Txt5腺病毒載體構建及其在心肌細胞中的表達
2015-12-03 09:18:20李佑鋒周作瓊吳秀山范雄偉李永青
懷化學院學報 2015年5期
彭 浩,陳 宇,李佑鋒,朱 旋,周作瓊,吳秀山,范雄偉,李永青
(湖南師范大學 心臟發育研究中心 蛋白質化學及魚類發育生物學教育部重點實驗室,湖南 長沙 410081)
Txt5位于2號染色體短臂11區2帶,長度為3239 bp,編碼蛋白含391個氨基酸,從N 端開始依次為LisH、CTLH、CRA 結構域.LisH 結構域被發現存在于很多病癥相關基因的產物之中[1-2].已經有研究證明,Txt5可能參與了MAPK 途徑對細胞生命活動的調控[3].RT-PCR 分析顯示這一基因在人類早期胚胎的多個組織中有表達,在心臟、肝和腎中表達水平較強.腺病毒轉染能夠使外源基因在原代細胞中高效表達[4],為了進一步研究Txt5 基因的功能,本文利用Ad-Easy 腺病毒系統研制了小鼠Txt5 基因的過表達腺病毒[5],為后續研究Txt5表達及生物學功能提供實驗依據.
1 材料和方法
1.1 實驗材料
1.1.1 生物材料
解剖小鼠取心臟提取mRNA,反轉錄獲得cDNAs.連接載體pMD18-T 購買于Takara.穿梭載體pAdTrack-CMV、BJ5183 菌株以及HEK293 A 細胞均為實驗室所保存.
1.1.2 所用主要試劑
PmeΙ、Pac Ι 內切酶均購買于NEB公司,T4 DNA連接酶、Sal Ι和Hind III 限制性內切酶買于Takara公司;膠回收純化試劑盒購買于康維世紀公司,逆轉錄試劑盒購買于Thermo fisher公司,小量快速質粒提取試劑盒購買于康維世紀公司;兔源HA 一抗為美國圣克魯斯公司生產;Lipofectamine 2000 試劑購買于英濰捷基公司;高糖DMEM 培養基、F12 培養基和胎牛血清購買于Hyclone公司.
1.2 方法
1.2.1 穿梭質粒pAdTtrack-CMV-Txt5的構建
根據NCBI 數據庫,設計特異引物如下:5'-AGATCTATGGATCAGTGCGTGACGGTG-3'和5'-AAGCTTTCAAGCGTAATCTGGAACATCGTATGGGTA GAAAAATATCTGTTTGGCATCTCCT-3'.
PCR 擴增目標片段經測序正確后連接至pMD-18 T 載體,轉化大腸桿菌感受態細胞,卡那霉素篩選陽性克隆.
1.2.2 pAd-Txt5的獲得
質粒小提試劑盒提取穿梭質粒pAdTrackCMV-Txt5,用Pme Ι 酶37℃酶切過夜線性化,純化回收酶切產物.將產物轉化進入BJ5183 感受態,通過其內的重組組件發生同源重組,涂布于卡那抗性平板上篩選.質粒小提試劑盒提取重組質粒,37℃Pac Ι 酶切過夜線性化,2%凝膠電泳觀察是否有兩條帶,利用酚氯仿法抽提DNA,保存于-20℃.
1.2.3 線性化pAd-Txt5 轉染HEK 293 A 細胞
復蘇凍存于液氮中的HEK 293A 細胞,培養于10 cm 培養皿中.當細胞融合率達80%左右后,取線性化的pAd-Txt5 4 μg,通過Lipofectamine 2000 轉染HEK 293 A 細胞.5 h 后換含10%胎牛血清的培養液10 ml,轉染12 h 后通過熒光顯微鏡觀察HEK 293 A 細胞綠色熒光情況,之后每隔12 或24 h 觀察熒光并拍照.在第10 d 左右,會有大團細胞漂浮.吸打下細胞,將細胞液用液氮反復凍融2~3次,2 000 r/min,10 min.上清即病毒粗提液,EP 管分裝于-80℃保存.
1.2.4 Ad-Txt5 病毒顆粒的擴增
10 cm 皿中293 A 細胞密度約達80%的時候,開始轉染:取0.4 ml 病毒粗提液溶液加到皿中,在第3 d左右,會有大團細胞漂浮.將細胞吸打下來,將細胞液用液氮反復凍融2~3次,2 000 r/min,10 min.上清即病毒粗提液,EP 管分裝于-80℃保存.
1.2.5 分離和培養原代心肌細胞
解剖出生一或兩天SD 大鼠,冰上剪取心臟的心室心尖,PBS 中洗去血液.接著剪碎組織,先后于0.06%胰酶中和II 型膠原酶中進行消化.然后將消化完全后的液體加入10 mL 含10% FBS 培養基中終止消化,混勻,2 000 r/min,10 min,倒去上清.加5 mL含10% FBS的培養基再洗一次,2 000 r/min,5 min.倒去上清,用20 mL 含10% FBS和0.067 mg/mL 濃度Brdu的DMEM-F12 培養液混勻細胞,200 目篩過濾細胞液至兩個10 cm 皿中.差速貼壁2 h 后,將懸浮細胞液體吸取到10 cm 皿中,2 d 后更換培養基.
