苜蓿-荒漠草地交錯帶步甲昆蟲多樣性
2015-12-03 05:56:00楊貴軍賈彥霞王新譜張大治
環境昆蟲學報 2015年3期
楊貴軍,賈彥霞,王新譜,3,張大治
(1.寧夏大學生命科學學院,銀川 750021;2.寧夏大學農學院,銀川 750021;3.寧夏大學西北退化生態系統恢復與重建省部共建教育部重點實驗室,銀川 750021)
生態交錯帶是兩個群落相連接的應力區,在這個應力區中,相鄰兩個群落的主要物種達到了它們的分布界限,是兩個相對均勻的相鄰群落相互過渡的突發轉換區域,具有其獨有的特性(孫儒泳,1992),交錯帶生境趨于異質復雜化,加之邊緣效應作用,導致群落交錯帶內生物多樣性變化(Harris,1988)。森林-草地交錯帶的地表昆蟲一直是生態研究和保護的重點(Davies and Margules,1998;Molnár et al.,2001;于曉東等,2006;Yu et al.2007,2009,2010;Brigic' et al.,2013;Barnes et al.,2014;)。許多研究表明,森林邊緣效應對甲蟲多樣性產生正面影響,森林邊緣內的物種多樣性會在短期內提高(Magura et al.,2001;Barbosa and Marquet,2002;Magura,2002;Mathe,2006;于曉東等,2006)。交錯帶地表昆蟲群落的多樣性還受森林斑塊面積、形成時間、邊緣植被構成以及季節,干擾等的影響(L?vei et al.,2006;Yu et al.,2007;Gaublomme et al.,2008)。
荒漠沙生植被受沙化環境和人為因素的影響,呈點、片、帶狀分布,構成了典型的沙化地破碎化荒漠植被景觀。荒漠昆蟲在維持荒漠生態系統的健康、保障荒漠生態的服務功能方面在一定程度上發揮著重要作用(黃人鑫,2005;婁巧哲等,2011),破碎化的荒漠景觀及其典型的交錯帶昆蟲多樣性逐步受到關注(Sanchez and Parmenter,2002;張大治等,2013;王巍巍等,2013)。在寧夏靈武荒漠草原-人工固沙檸條林和荒漠草原-固定半固定沙地兩種類型的交錯帶,邊緣的地表甲蟲多樣性特征高于相鄰生境,但邊緣效應存在差異(王巍巍等,2013)。賈彥霞和王新譜(2013)研究寧夏鹽池檸條-荒漠草地交錯帶表明,地表甲蟲的個體數量、多樣性和均勻度均以檸條灌叢內部最高,邊緣效應強度為正效應。
鹽池縣位于寧夏東部的屬鄂爾多斯臺地向黃土高原過渡地帶,沙生植被草地是鹽池縣草原的主體,具有典型的沙化地破碎化荒漠植被景觀。近年來,伴隨著人工沙漠治理工程和草畜產業開展,鹽池縣在沙化植被區墾植紫花苜蓿面積超過50 萬畝。為此我們選擇位于寧夏鹽池縣四墩子的紫花苜蓿-荒漠草地這一典型的交錯帶景觀,以經常作為環境變化指示生物的步甲昆蟲(婁巧哲等,2011)為研究對象,調查紫花苜蓿草地、荒漠草地及紫花苜蓿草地-荒漠草地交錯帶步甲昆蟲群落組成及季節動態,并分析影響步甲昆蟲分布的植被特征,探討苜蓿草地-荒漠草地交錯帶對步甲昆蟲多樣性的影響及其群落組成演替,以期為荒漠植被恢復與重建以及維持荒漠草原生態系統平衡提供參考。
1 研究地區與研究方法
1.1 研究地概況
研究區位于寧夏東部的鹽池縣(N37°04'-38°10',E106°30'-107°41'),該地北與毛烏素沙漠相連,南靠黃土高原,處于典型中溫帶大陸性氣候帶,年均氣溫8.1℃,年降水量250-350 mm,植被類型是從干草原向荒漠草原的過渡地帶,主要有灌叢、草原、草甸、沙地植被和荒漠植被,其中灌叢、草原、沙地植被數量較大,分布也廣。草原群落中常見植物種類以旱生和中旱生類型為主。
