yè)微生物酶工程重點(diǎn)實(shí)驗(yàn)室,河南鄭州,450002)
全球能源消耗預(yù)計(jì)到2030年將增加57%,化石燃料的大規(guī)模使用同時(shí)還帶來(lái)諸多的能源緊缺、環(huán)境壓力加劇和燃料價(jià)格大幅度上漲等問(wèn)題[1],這就使得各國(guó)政府急切尋求一種可持續(xù)環(huán)保的再生綠色能源,其中生物燃料乙醇得到了世界廣泛關(guān)注[2]。合成氣乙醇發(fā)酵是目前公認(rèn)的一種極具潛力和競(jìng)爭(zhēng)力的生物燃料乙醇生產(chǎn)技術(shù)[3],該技術(shù)具有原料來(lái)源廣泛、生物催化特異性高、能耗低、耐毒性強(qiáng)和H2∶CO的無(wú)限制比率等優(yōu)點(diǎn)[4]。合成氣是由氣化原材料,如農(nóng)林業(yè)廢棄物和生活有機(jī)廢物等各種生物質(zhì)材料、工業(yè)廢氣、次品煤和天然氣等[5-6]經(jīng)氣化裝置在高溫下含有水蒸氣和氧的條件下裂解為富含H2、CO和CO2三種主要成分的混合氣體,另外還包括其他組分,如:NH3、CH4、NOx、N2、C2H2、C2H4、C6H6、C2H6、SO2和焦油等[7],但根據(jù)原料和氣化設(shè)備不同,合成氣的組分和比列略有差異[8-9]。
合成氣厭氧乙醇發(fā)酵微生物目前的種類(lèi)及數(shù)量較少,且乙醇發(fā)酵產(chǎn)量較低,如C.ljungdahlii和C.autoethanogenum等[10],而國(guó)內(nèi)還未有新菌的報(bào)道。Wood-Ljungdahl途徑是該類(lèi)菌體的唯一代謝路徑,且氫化酶(H2ase,hydrogenase)、一氧化碳脫氫酶(CODH,carbon monoxide dehydrogenase)、乙醇脫氫酶(ADH,alcohol dehydrogenase)和乙酸激酶(ACK,acetate kinase)為其四大關(guān)鍵酶,而H2ase為還原力代謝的必需酶[3,7],H2ase活性的高低與 CO 利用效率的大小成正相關(guān)。在合成氣乙醇發(fā)酵中,合成氣雜質(zhì)氣體多對(duì)發(fā)酵產(chǎn)生有害影響,如Asma Ahmed等人[11]發(fā)現(xiàn),微量的焦油和NO能抑制發(fā)酵和促進(jìn)菌體休眠。同時(shí),NO抑制合成氣發(fā)酵菌C.carboxidivorans的 H2ase活性而阻礙 H2利用[12]。XU 等[13]還得出合成氣中的NH3為H2ase的非競(jìng)爭(zhēng)抑制劑的結(jié)論。研究發(fā)現(xiàn),雜質(zhì)氣體NH3極易溶解于發(fā)酵培養(yǎng)基內(nèi),能快速轉(zhuǎn)化為NH4+且其濃度不斷累積[14],同時(shí)雜質(zhì)NH3還會(huì)腐蝕設(shè)備,工業(yè)上常利用濕式洗滌技術(shù)進(jìn)行脫氨處理[15],因而NH3對(duì)未來(lái)合成氣乙醇發(fā)酵工業(yè)化影響較大。相比,有關(guān)合成氣雜質(zhì)氣體NH3對(duì)合成氣乙醇發(fā)酵過(guò)程的影響還未有報(bào)道,且NH3對(duì)發(fā)酵代謝關(guān)鍵酶的作用也未有研究,為此,本文系統(tǒng)研究了不同初始濃度的NH4+(NH4Cl)對(duì)菌株A-fm4,C.ljungdahlii和 C.autoethanogenum DSM10061利用生物質(zhì)合成氣發(fā)酵乙醇的影響,并進(jìn)一步測(cè)定菌株在菌體生長(zhǎng)期和乙醇發(fā)酵期內(nèi)4種關(guān)鍵酶的活性變化。以期研究NH3對(duì)合成氣乙醇發(fā)酵的影響和作用機(jī)理,為未來(lái)合成氣乙醇發(fā)酵工業(yè)化發(fā)展提供參考依據(jù)。另外,實(shí)驗(yàn)中進(jìn)行了菌株耐銨能力的研究,便于為日后耐銨菌株馴化提供菌種材料,希望獲得能夠利用含NH3粗合成氣組分發(fā)酵的優(yōu)良菌株。
1 材料與方法
1.1 菌株