1.2.6 免疫印跡鑒定
將Ad-Txt5 重組腺病毒和Ad-GFP 空載腺病毒感染心肌細胞,72 h 之后可見到綠色熒光,收取心肌細胞,提RNA 及蛋白.將蛋白進行電泳:配置電泳膠,加入等體積的蛋白樣品40 μg,120 V 電泳20 min;濕轉:100 V,90 min 將蛋白轉到PVDF 膜上,15 V,1 h.加封閉液(TBST 加5%脫脂奶粉),37℃封閉1~2 h;用TBST 洗去牛奶的顏色;兔源HA 一抗孵育,4℃過夜;TBST 溶液洗膜,3次,每次5 min,然后進行二抗的孵育,加入二抗,室溫1 h;TBST 溶液再次洗膜3次,每次10 min;加入ECL 系統顯色,壓片,洗膜,觀察結果.
1.2.7 RT-PCR
按mRNA Selective PCR Kit 試劑盒說明書取適量的RNA 進行逆轉錄.利用cDNA為模板,進行一般的PCR實驗,反應程序:95℃,預變性2 min,95℃變性30 s,60℃退火30 s,72℃延伸1 min,30個循環;72℃延伸8 min.4℃保存,通過凝膠電泳來分析PCR 結果.
RT-GAPDH-S:5'AGCCCAGAACATCATCCCT3'
RT-GAPDH-A:5'GGTCCTCAGTGTAGCCCAAG3'
2 結果分析
2.1 pAd-Txt5 質粒的獲得與鑒定
將pMD18-T-Txt5 利用Sal Ι和Hind III 限制性內切酶進行雙酶切,純化回收目的片段連接于同樣雙酶切后的穿梭載體pAdtrack-CMV 上,接著進行轉化,提取出穿梭質粒pAdTrack-CMV-Txt5.再次Sal Ι和Hind III 限制性內切酶雙酶切穿梭質粒進行鑒定,1.8%瓊脂糖凝膠電泳結果有2條帶,上方條帶約為9 200 bp,下方條帶約為1 000 bp,證明穿梭質粒構建成功(圖1A).將重組完成后的腺病毒質粒用Pac Ι 酶切后進行1.8%瓊脂糖凝膠電泳結果有2條帶,上方條帶約為34 kb,下方條帶約為4 kb 亮帶,說明腺病毒質粒pAd-Txt5 成功獲得(圖1B).
2.2 腺病毒顆粒的獲得
腺病毒質粒pAd-Txt5 線性化之后轉染HEK 293A細胞,24 h 后可見到綠色熒光(GFP),之后熒光逐漸增多,48 h 時熒光大量出現(圖2),12 d 左右,細胞大團飄起,移液器將細胞吸打下來,反復凍融獲得病毒溶液,過濾后再次感染293 A 細胞,12 h 后依然可以看到綠色熒光.
2.3 Txt5 過表達腺病毒在心肌細胞中的表達

圖1 穿梭質粒pAdTrack-CMV-Txt5和腺病毒質粒pAd-Txt5酶切后電泳圖
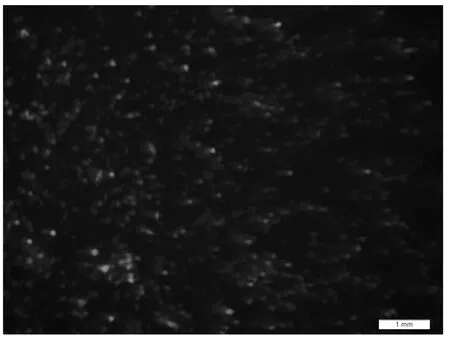
圖2 線性化pAd-Txt5轉染48 h后觀察結果(×20)
Txt5 基因通過IRES 序列與腺病毒載體基本骨架中的GFP 序列共表達,感染了腺病毒的心肌細胞就會出現綠色熒光.Txt5 過表達腺病毒感染SD 大鼠原代心肌細胞,48 h 后出現比較強的綠色熒光.病毒沒有使心肌細胞出現明顯的死亡現象且形態完整,可見毒性還算安全(圖4).
為檢測腺病毒是否能發揮應有的作用,將過濾后的病毒粗提液感染SD 大鼠原代心肌細胞.提取RNA和蛋白,然后進行RT-PCR,另外用兔HA 一抗和羊抗兔二抗做Western blot 檢測.RT-PCR 顯示,Txt5表達上調;Western blot的結果顯示,感染Txt5 過表達腺病毒的心肌細胞中檢測到與Txt5 融合表達的HA,而感染空載腺病毒Ad-GFP的對照組中未檢測到該條帶(圖4),說明該腺病毒成功介導Txt5 在SD 大鼠原代心肌細胞中的表達.