研究樣地為人工種植紫花苜蓿Medicago sativa草地和荒漠草地的交錯帶。其中苜蓿草地每年夏季和秋季各刈割一次,并有施肥、除草等農藝措施,連片面積約4.43 km2,苜蓿行間距約2 m,株距約0.4 m,苜蓿行間主要有稗草Echinochloa crusgalli、沙生冰草Agropyron desertorum、豬毛蒿Artemisia scoparia、櫛葉蒿Neopallasia pectinata 和牻牛兒苗Erodium stephanianum,夏季植物高度約26 cm,蓋度約55%。荒漠草地為沒有開墾過的原始荒漠草本植物帶,主要植物有短花針茅Stipa breviflora、鐵桿蒿Artemisia sacrorum、豬毛蒿、櫛葉蒿、二裂委陵菜Potentilla bifurca、糙隱子草Cleistogenes squarrosa、達烏里胡枝子 Lespedeza davurica、匍根駱駝蓬Peganum nigellastrum 和沙生冰草等,夏季植物高度約21 cm,蓋度約69%。
1.2 調查方法
本研究以巴氏罐誘法進行標本采集(Yu et al.,2007,2009),采用400 mL 塑料杯(高9 cm,口徑7.5 cm)作為容器,杯壁上方1/4 處(杯口下方約2.5 cm)打一小孔(直徑約0.5 cm),以防雨水過多使標本流失;引誘劑為醋、糖、酒精和水的混合物,重量比為2∶1∶1∶20,每個誘杯內放引誘劑40-60 mL。采集到的標本存放在70%的酒精內。采樣時間為2012年6月到10月,每月采樣1 次,每次放置誘杯時間3 d。
邊緣效應的研究參考(Meiners and Pickett,1999;Heliola et al.,2001)方法,以距離梯度形式研究苜蓿草地內部、苜蓿-荒漠草地交錯帶和荒漠草地內部步甲群落的差異。在設計樣點時,每條樣帶以苜蓿草地邊緣為中心,分別向荒漠草地和苜蓿草地延伸100 m,以每隔25 m 的距離設置樣點,這樣每個研究樣帶包括9個研究樣點,苜蓿草地邊緣1個,苜蓿草地內和荒漠草地內各4個(距苜蓿草地邊緣的距離分別為25 m、50 m、75 m 和100 m);每個樣點由5個誘杯組成,間距1-2 m,呈五點排列,平行于苜蓿草地邊緣,每個樣帶包括45個誘杯(圖1)。
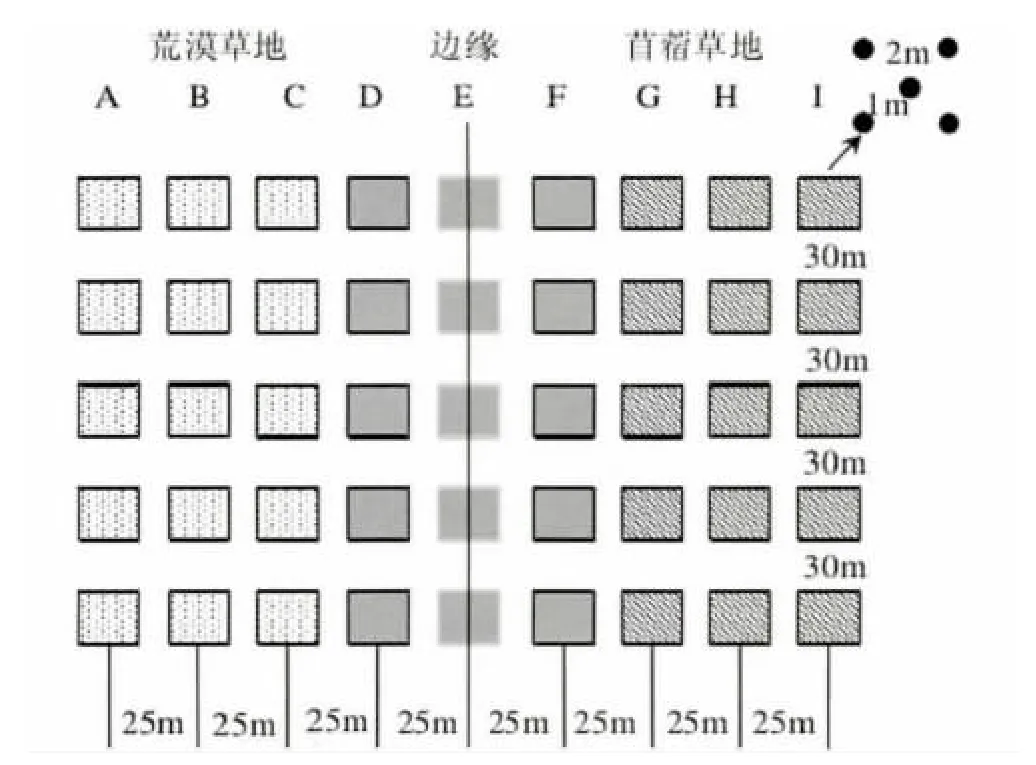
圖1 研究樣地設置Fig.1 Map of the study site showing actual positions of each plot
為了研究植被特征對步甲分布的影響,我們在實驗中期(2012年8月)調查了9個研究樣帶(以樣點中央誘杯為中心,直徑2 m 的范圍)的植被種類、密度、高度和蓋度等特征。
1.3 數據處理與分析
以步甲昆蟲的物種數、個體數量、多樣性、均勻度及優勢度為對象,將45個研究樣點(5個研究樣帶,每個樣帶包括9個研究樣點)根據與苜蓿草地邊緣的距離聚成3 組:苜蓿草地生境包括深入到苜蓿草地內部的3個距離組(50 m、75 m以及100 m),荒漠草地生境包括深入到荒漠草地內部的3個距離組(50 m、75 m 以及100 m),交錯帶包括其他3個距離組(邊緣、深入苜蓿草地生境25 m,深入荒漠草地生境25 m)。
以“活動密度”反映不同樣地步甲昆蟲的數量,其含義為樣地中荒漠草地、交錯帶和苜蓿草地各75個巴氏罐3 d 所捕獲的甲蟲個體數。