菌株 A-fm4富集自羊駝的新鮮糞便[16];菌株Closlridium ljungdahlii和Closlridium autoethanogenum DSM10061均購(gòu)自德國(guó)微生物菌種保藏中心(DSMZ)。
1.2 生物質(zhì)合成氣
實(shí)驗(yàn)用合成氣按照玉米秸稈氣化氣組分配制,成分及比例如下:CO 85.5%,H210%,CO24.5%。購(gòu)于河南源正科技發(fā)展有限公司。
1.3 培養(yǎng)基及配制方法
種子培養(yǎng)基(g/L)[17]:NaCl 0.80,NH4Cl 1.00,KCl 0.10,MgSO4·7H2O 0.20,CaCl20.04,KH2PO40.20,NaHCO31.00,酵母膏 1.00,MES[2-(N-嗎啡啉)乙磺酸]5.00,L-鹽酸半胱氨酸0.40,木糖5.00。添加微量元素溶液10 mL和維生素溶液10mL,pH 5.75。
發(fā)酵培養(yǎng)基(g/L)[17]:NH4Cl 1.00,NaCl 1.00,MgSO40.15,KH2PO40.10,CaCl20.04,胰蛋白胨2.00,酵母膏0.30,L-鹽酸半胱氨酸0.20,MES 10.00,礦質(zhì)元素溶液10 mL,維生素溶液10 mL,pH 7.0。
實(shí)驗(yàn)中發(fā)酵培養(yǎng)基初始NH4+濃度設(shè)置了6個(gè)梯度,分別為 0X、0.5X、1X、2X、4X 和 8X,因雜質(zhì)氣體NH3極易溶解于發(fā)酵培養(yǎng)基內(nèi),且能夠快速轉(zhuǎn)化為NH4+使得 NH4+濃度不斷累積[14],所以實(shí)驗(yàn)中利用NH4+的濃度進(jìn)行研究,即,將原發(fā)酵培養(yǎng)基的NH4Cl濃度標(biāo)記為1 X。
維生素溶液均采用0.22 μm的濾膜過(guò)濾除菌,待接種后添加至培養(yǎng)基內(nèi)。
上述所有培養(yǎng)基均準(zhǔn)確配制,每80 mL培養(yǎng)液分裝至200 mL厭氧培養(yǎng)瓶?jī)?nèi)(發(fā)酵培養(yǎng)基為每60 mL培養(yǎng)液分裝至300 mL厭氧培養(yǎng)瓶?jī)?nèi)),再轉(zhuǎn)入?yún)捬跖囵B(yǎng)箱內(nèi)添加刃天青0.5 mg/L,靜置直至瓶?jī)?nèi)顏色由粉色轉(zhuǎn)為無(wú)色時(shí)分別蓋上瓶塞,于121℃滅菌20 min(滅菌過(guò)程中瓶塞插入針頭)。滅菌后立刻拔去針頭且移至厭氧培養(yǎng)箱內(nèi),待溫度降至約37℃時(shí)進(jìn)行后續(xù)實(shí)驗(yàn)。
1.4 合成氣厭氧乙醇發(fā)酵
菌種活化參考文獻(xiàn)[16],將對(duì)數(shù)期的各菌種子液按10%接種量轉(zhuǎn)接至含有60 mL發(fā)酵培養(yǎng)液的厭氧培養(yǎng)瓶中,用注射器向厭氧瓶?jī)?nèi)加入240 mL合成氣(以上操作均在厭氧培養(yǎng)箱內(nèi)完成)。37℃,150 r/min搖床發(fā)酵7 d,每24 h取樣測(cè)OD600值及乙醇含量。
1.5 產(chǎn)物測(cè)定及計(jì)算方法
1.5.1 乙醇和乙酸含量測(cè)定
將1.5 mL發(fā)酵液于4℃,8 000 r/min離心10 min,上清液用于兩者濃度的測(cè)量。測(cè)定條件為:FID檢測(cè)器,柱子為30 m ×0.32 mm ×0.3 μm HP-FFAP,氮?dú)庾鳛檩d氣,起始3 min流速為1.9 mL/min,再以0.5 mL/min 升至 4 mL/min,分流比 50∶1,柱箱起始溫度40℃,保持1.5 min后以40℃/min達(dá)到210℃并保持2 min,色譜進(jìn)樣口和檢測(cè)器的溫度分別為200℃和250℃。
1.5.2 細(xì)胞干重和指標(biāo)計(jì)算
根據(jù)OD值及其對(duì)應(yīng)的細(xì)胞干重(DCW)可獲得發(fā)酵菌株的干重(y,g/L)與OD值(x)關(guān)系回歸方程式。
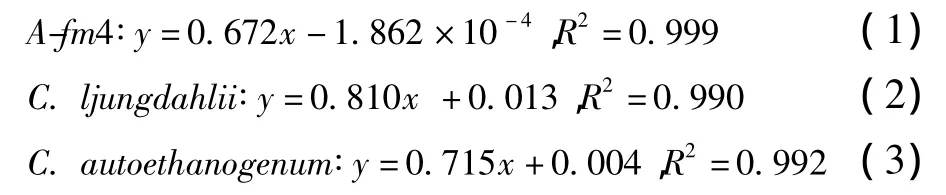