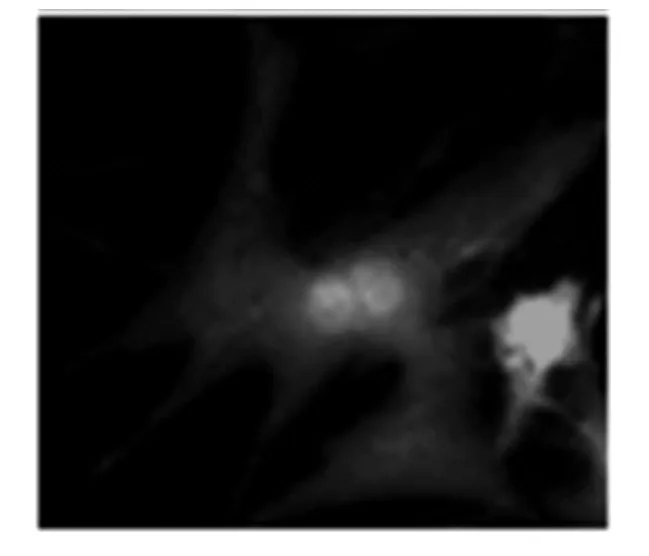
圖3 Txt5過表達腺病毒感染原代心肌細胞觀察結果

圖4 RT-RCR與免疫印跡法檢測腺病毒效果
3 討論
研究發現許多神經系統疾病相關基因的產物都具有LisH 結構域[6].這一結構域可能參與調節微管的動力,參與二聚化或與微管胞質動力蛋白重鏈相結合.Txt5 蛋白包含有LisH 結構域,這給我們啟示,是否Txt5和MAPK 信號通路有關[7].MAPK是一類重要的細胞信號轉導激酶,調節多種激素、神經遞質、細胞因子、生長因子對細胞生理活動的作用,該激酶作用底物多為例如Elk21、AP21[8]等的轉錄因子.
腺病毒在研究基因功能和治療方面的應用具有以下優勢:1)比較廣的宿主范圍,可以感染原代心肌細胞;2)能有效進行增殖,滴度較高,易于培養;3)基本上不插入宿主的基因組,不造突變[9-10].本文所用的腺病毒載體基本骨架中便含有GFP 序列,通過IRES 序列Txt5 基因與GFP 共表達,綠色熒光就會出現在感染了腺病毒的心肌細胞,視覺上就能判定感染效率.而RT-PCR 證明mTxt5 水平的上調,western blot發現HA表達,說明腺病毒效果良好,符合實驗要求.
本文研制小鼠Txt5 過表達腺病毒的操作簡單,周期較短,并且獲得的腺病毒滴度較高,毒性不強,能滿足體外實驗的要求.而現今市面上檢測Txt5的抗體比較少,本文研究設計中將Txt5 與HA 標簽融合表達,從而可以通過常用的HA 對應抗體檢測Txt5表達,給Txt5表達水平和位置的研究帶來了極大的幫助.
[1]SIMS RJ,WEIHE EK,ZHU L,et al.M-Bop,a repressor protein essential for cardiogenesis,interacts with skNAC,a heart-and muscle-specific transcription factor[J].The Journal of Biological Chemistry,2002,277(29):26524-26529.
[2]GOTTLIEB PD,PIERCE SA,SIMS RJ,et al.Bop encodes a muscle-restricted protein containing MYND and SET domains and is essential for cardiac differentiation and morphogenesis[J].Nature Genetics,2002,31 (1):25-32.
[3]唐文峴,王躍群,吳秀山,等.人類新基因RMND5A的克隆及功能研究[J].湖南師范大學自然科學學報,2008,31 (1):85-89.
[4]Ma H,Liu Y,Liu S,et al.Oral adeno-associated virus-TRAIL gene therapy suppresses human hepatocellular carcinoma growth in mice[J].Hepatology,2005,42(6):1355-1363.
[5]He TC,Zhou S,Da Costa LT,et al.A simplified system for generating recombinant adenovi-ruses[J].Proc Natl A cad Sci USA,1998,95 (5):2509-2514.
[6]KIM MH,COOPER DR,OLEKSY A,et al.The structure of the N2 terminal domain of the product of the lissencephaly gene Lis1 and its functional implications[J].Structure,2004,12 (6):987-98.
[7]UMEDA M,NISHITANI H,NISHIMOTO T,et al.A novel nuclear protein,Twa1,and Muslin comprise a complex with Ran BPM[J].Gene,2003,16 (303):47-54.
[8]DAVIS RJ.Signal transduction by the JNK group of MAP kinases[J].Cell,2000,103 (2):239-252.
[9]陳娜娜,向冬喜,鄭叢龍,等.腺病毒及其研究進展[J].大連醫科大學學報,2010 (5):586-590.
[10]邱峰,陳世知,陳虹,等.新型腺病毒載體的制備及其特性研究[J].重慶醫科大學學報,2006 (3):330-333.