物種多樣性(H')分析采用Shannon-Wiener多樣性指數,計算公式為:H'=-∑PilnPi,其中Pi=Ni/N,Pi是第i種個體數占總個體數的比率,Ni是第i種的個體數,N 是總個體數;均勻度(J)分析采用Pielou 均勻度指數,計算公式為:J=H'/lnS,S 是物種數;優勢度(C)分析采用Simpson 優勢度指數,計算公式為:C=∑(ni/N)2(趙志模和郭依泉,1990;馬克平和劉玉明,1994)。
利用SPSS 16.0 對步甲和環境數據的顯著性檢驗:對服從正態分布或轉換后服從正態分布的數據,采用單因素方差以及Tukey 多重比較分析荒漠草地、交錯帶以及苜蓿草地間步甲蟲科的物種數、活動密度、多樣性、均勻度以及優勢度的差異。采用多元線性逐步回歸(stepwise)檢驗步甲昆蟲的豐富度、活動密度、多樣性、均勻度以及優勢度與植被特征之間關系。
基于Bray-Curtis 相似性系數,使用統計軟件PAST 進行主坐標分析(PCoA,principal coordinate ajlalysis)排序(Pielou,1984;Ludwig & Reynolds,1988),對苜蓿草地內部、邊緣以及荒漠草地間步甲甲蟲群落組成進行分析。
2 結果與分析
2.1 步甲昆蟲群落種類組成
共采集步甲昆蟲1944 頭,分屬11 屬22種。其中直角通緣步甲Pterostichus gebleri1320 頭,占總個體數的67.90%,為該地區的優勢種類。蒙古偽葬步甲Pseudotaphoxnus Mongolicus、短翅偽葬步甲P.brevipennis、谷婪步甲Harpalus calceatus、黃鞘婪步甲H.pallidipennis、直角婪步甲H.corporosus、徑婪步甲H.salinus、赤胸梳步甲Dolichus halensis、雙斑猛步甲 Cymindis binotata、皮步甲Corsyra fusula 和星斑虎甲Cicindela raleea個體數量在1%-10%之間,為該地區常見類群。其他步甲體數量少于1%,為該地區比較不常見的類群。
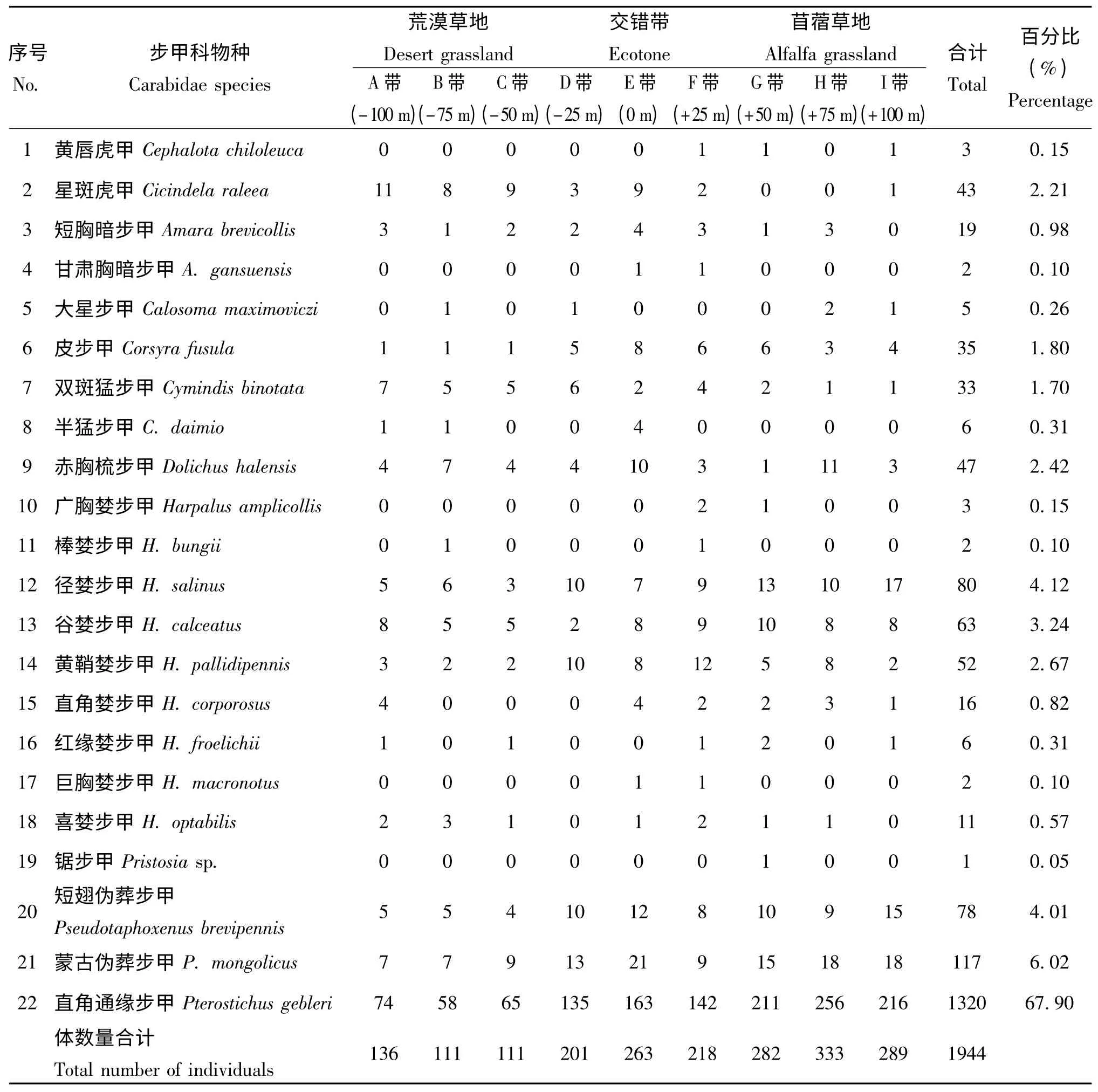
表1 不同樣帶步甲昆蟲的組成和數量分布Table 1 Species and individuals of Carabidae in different transect
對距邊緣不同距離的步甲昆蟲群落豐富度和個體數量進行比較,結果見圖2。步甲昆蟲個體數量沿邊緣向苜蓿草地升高,而向荒漠草地內部降低,總體上以苜蓿草地內部的個體數量高于荒漠草地內部,但各樣帶差異不顯著。步甲豐富度沿邊緣分別向荒漠草地內部和荒漠草地降低,且荒漠草地的步甲豐富度略高于苜蓿草地內部,但各樣帶差異亦不顯著。
優勢種直角通緣步甲的個體數量沿荒漠草地、交錯帶到苜蓿草地逐漸增加,偏好于苜蓿草地生境,但樣帶間差異不顯著(圖3)。

圖2 不同邊緣距離的步甲昆蟲群落個體數和豐富度Fig.2 Abundance and individuals of ground beetles community in different transect

圖3 優勢種在不同樣帶的數量分布Fig.3 Quantitative distribution of dominant species of ground beetles community in different transect
2.2 步甲昆蟲群落多樣性及邊緣效應分析