1.6 關(guān)鍵酶活性測(cè)定
關(guān)鍵酶H2ase、CODH和ADH活性測(cè)定方法分別參考文獻(xiàn)[18];關(guān)鍵酶ACK活性測(cè)定方法參考文獻(xiàn)[19]和[20],稍有改進(jìn)。反應(yīng)為:H2ase活性反應(yīng)體系具體包含:0.4 mL 1 mol/L Tris-HCl(pH=7.5),0.2 mL 0.04 mol/L甲基紫精,0.2 mL 0.04 mol/L二硫蘇糖醇,0.1 mL體積分?jǐn)?shù)為5%的Triton-X100,2 mL脫氣去離子水;CODH活性反應(yīng)體系具體包含:0.8 mL 0.5 mol/L Tris-HCl(pH=6.8),0.2 mL 0.04 mol/L甲基紫精(MV),0.2 mL 0.04 mol/L二硫蘇糖醇,0.1 mL體積分?jǐn)?shù)為5%的Triton-X100,1.2 mL脫氣去離子水。ADH活性反應(yīng)體系具體包含:0.4 mL 1 mol/L Tris-Hcl(pH=7.5),0.12 mL 0.01 M NADH,0.4 mL 0.1 M 乙醛,0.5 mL 0.08 mol/L 二硫蘇糖醇,0.1 mL體積分?jǐn)?shù)為5%的Triton-X100,1.5 mL脫氣去離子水;ACK活性反應(yīng)體系具體包含:0.145 mL 1 mol/L Tris-HCl(pH=7.5),0.1 mL 2 mol/L乙酸鉀,0.25 mL 0.08 mol/L二硫蘇糖醇,0.1 mL體積分?jǐn)?shù)為5%的Triton-X100,0.1 mL 0.1 mol/L MgCl2,0.1 mL 0.1 mol/L ATP,0.2 mL 3.5 mol/L 中性NH4OH-HCl,2 mL脫氣去離子水。且二硫蘇糖醇溶液現(xiàn)配現(xiàn)用。
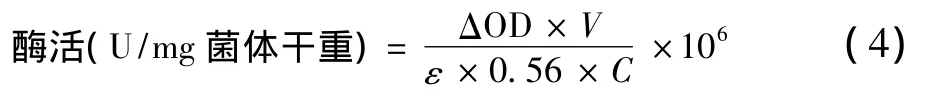
式中:△OD為吸收值每分鐘降低的變化值,min-1;V為反應(yīng)體系的體積,mL;E為摩爾消光系數(shù),103L/(mol·cm);0.56為96孔板的光程,cm;C為測(cè)定中所加入菌體干物質(zhì)量,g;106為摩爾轉(zhuǎn)化為微摩爾的系數(shù)。酶活單位:單位干菌體每分鐘還原1 μmol MV(1e-1還原)。
1.7 NH4+對(duì)菌株生長(zhǎng)半抑制濃度
NH4+對(duì)發(fā)酵菌株生長(zhǎng)半抑制濃度(Ki)的計(jì)算參考文獻(xiàn)[21],通過(guò)對(duì)比不同發(fā)酵菌株的Ki值大小來(lái)衡量菌株的NH4+耐受能力強(qiáng)弱。
1.8 統(tǒng)計(jì)分析方法
為比較在不同初始NH4+濃度下對(duì)菌株發(fā)酵乙醇產(chǎn)量及其各發(fā)酵參數(shù)的影響,同時(shí)分析比較對(duì)兩時(shí)期4種關(guān)鍵酶活性水平的差異,實(shí)驗(yàn)分別采用了方差分析和t值檢驗(yàn),以上各統(tǒng)計(jì)分析均由軟件SPSS16.0完成。
2 結(jié)果與討論
2.1 初始N H4+濃度對(duì)合成氣發(fā)酵菌株生長(zhǎng)的影響
不同梯度濃度NH4+對(duì)發(fā)酵菌株的生長(zhǎng)影響見(jiàn)圖1。菌株 A-fm4、C.ljungdahlii和 C.autoethanogenum的生物量分別在NH4+濃度0.5X、0X和0X于發(fā)酵4 d、1 d和2 d達(dá)到最大,菌體量分別為:0.045,0.166和0.097 g/L。菌株 A-fm4和 C.autoethanogenum DSM10061的生長(zhǎng)過(guò)程受NH4+影響明顯,NH4+對(duì)菌株C.ljungdahlii的影響主要表現(xiàn)在生長(zhǎng)穩(wěn)定期及以后,且隨NH4+梯度濃度增大對(duì)菌株生長(zhǎng)量和生長(zhǎng)速率具有抑制能力。
由此可知,合成氣發(fā)酵菌株對(duì)無(wú)機(jī)氮源的利用有限,發(fā)酵液內(nèi)NH4+累積濃度高于9.35 mmol/L時(shí)生長(zhǎng)受到不同程度的抑制作用。據(jù)報(bào)道,Escherichia coli已作為一種模式生物進(jìn)行研究細(xì)菌對(duì)NH4+的吸收和排除是經(jīng)過(guò)一種高K+親和性的K+-NH4+交換系統(tǒng)(Kdp)[22],其中 Buurman 等[23]對(duì) E.coli在高濃度NH4+環(huán)境中代謝活動(dòng)的研究發(fā)現(xiàn)在高濃度NH4+條件下需要一種額外轉(zhuǎn)運(yùn)系統(tǒng),即Kdp,來(lái)負(fù)責(zé)NH4+的攝入和泵出維持胞內(nèi)外NH4+正常梯度及胞內(nèi)pH的穩(wěn)定,且高濃度NH4+使得菌體對(duì)能量的利用效率顯著降低。同樣,葉貴子等[21]對(duì)產(chǎn)琥珀酸放線桿菌的研究也得到相似結(jié)果。雖然目前還未見(jiàn)針對(duì)合成氣厭氧發(fā)酵菌在Kdp系統(tǒng)與能量效率方面的確切報(bào)道,但可推測(cè)在本實(shí)驗(yàn)發(fā)酵菌體細(xì)胞內(nèi)也存在類(lèi)似機(jī)制。另外,數(shù)據(jù)顯示K+的水合半徑為5.32,近于NH4+的5.37,NH4+對(duì)K+在細(xì)胞膜同一載體上可競(jìng)爭(zhēng)同一結(jié)合部位,這一競(jìng)爭(zhēng)會(huì)破壞K+對(duì)細(xì)胞正常的生理功能,且隨NH4+濃度的增大不斷顯著,進(jìn)而導(dǎo)致對(duì)菌體的生長(zhǎng)抑制。
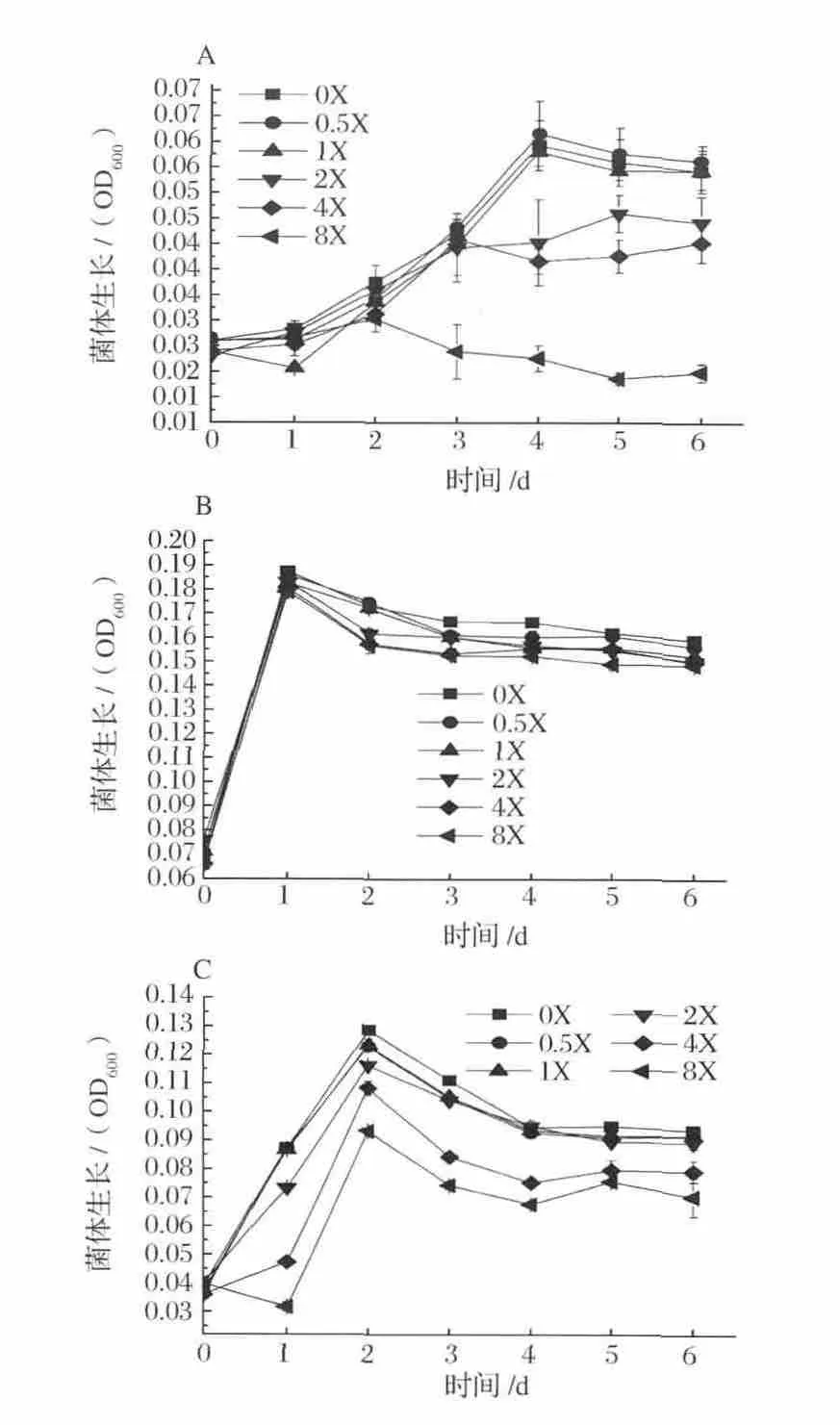
圖1 不同梯度NH4+濃度下發(fā)酵菌生長(zhǎng)曲線Fig.1 Growth curve of organisms at different concentrations of NH4+
2.2 初始NH4+濃度對(duì)合成氣發(fā)酵乙醇特性的影響
菌株 A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii的乙醇產(chǎn)量均在NH4+濃度0X下于發(fā)酵5 d、2 d和3 d達(dá)到最大,其乙醇量分別為124.56,137.71和137.67 mg/L(圖2)。其中,菌群A-fm4的乙醇發(fā)酵受NH4+濃度影響最大,其次為菌株C.autoethanogenum DSM10061,再次是菌株 C.ljungdahlii。NH4+濃度高于0.5X(9.35 mmol/L)時(shí)開(kāi)始減緩菌株乙醇發(fā)酵速率,當(dāng)濃度高于4X(74.8 mmol/L)其抑制作用更顯著,因此不存在或低濃度NH4+會(huì)促進(jìn)乙醇發(fā)酵,高濃度NH4+能降低乙醇發(fā)酵能力。
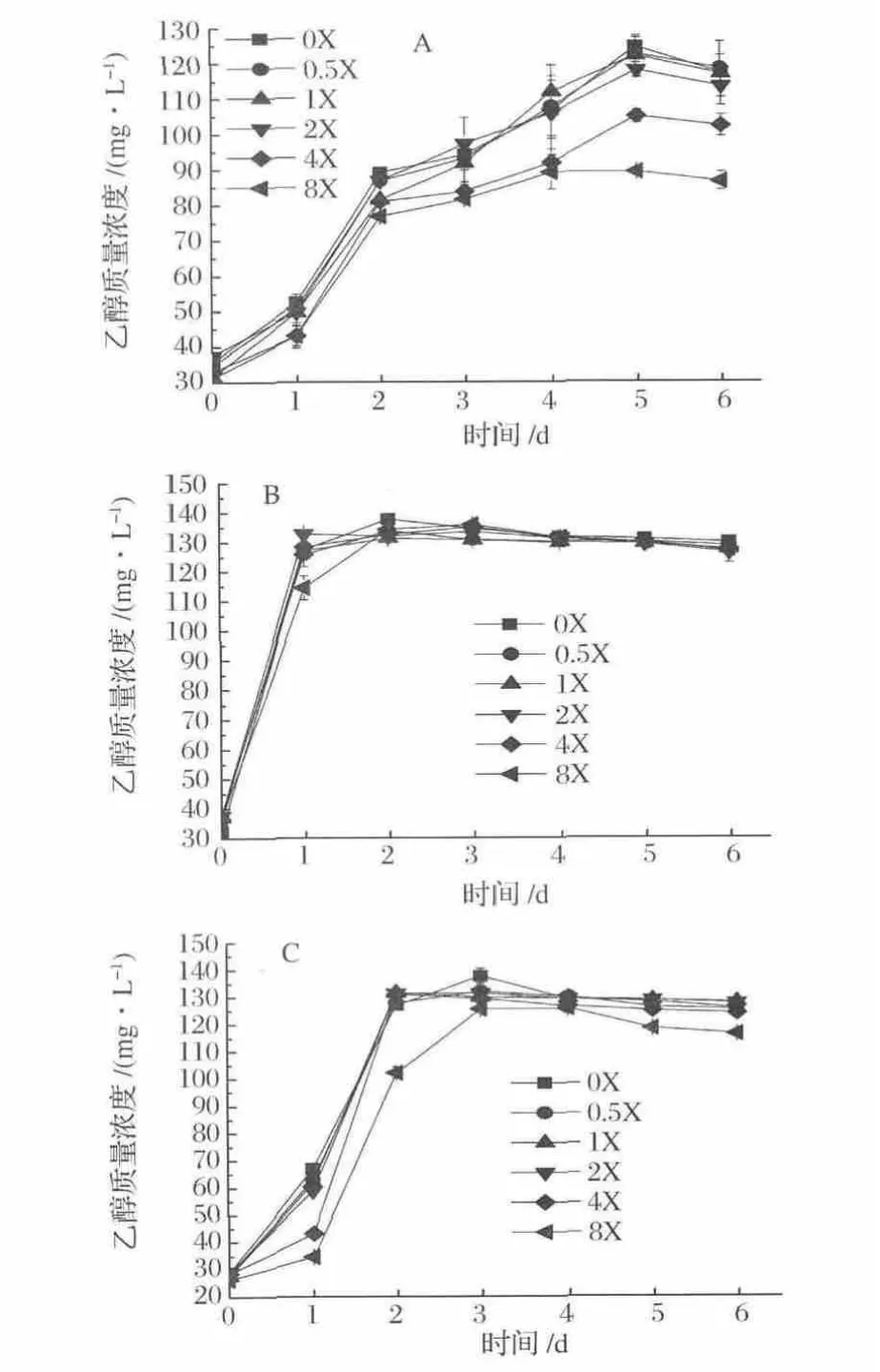
圖2 不同梯度NH4+濃度下乙醇產(chǎn)量Fig.2 Ethanol production fermented at different concentrations of NH4+
2.3 初始N H4+濃度對(duì)合成氣發(fā)酵乙酸特性的影響
圖3為不同梯度濃度NH4+對(duì)合成氣發(fā)酵菌株發(fā)酵乙酸特性的影響。菌株A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii的乙酸產(chǎn)量分別在NH4+濃度2X、2X和0X或2X下于發(fā)酵5 d、4 d和3 d或4 d達(dá)到最大,乙酸量分別為218.19,302.72和304.80或304.95 mg/L。相比菌株A-fm4和C.autoethanogenum,菌株C.ljungdahlii乙酸發(fā)酵受 NH4+影響較弱,另外,NH4+濃度0.5X~4X(9.35~74.8 mmol/L)能不同程度地促進(jìn)菌株A-fm4、C.autoethanogenum DSM10061和C.ljungdahlii乙酸發(fā)酵速率。結(jié)果表明,NH4Cl濃度在18.7~74.8 mmol/L會(huì)促進(jìn)乙酸的發(fā)酵,濃度為149.6 mmol/L抑制乙酸發(fā)酵。