以活動密度表征3種生境步甲昆蟲的個體數量變化,表現為從苜蓿草地、交錯帶到荒漠草地逐漸降低,3種生境間有極顯著差異(F=42.05,P=0.0001)(見表2)。交錯帶的步甲群落的物種數高于荒漠草地和苜蓿草地,但沒有達到顯著差異。步甲群落Shannon-Wiener 多樣性沿荒漠草地、邊緣到苜蓿草地依次降低,差異顯著(F=13.105,P=0.001)。均勻度的變化趨勢與Shannon-Wiener 多樣性一致,且呈極顯著正相關(r=0.917,P=0.001),不同生境間差異顯著(F=8.551,P=0.0049)。優勢度的變化趨勢分別與均勻度(r=-0.964,P<0.0001)和Shannon-Wiener 多樣性(r=-0.989,P<0.0001)極負相關顯著,苜蓿草地步甲群落優勢度最高,不同生境間差異顯著(F=10.854,P=0.002)。
從基于步甲昆蟲群落組成為屬性的PcoA 排序圖看(圖4,第1 和第2 軸分別解釋了92.18%和2.03%的變異),荒漠草地內部和苜蓿草地內部沒有重疊,交錯帶分別與荒漠草地、苜蓿草地都有部分重疊。這一結果說明,荒漠草地的步甲昆蟲群落與苜蓿草地內部的群落組成已經有了明顯分化。
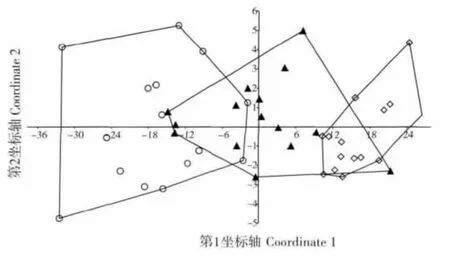
圖4 以步甲群落組成為屬性的對荒漠草地、交錯帶及苜蓿草地生境的主坐標排序圖Fig.4 Ordination plot(PCoA,Principal Coordinate Analysis)of the range of pitfall catches per trap location indesert grassland,alfalfa grassland-desert grassland ecotone and Alfalfa grassland

表2 不同生境的步甲昆蟲群落多樣性特征Table 2 Comparison of ground beetles community diversity indices among different transect
邊緣效應強度是對邊緣效應的強弱進行測量的定量指標,據王伯蓀等(1986)所組建的測度模型來計算,EH'=1.06>1,EC=0.93(變換為倒數為1.07>1),可以看出這種邊緣類型呈正的邊緣效應作用。
2.3 步甲昆蟲群落多樣性時間變化
步甲昆蟲群落多樣性時間變化見圖5。在整個調查期,交錯帶的步甲昆蟲物種數比荒漠草地和苜蓿草地內部高(表2),但從不同月份調查看,基本上呈從荒漠草地、交錯帶到苜蓿草地逐漸降低的趨勢;3種生境步甲群落物種數隨調查時間變化趨勢一致,6月和8月的物種數最多,10月最低。活動密度的月份變化與整個調查期的總趨勢相符,從苜蓿草地、交錯帶到荒漠草地依次逐漸遞減;3種生境步甲活動密度變化趨勢一致,在8月最高。物種多樣性月份變化在調查前、中期與整個調查期的總和趨勢一致,而后期則不同;荒漠草地步甲物種多樣性隨月份逐漸降低,交錯帶和苜蓿草地的變化趨勢一致,在調查的前、中期降低,在后期的9月份出現小的高峰。均勻度的月份變化與整個調查期的總和趨勢不同;荒漠草地和苜蓿草地月份變化趨勢一致,在前、后期較高,在中期最低,而交錯帶則表現為在6月和9月最高。
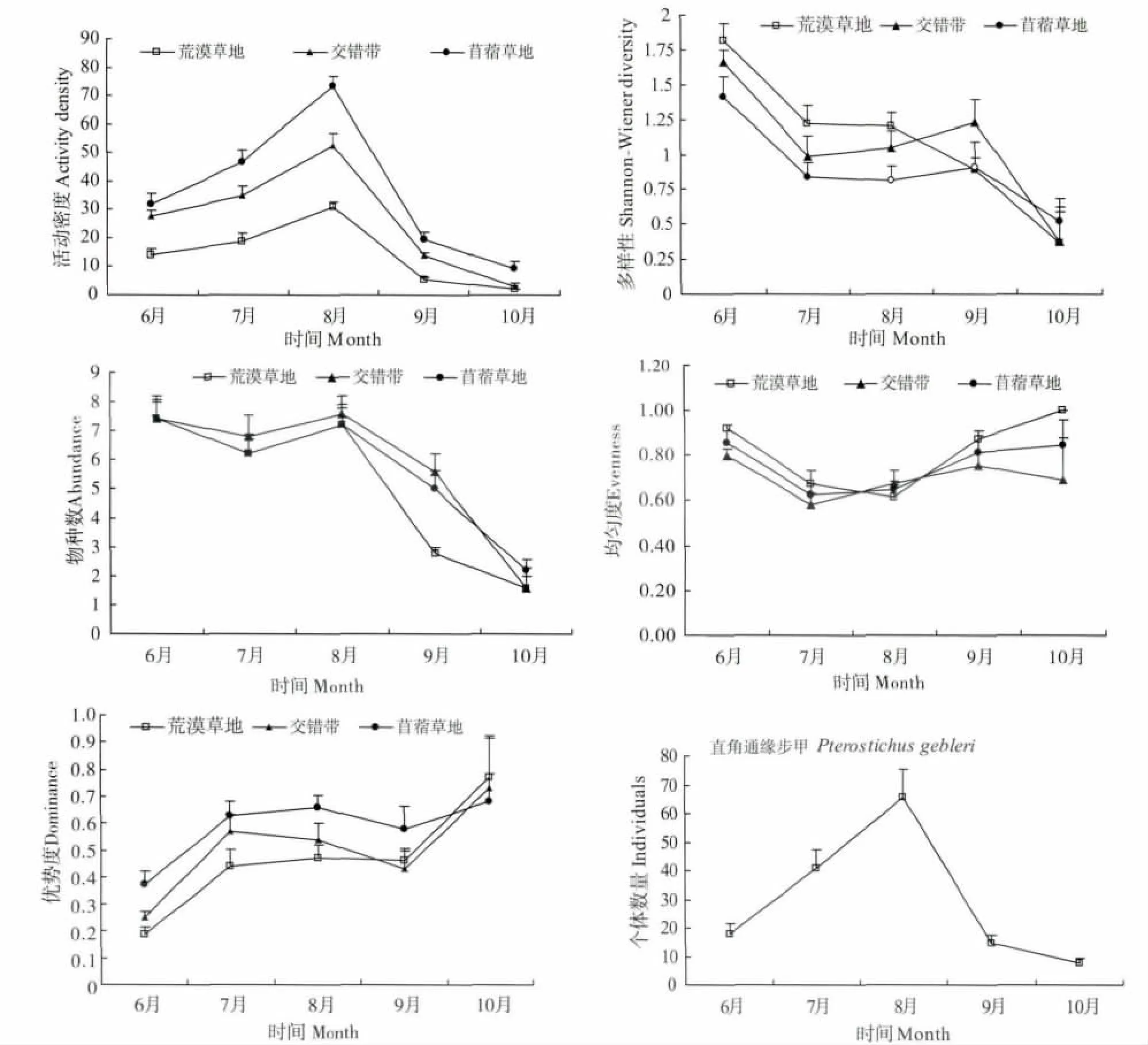
圖5 荒漠草地、交錯帶和苜蓿草地間步甲昆蟲活動密度、多樣性、物種數、均勻度和優勢度以及優勢優勢種的時間變化Fig.5 Monthly changes ofactivity density,diversity,evenness,richness and dominance of ground beetles in desert grassland,alfalfa-desert grassland ecotone and alfalfa grassland and number of individuals of dominant species