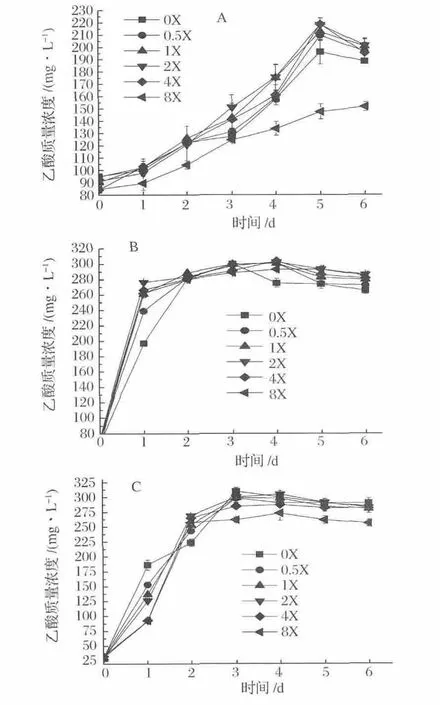
圖3 不同梯度NH4+濃度下乙酸產(chǎn)量Fig.3 Acetate production fermented at different concentrations of NH4+
綜合分析發(fā)酵菌株在不同梯度NH4+環(huán)境內(nèi)的發(fā)酵情況可得:低濃度NH4+利于乙醇和乙酸發(fā)酵,高濃度NH4+對(duì)乙醇和乙酸均表現(xiàn)出抑制作用。當(dāng)NH4+濃度高于9.35 mmol/L時(shí),乙醇發(fā)酵隨濃度的增大而抑制顯著,NH4+濃度為18.7~74.8 mmol/L時(shí)會(huì)促進(jìn)乙酸的發(fā)酵。發(fā)酵培養(yǎng)液內(nèi)不含NH4+時(shí)利于更多的代謝流流向乙醇,促進(jìn)乙醇且減弱乙酸發(fā)酵。
2.4 初始NH4+濃度對(duì)合成氣發(fā)酵代謝關(guān)鍵酶活性的影響
不同初始NH4+濃度對(duì)合成氣厭氧發(fā)酵菌株發(fā)酵代謝途徑關(guān)鍵酶活性變化的影響結(jié)果見(jiàn)圖4。H2ase和CODH酶活水平隨NH4+濃度增大而抑制極顯著(P<0.01),生長(zhǎng)期酶活水平高于發(fā)酵期,NH4+濃度低于2X(37.4 mmol/L)促進(jìn)菌株A-fm4該酶的高水平維持(A1~C1,A2~C2);除菌株 C.ljungdahlii外(B3),NH4+濃度大于2X(37.4 mmol/L)時(shí)ADH酶活大小受到抑制作用,其中菌株A-fm4生長(zhǎng)期酶活水平大于發(fā)酵期,兩時(shí)期酶活均受NH4+影響顯著(A3),菌株C.autoethanogenum DSM10061生長(zhǎng)期酶活水平受NH4+影響,而發(fā)酵期 ADH酶活大小趨于穩(wěn)定(C3)。相比,菌株C.ljungdahlii兩時(shí)期ADH酶活均受NH4+影響較小;對(duì)于ACK,菌株A-fm4和C.ljungdahlii在低濃度和高濃度NH4+下其活性水平均受抑制,NH4+濃度1X~2X(18.7 mmol/L~37.4 mmol/L)可促進(jìn)ACK活性,且生長(zhǎng)期酶活水平一般高于發(fā)酵期(A4和B4),而菌株C.autoethanogenum DSM10061在NH4+濃度低于2X(37.4 mmol/L)時(shí)可維持較高活性水平,高濃度NH4+抑制酶活(C4)。因此,低濃度NH4+可促進(jìn)關(guān)鍵酶活性,高濃度NH4+具有抑制作用,且生長(zhǎng)期酶活水平較大于發(fā)酵期的值。
Wood-Ljungdahl代謝途徑中H2ase和CODH可分別通過(guò)催化氧化H2和CO產(chǎn)生還原力和質(zhì)子,其中H2ase為代謝過(guò)程中還原力獲得的必需酶,H2ase活性水平會(huì)直接影響到 CO的利用效率,高活性H2ase可弱化由CO經(jīng)CODH產(chǎn)生還原力的途徑,促進(jìn)CO作為碳源和能源,而H2ase低活性時(shí)其結(jié)果相反[1]。由上述結(jié)果可知,NH4+濃度能影響 H2ase和CODH的活性大小,而對(duì)H2ase的活性影響顯著,因此從酶學(xué)層面上可解釋NH4+濃度大于2X時(shí)逐漸抑制發(fā)酵菌株的生長(zhǎng),而低濃度或不存在NH4+可促進(jìn)菌體的生長(zhǎng)(圖1)。另外,NH4+對(duì)H2ase和CODH的抑制機(jī)制可理解如下,即,NH3和NH4+在發(fā)酵液中快速轉(zhuǎn)化,NH3易通過(guò)細(xì)胞膜進(jìn)入胞內(nèi),繼續(xù)電離NH4+[24],改變胞漿內(nèi)外的原有正常NH4+梯度,使得H2ase和CODH暴露于更多NH4+環(huán)境內(nèi),自由NH4+的增多影響到酶結(jié)構(gòu)域的電位變化導(dǎo)致其活性降低,NH4+濃度越高抑制酶活作用越強(qiáng)。研究證明,H2ase和CODH常依附于胞膜內(nèi)側(cè)[25],鑒于高濃度NH4+會(huì)對(duì)其抑制及NH3易出入細(xì)胞膜的能力,可假設(shè)在菌株發(fā)酵中存在于菌體胞內(nèi)的H2ase和CODH受NH4+抑制濃度可能為37.4 mmol/L。ADH是 Wood-Ljungdahl中乙醇代謝的關(guān)鍵酶,實(shí)驗(yàn)中菌株 A-fm4的ADH酶活受NH4+濃度影響顯著,當(dāng)NH4+濃度在2X以?xún)?nèi)時(shí)該酶活性維持較高水平(圖4-A3),進(jìn)而促進(jìn)乙醇發(fā)酵(圖2-A)。對(duì)于該菌群,理論上ADH酶活應(yīng)在發(fā)酵期達(dá)到較高水平,實(shí)際上發(fā)酵期獲得最大乙醇產(chǎn)量但其活性降低,可能的解釋是ADH活性雖在后期降低,但足以滿(mǎn)足合成乙醇所必須的酶活大小,或是在發(fā)酵期ADH隨著發(fā)酵環(huán)境的改變ADH活性降低,但在菌體內(nèi)的表達(dá)量卻在增加來(lái)滿(mǎn)足乙醇代謝[18]。針對(duì)菌株 C.ljungdahlii和 C.autoethanogenum DSM10061,ADH的活性受NH4+影響小,酶活水平相當(dāng),具有一定的耐受能力,有學(xué)者[26]對(duì)菌體C.ljungdahlli內(nèi)參基因穩(wěn)定性及重要基因表達(dá)量的研究發(fā)現(xiàn),與乙醇合成相關(guān)的重要基因aor2在不同碳源內(nèi)的表達(dá)量穩(wěn)定,這可提示本實(shí)驗(yàn)兩菌株在不同濃度NH4+發(fā)酵環(huán)境中該基因或ADH相關(guān)重要基因也存在類(lèi)似穩(wěn)定表達(dá)的能力進(jìn)而使得ADH活性水平穩(wěn)定,只是對(duì)于C.autoethanogenum DSM10061這種穩(wěn)定表達(dá)在乙醇發(fā)酵期得到主要體現(xiàn)。另外,菌株C.ljungdahlii和C.autoethanogenum DSM10061在乙醇發(fā)酵期的ADH雖具有一定的耐受性(圖4-B3,B4),但乙醇產(chǎn)量卻低于低NH4+濃度(小于2X)的發(fā)酵乙醇產(chǎn)量,關(guān)鍵問(wèn)題是在較高NH4+濃度下H2ase活性受到抑制,進(jìn)而在代謝流中用于乙醇代謝的還原力減少導(dǎo)致乙醇合成降低。
合成氣厭氧發(fā)酵的乙酸發(fā)酵離不開(kāi)關(guān)鍵酶ACK,實(shí)驗(yàn)發(fā)現(xiàn),NH4+濃度在18.7~37.4 mmol/L可促進(jìn)其活性,相繼乙酸產(chǎn)量增加(圖3)。室溫下,乙酸解離常數(shù)pKa為4.75,在菌體胞漿的偏中性中其主要以 Ac-和 H+存在,因此在1X和2X下胞內(nèi)NH4+的電離利于胞漿正常pH范圍的穩(wěn)定,減少大量乙酸在胞內(nèi)對(duì)酶促反應(yīng)的抑制,隨分子乙酸由細(xì)胞調(diào)節(jié)機(jī)制[27]不斷移出胞內(nèi)使發(fā)酵液內(nèi)乙酸濃度不斷增加。基于0X下促進(jìn)乙醇而降低乙酸發(fā)酵,因此當(dāng)不存在NH4+時(shí)利于更多的代謝流流向乙醇。
2.5 發(fā)酵菌株對(duì)NH4+的耐受能力對(duì)比
由表1知,NH4+對(duì)不同種類(lèi)發(fā)酵菌的生長(zhǎng)抑制程度不同,3種發(fā)酵出發(fā)菌中僅菌株A-fm4對(duì)NH4+最為敏感,其 Ki值為 123.61 mmol/L。相比,菌株 C.ljungdahlii和 C.autoethanogenum DSM10061的 Ki值分別為208.13 mmol/L和240.37 mmol/L,耐NH4+能力較強(qiáng),這或許為未來(lái)該菌直接利用含氨氣的粗制合成氣發(fā)酵乙醇提供重要啟示。同時(shí)菌株C.ljungdahlii和C.autoethanogenum DSM10061可為未來(lái)工業(yè)菌株耐銨馴化提供菌種材料,耐銨菌種的使用利于合成氣厭氧乙醇發(fā)酵的工業(yè)化進(jìn)程,為降低粗制合成氣脫氨設(shè)備的資金投入甚至刪減脫氨工藝工序都具有重要意義。