優勢種直角通緣步甲不同月份的個體數量差異極顯著(F=17.418,P=0.0001),8月份最多,10月份最少。
2.4 昆蟲群落多樣性與植被特征的相關性
由表3 可以看出,不同研究樣帶的植物Shannon-Wiener 多樣性差異極顯著(F=7.035,P=0.0003),沿荒漠草地(A、B、C 帶)、交錯帶(D、E、F 帶)到苜蓿草地(G、H、I 帶)降低。植被密度沿荒漠草地、交錯帶到苜蓿草地升高,不同樣帶間差異不顯著。植被蓋度變化總趨勢與植物Shannon-Wiener 多樣性一致,但不同樣帶間差異不顯著。植被高度從苜蓿草地邊緣分別向荒漠草地、苜蓿草地增高,不同樣帶間差異亦不顯著。
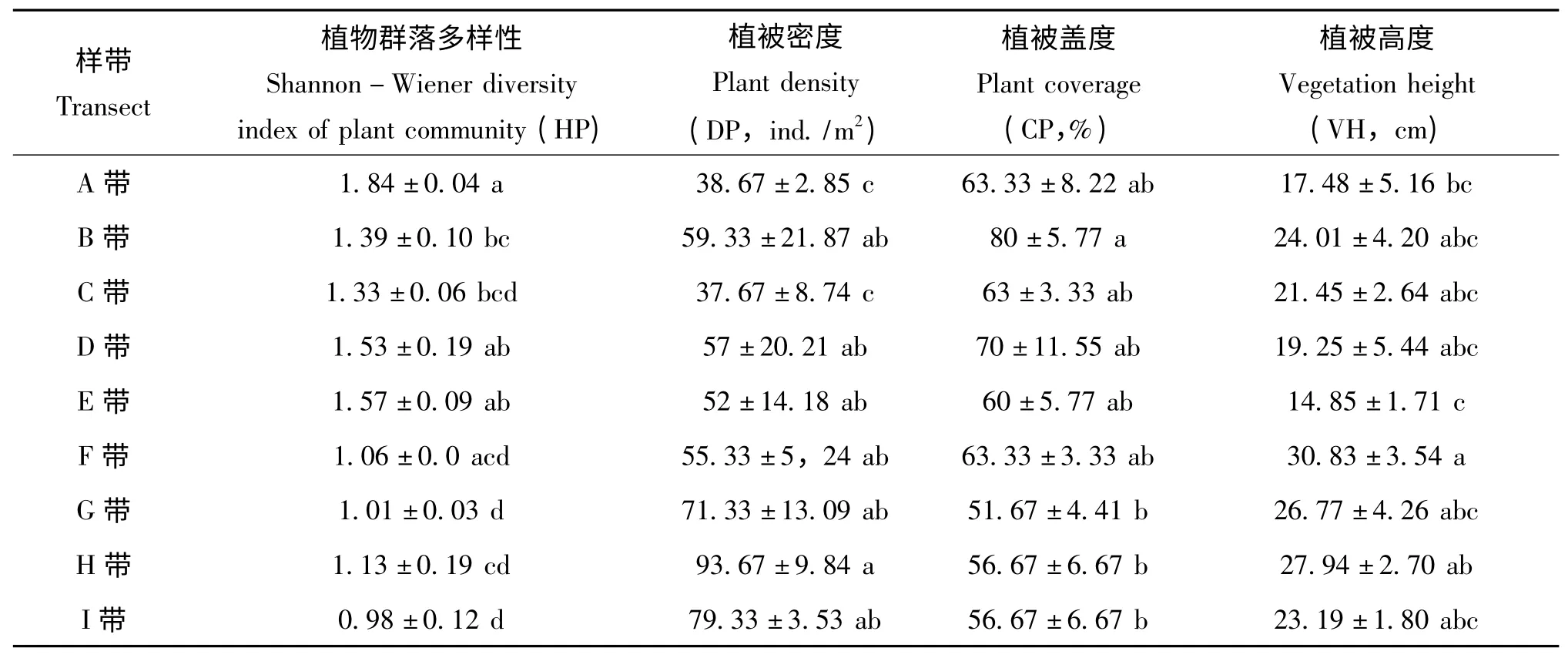
表3 不同研究樣帶的植被特征Table 3 Vegetation characteristicsof different transect
對步甲昆蟲物種數、活動密度、多樣性、均勻度以及優勢度與這4種植被特征進行多元回歸檢驗,結果發現植被密度是影響步甲Shannon-Wiener 多樣性(r2=0.711,F=17.205,P=0.004)、均勻度(r2=0.649,F=12.971,P=0.009)和優勢度(r2=0.691,F=15.676,P=0.005)的決定因素;植被密度、植被蓋度是影響步甲活動密度(r2=0.870,F=20.061,P=0.002)決定因素。植被特征對步甲的物種數沒有顯著的回歸關系。對優勢種直角通緣步甲的回歸分析顯示,其活動密度由植被密度和植被蓋度(r2=0.939,F=46.293,P<0.0001)決定。

表4 植被特征與步甲昆蟲活動密度、物種數、多樣性、均勻度及優勢度間回歸分析Table 4 Regression analysis of vegetation characteristics andactivity density,richness,diversity,evenness and dominance e of ground beetles
3 結論與討論
本研究表明,苜蓿-荒漠草地交錯帶步甲物種組成表現為邊緣>荒漠草地>苜蓿草地,在不同生境差異不顯著,幾乎所有的物種(除鋸步甲外)在交錯帶都被采集到,說明交錯帶的物理條件沒有對相鄰生境的種類組成造成不利影響,回歸分析也說明,植被特征對步甲的物種組成也沒有顯著的回歸關系,植被群落多樣性對步甲的多樣性并沒有明顯影響。地表甲蟲的組成和分布受棲息地的植被類型、土壤基質和地勢方位等環境因素的影響(Antvogel and Bonn,2001;婁巧哲等,2011;Li,et al.,2013),本文研究對象步甲在功能類型上基本全屬于捕食性昆蟲,因此植被組成等環境因素對其分布的影響是間接地。
苜蓿-荒漠草地交錯帶步甲個體數量沿苜蓿草地、邊緣到荒漠草地依次降低,差異極顯著。回歸分析說明,植被密度是影響步甲的Shannon-Wiener 多樣性和優勢度,植被密度和植被蓋度是影響整個步甲群落和優勢種直角通緣步甲活動密度的決定因素。苜蓿草地的植被密度較荒漠草地高,所以聚集了較多的食草和腐食性昆蟲,為捕食性步甲提供了豐富的食物資源,同時苜蓿草地相對溫和的微環境可為步甲昆蟲提供安全的產卵場所,所以苜蓿草地內部步甲的活動密度較大。已有關于灌叢草地的研究也有類似結果,荒漠灌叢密度和蓋度影響植被的郁閉度,灌木覆蓋的“頂篷”效應改善了灌叢周圍的微環境(Li et al.,2008;Wang et al.,2011),對地表昆蟲的豐富度和多樣性具有促進作用(Stapp,1997;Titus,et al.,2002;Li et al.,2005;Mazía et al.,2006;Liu et al.,2012)。