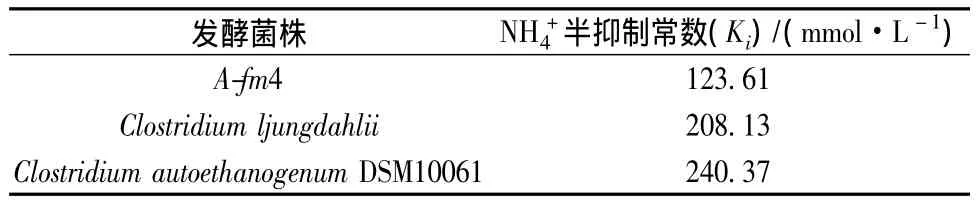
表1 NH4+對(duì)發(fā)酵菌株的半抑制常數(shù)(Ki)Table 1 Ammnia half-inhibitin constant(Ki)for fermentation microorganisms
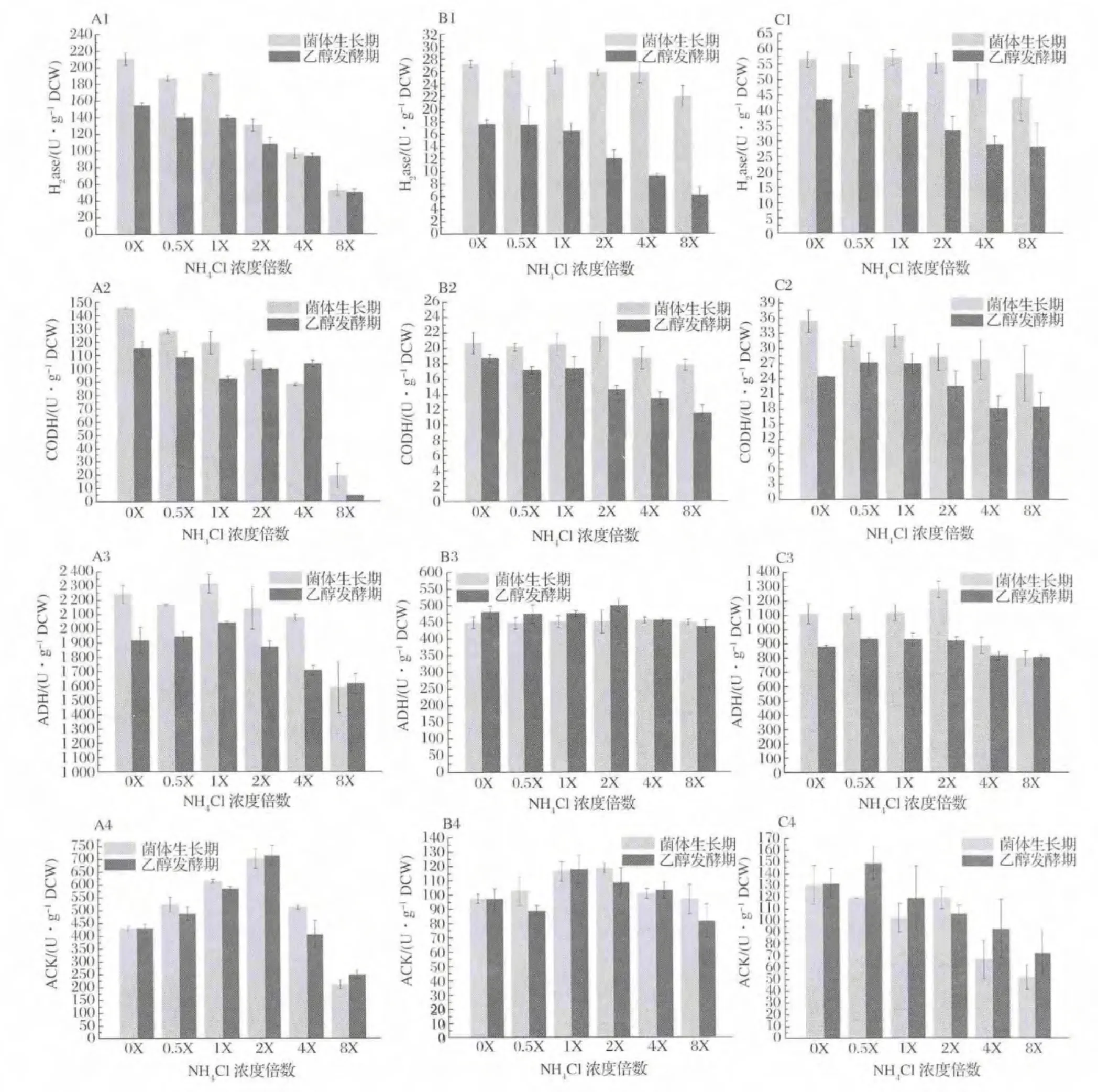
圖4 NH4+對(duì)菌株發(fā)酵過(guò)程中兩時(shí)期關(guān)鍵酶活性的影響Fig.4 The activity of key enzymes existed in growth period and ethanol fermentation period at different concentrations of NH4+
3 結(jié)論
(1)發(fā)酵培養(yǎng)基內(nèi)不同初始濃度的NH4+對(duì)菌株A-fm4,C.ljungdahlii和 C.autoethanogenum DSM10061發(fā)酵過(guò)程中的生長(zhǎng)影響顯著,其中對(duì)菌株C.ljungdahlii的影響主要表現(xiàn)在生長(zhǎng)穩(wěn)定期及以后。結(jié)果表明,菌株 A-fm4、C.ljungdahlii和 C.autoethanogenum DSM10061分別在NH4+濃度0.5X、0X和0X下獲得最大生物量,分別為0.045,0.166和0.097 g/L。當(dāng)NH4+濃度高于9.35 mmol/L時(shí),隨NH4+梯度濃度增大對(duì)菌株生長(zhǎng)量和生長(zhǎng)速率具有抑制能作用。
(2)綜合分析合成氣發(fā)酵過(guò)程可知:低濃度NH4+利于乙醇和乙酸發(fā)酵,高濃度NH4+對(duì)乙醇和乙酸均表現(xiàn)出抑制作用。當(dāng)NH4+濃度高于9.35 mmol/L時(shí),乙醇發(fā)酵隨濃度的增大而抑制顯著,NH4+濃度為18.7~74.8 mmol/L時(shí)會(huì)促進(jìn)乙酸的發(fā)酵。發(fā)酵培養(yǎng)液內(nèi)不含NH4+時(shí)利于更多的代謝流流向乙醇,促進(jìn)乙醇且減弱乙酸發(fā)酵。
(3)關(guān)鍵酶 H2ase,CODH,ADH和 ACK均受高濃度(149.6 mmol/L)NH4+抑制,且菌體生長(zhǎng)期的酶活水平一般高于發(fā)酵期,H2ase和CODH隨NH4+濃度升高而酶活下降極顯著,H2ase活性的高低顯著影響發(fā)酵過(guò)程中還原力的供給,直接影響菌體生長(zhǎng)和乙醇發(fā)酵。除菌株A-fm4外,菌株C.ljungdahlii和C.autoethanogenum DSM10061的ADH受NH4+影響較小,推測(cè)該酶在胞內(nèi)具有穩(wěn)定表達(dá)機(jī)制。NH4+濃度1X~2X(18.7~37.4 mmol/L)可促進(jìn)ACK活性,且不存在NH4+時(shí)也會(huì)降低該酶活性。
(4)菌株 C.ljungdahlii和 C.autoethanogenum DSM10061的Ki值分別為208.13 mmol/L和240.37 mmol/L,耐銨能力較強(qiáng),可為未來(lái)耐銨菌種的馴化提供菌種材料,為將來(lái)降低脫氨設(shè)備資金投入和減少合成氣凈化的復(fù)雜工序提供技術(shù)準(zhǔn)備。
[1] Latif H,Zeidan A A,Nielsen A T,et al.Trash to treasure:production of biofuels and commodity chemicals via syngas fermenting microorganisms[J].Current Opinion in Biotechnology,2014,27:79-87.
[2] Vohra M,Manwar J,Manmode R,et al.Bioethanol production:Feedstock and current technologies[J].Journal of Environmental Chemical Engineering,2014,2(1):573-584.
[3] Bengelsdorf F R,Straub M,Dürre P.Bacterial synthesis gas(syngas)fermentation[J].Environmental Technology,2013,34(13):1 639-1 651.
[4] Henstra A M,Sipma J,Rinzema A,et al.Microbiology of synthesis gas fermentation for biofuel production[J].Current Opinion in Biotechnology,2007,18(3):200-206.
[5] Kirkels A F,Verbong G P.Biomass gasification:Still promising?A 30-year global overview[J].Renewable and Sustainable Energy Reviews,2011,15(1):471-481.
[6] Phillips J,Klasson K,Clausen E,et al.Biological production of ethanol from coal synthesis gas[J].Applied Biochemistry and Biotechnology,1993,39(1):559-571.
[7] Abubackar H N,Veiga M C,Kennes C.Biological conversion of carbon monoxide:rich syngas or waste gases to bioethanol[J].Biofuels,Bioproducts and Biorefining,2011,5(1):93-114.
[8] Somerville C,Youngs H,Taylor C,et al.Feedstocks for lignocellulosic biofuels[J].Science(Washington),2010,790(329):790-792.
[9] Rollins M L,Reardon L,Nichols D,et al.National Energy Technology Lab.,Pittsburgh,PA(US);National Energy Technology Lab.,Morgantown,WV(US),2002.
[10] Cotter J L,Chinn M S,Grunden A M.Influence of process parameters on growth of Clostridium ljungdahlii and Clostridium autoethanogenum on synthesis gas[J].Enzyme and Microbial Technology,2009,44(5):281-288.
[11] Ahmed A,Cateni B G,Huhnke R L,et al.Effects of biomass-generated producer gas constituents on cell growth,product distribution and hydrogenase activity of Clostridium carboxidivorans P7T[J].Biomass and Bioenergy,2006,30(7):665-672.
[12] Ahmed A,Lewis R S.Fermentation of biomass-generated synthesis gas:Effects of nitric oxide[J].Biotechnology and Bioengineering,2007,97(5):1 080-1 086.
[13] XU D,Lewis R S.Syngas fermentation to biofuels:Effects of ammonia impurity in raw syngas on hydrogenase activity[J].Biomass and Bioenergy,2012,45:303-310.
[14] CHEN S,LING J,Blancheton J-P.Nitrification kinetics of biofilm as affected by water quality factors[J].Aquacultural Engineering,2006,34(3):179-197.
[15] Daniell J,K?pke M,Simpson S D.Commercial biomass syngas fermentation[J].Energies,2012,12(5):5 372-5 417.
[16] 宋安東,馮新軍,王風(fēng)芹,等.合成氣發(fā)酵制取乙醇微生物的對(duì)比[J].生物工程學(xué)報(bào),2013,29(3):242-249.
[17] 王風(fēng)芹,張炎達(dá),謝慧,等.生物質(zhì)氣化氣發(fā)酵生產(chǎn)乙醇優(yōu)良菌株的篩選[J].農(nóng)業(yè)工程學(xué)報(bào),2015,31(7):221-226.
[18] Ukpong M N,Atiyeh H K,De Lorme M J,et al.Physiological response of Clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas‐fed bioreactor[J].Biotechnology and Bioengineering,2012,109(11):2 720-2 728.
[19] Aceti D J,F(xiàn)erry J G.Purification and characterization of acetate kinase from acetate-grown Methanosarcina thermophila.Evidence for regulation of synthesis[J].Journal of Biological Chemistry,1988,263(30):15 444-15 448.
[20] Sridhar J Eiteman M A,Wiegel J W.Elucidation of enzymes in fermentation pathways used by Clostridium thermosuccinogenes growing on inulin[J].Applied and Environmental Microbiology,2000,66(1):246-251.
[21] 葉貴子,姜岷,陳可泉,等.耐銨型產(chǎn)琥珀酸放線桿菌的選育及銨離子對(duì)其生長(zhǎng)代謝的影響[J].生物工程學(xué)報(bào),2010,26(2):183-188.
[22] Marinus M,Neidhardt F,Curtiss I,et al.Escherichia coli and Salmonella:cellular and molecular biology[J].Escherichia coli and Salmonella:Cellular and Molecular Biology,1996,2:1 579-1 601.
[23] Buurman E T,De Mattos M J T,Neijssel O M.Futile cycling of ammonium ions via the high affinity potassium uptake system(Kdp)of Escherichia coli[J].Archives of Microbiology,1991,155(4):391-395.
[24] Khademi S,O'connell J,Remis J,et al.Mechanism of ammonia transport by Amt/MEP/Rh:structure of AmtB at 1.35 ?[J].Science,2004,305(5 690):1 587-1 594.
[25] Bagyinka C,Kovács K L,Rak E.Localization of hydrogenase in Thiocapsa roseopersicina photosynthetic membrane[J].Biochemical Journal,1982,202(1):255-258.
[26] 劉娟娟.Clostridium ljungdahlii內(nèi)參基因穩(wěn)定性評(píng)價(jià)及其在重要基因相對(duì)表達(dá)量研究中的應(yīng)用[D].重慶:西南大學(xué),2013:41-50.
[27] Russell J.Another explanation for the toxicity of fermentation acids at low pH:anion accumulation versus uncoupling[J].Journal of Applied Bacteriology,1992,73(5):363-370.
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