步甲群落Shannon-Wiener 多樣性和均勻度沿荒漠草地、邊緣到苜蓿草地依次降低,而優勢度的變化趨勢與均勻度和Shannon-Wiener 多樣性相反,說明苜蓿草地內部與荒漠草地內部的步甲群落特征差異比較明顯。主成分PCoA 排序也表明苜蓿草地內部與荒漠草地內部的群落組成沒有重疊,苜蓿草地內部的步甲群落與荒漠草地有明顯的分化,邊緣分別與苜蓿草地和荒漠草地有部分重疊,具有這兩個生境步甲昆蟲群落的特征,這一結果與前人關于森林邊緣的步甲研究相似(Heli?l?,et al.,2001;Magura,2002;于曉東等,2006),交錯帶的物種組成來相鄰生境的擴散。
邊緣效應強度分析EH'值大于1,說明這種邊緣類型呈正的邊緣效應作用。一般情況下,棲境邊緣地帶常會有新的微觀環境(包括生物和非生物兩方面),可能導致有高的生物多樣性(Murcia,1995)。已有關于森林邊緣的地表甲蟲的研究,一般表現為正的邊緣效應(Magura et al.,2001;Magura,2002)。王巍巍等(2013)研究荒漠草原-人工固沙檸條林和荒漠草原-固定半固定沙地兩種類型的交錯帶地表甲蟲群落,前者表現弱的負邊緣效應,賈彥霞和王新譜(2013)研究的檸條-荒漠草地交錯帶地表昆蟲群落邊緣效應強度為正效應,這種差異可能與邊緣的植被類型、地勢、邊緣區域發育時間以及某些甲蟲類群對特定環境的喜好性和敏感性有關。本文研究的交錯帶類型與上述研究結果不完全一致,首先與交錯帶類型差異有關,其次與研究對象有關,本文中的研究對象為步甲,在功能群和擴散能力等方面有較大差異,另外,本研究樣地苜蓿草地每年夏季和秋季各刈割一次,并有施肥、除草等農藝措施,所以人為干擾強度的差異也會影響交錯帶地表昆蟲群落多樣性水平。
References)
Antvogel H,Bonn A.Environmental parameters and micro-spatial distribution of insects:a case study of carabids in an alluvial forest[J].Ecography,2001,24(4):470-482.
Barbosa O,Marquet PA.Effects of forest fragmentation on the beetle assemblage at the relict forest of Fray Jorge Chile[J].Oecologia,2002,132(2):296-306.
Barnes AD,Emberson RM,Chapman HM,et al.Matrix habitat restoration alters dung beetle species responses across tropical forest edges[J].Biological Conservation,2014,170:28-37.
Gaublomme E,Hendrickx F,Dhuyvetter H,et al.The effects of forest patch size and matrix type on changes in carabid beetle assemblages in an urbanized landscape[J].Biological Conservation,2008,141(10):2585-2596.
He Q,Wang XP,Yang GJ.Species diversity of carabid beetles in desert-steppe in Yanchi of Ningxia,China[J].Acta Ecologica Sinica,2011,31(4):923-932.[賀奇,王新譜,楊貴軍.寧夏鹽池荒漠草原步甲物種多樣性[J].生態學報,2011,31(4):923-932]
Heli?l? J,Koivula M,Niemel? J.Distribution of carabid beetles(Coleoptera,Carabidae)across a boreal forest-clearcut ecotone[J].Conservation Biology,2001,15(2):370-377.
Huang RX.The Fauna of the Desert Insects of Xinjiang and its Formation and Evolution[M].Urumqi:Xinjiang Science and Technology Publishing House,2005.[黃人鑫.新疆荒漠昆蟲區系及其形成與演變[M].烏魯木齊:新疆科學技術出版社,2005]
Jia YX,Wang XP.Diversity of ground-dwelling beetles at the edge of grassland and artificial Caragana fields in Sidunzi,Yanchi County,Northwest China[J].Acta Prataculturae Sinica,2013,22(5):345-350.[賈彥霞,王新譜.鹽池四墩子人工檸條帶-草地群落交錯帶地表甲蟲多樣性研究[J].草業學報,2013,22(5):345-350]
Li FR,Liu JL,Liu CA,et al.Shrubs and species identity effects on the distribution and diversity of ground-dwelling arthropods in a Gobi desert[J].Journal of Insect Conservation,2013,17(2):319-331.
Li FR,Wang T,Zhang AS,et al.Wind-dispersed seed deposition patterns and seedling recruitment of Artemisia halodendron in a moving sandy land[J].Annals of Botany,2005,96(1):69-80.
Li XY,Liu LY,Gao SY,et al.Stemflow in three shrubs and its effect on soil water enhancement in semiarid loess region of China[J].Agricultural and Forest Meteorology,2008,148(10):1501-1507.
Liu JL,Li FR,Liu CA,et al.Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem[J].Biodiversity and Conservation,2012,21:2601-2619.
Lou QZ,Xu YC,Ma JH,et al.Diversity of ground-dwelling beetles within the southern Gurbantunggut Desert and its relationship with environmental factors[J].Biodiversity Science,2011,19(4):441-452.[婁巧哲,徐養誠,馬吉宏,等.古爾班通古特沙漠南緣地表甲蟲物種多樣性及其與環境的關系[J].生物多樣性,2011,19(4):441-452.]
L?vei GL,Magura T,Tóthmérész B,et al.The influence of matrix and edges on species richness patterns of ground beetles(Coleoptera:Carabidae)in habitat islands[J].Global Ecology and Biogeography,2006,15(3):283-289.
Ludwig JA,Reynolds JF.Statistical Ecology:A Primer on Method and Computing[M].New York:John Wiley and Sons,1988.
Ma KP,Liu YM.Measurement of biotic community diversity I:α diversity(Part 2)[J].Biodiversity Science,1994,2(4):231-239.[馬克平,劉玉明.生物多樣性測度方法:α-多樣性[J].生物多樣性,1994,2(4):231-239.]
Magura T,Tóthmérész B,Molnár T.Forest edge and diversity:carabids along forest-grassland transects[J].Biodiversity and Conservation,2001,10(2):287-300.
Magura T.Carabids and forest edge:spatial pattern and edge effect[J].Forest Ecology and Management,2002,157(1):23-37.
Mathe I.Forest edge and carabid diversity in a Carpathian beech forest[J].Community Ecology,2006,7(1):91-97.
Mazía CN,Chaneton EJ,Kitzberger T.Small-scale habitat use and assemblage structure of 6 ground-dwelling beetles in a Patagonian shrub steppe[J].Journal of Arid Environments,2006,67:177-194.
Meiners SJ,Pickett STA.Changes in community and population responses across a forest field gradient[J].Ecography,1999,22(3):261-267.
Murcia C.Edge effects in fragmented forests:implications for conservation[J].Trends in Ecology and Evolution,1995,10(2):58-62.
Pielou EC.TheInterpretation of Ecological Data:A Primer on Classification and Ordination[M].New York:John Wiley and Sons,1984.
Sanchez BC,Parmenter RR.Patterns of shrub-dwelling arthropod diversity across a desert shrubland-grassland ecotone:a test ofisland biogeographic theory[J].Journal of Arid Environments,2002,50(2):247-265.
Stapp P.Microhabitat use and community structure of darkling beetles(Coleoptera:Tenebrionidae)in shortgrass prairie:effects of season shrub and soil type[J].American Midland Naturalist,1997:298-311.
Titus JH,Nowak RS,Smith SD.Soil resource heterogeneity in the Mojave Desert[J].Journal of Arid Environments,2002,52(3):269-292.
Wang WW,He DH,Zhang DZ.Edge effects on ground-dwelling beetle communities in a desert landscape[J].Chinese Journal of Applied Entomology,2013,50(5):1383-1391.[王巍巍,賀達漢,張大治.荒漠景觀地表甲蟲群落邊緣效應研究[J].應用昆蟲學報,2013,50(5):1383-1391]
Wang XP,Wang ZN,Berndtsson R,et al.Desert shrub stemflow and its significance in soil moisture replenishment[J].Hydrology and Earth System Sciences,2011,15:561-567.
Yu XD,Luo TH,Zhou HZ,et al.Distribution of carabid beetles(Coleoptera:Carabidae)across a forest-grassland ecotone in Southwestern China[J].Environmental Entomology,2007,36(2):348-355.
Yu XD,Luo TH,Zhou HZ,et al.Influence of edge effect on diversity of ground-dwelling beetles across forest-grassland ecotone in Wolong Natural Reserve,Southwest China[J].Acta Entomologica Sinica,2006,49(2):277-286.[于曉東,羅天宏,周紅章,等.邊緣效應對臥龍自然保護區森林-草地群落交錯帶地表甲蟲多樣性的影響[J].昆蟲學報,2006,49(2):277-286]
Yu XD,Luo TH,Zhou HZ.Distribution of carabid beetles(Coleoptera:Carabidae)across ecotones between regenerating and mature forests in southwestern China[J].Environmental Entomology,2009,38(4):1053-1060.
Yu XD,Luo TH,Zhou HZ.Distribution of ground-dwelling beetle assemblages(Coleoptera)across ecotones between Natural Oak Forests and Mature Pine Plantations in North China[J].Journal of Insect Conservation,2010,14(6):617-626.
Yu XD,Luo TH,Zhou HZ.Effects of forestry practices and forest fragmentation on beetle diversity and conservation strategies[J].Acta Entomologica Sinica,2006,49(1):126-136.[于曉東,羅天宏,周紅章.林業活動和森林片斷化對甲蟲多樣性的影響及保護對策[J].昆蟲學報,2006,49(1):126-136]
Zhang DZ,Ma Y,Li YC,et al.Effects of Caragana scrubland fragmentation on fine-scale diversity of ground-dwelling beetles in a desert landscape[J].Chinese Journal of Applied Entomology,2013,50(4):934-941.[張大治,馬艷,李岳誠,等.小尺度下檸條林破碎化生境對地表甲蟲多樣性的影響[J].應用昆蟲學報,2013,50(4):934-941]
Zhao ZM,Guo YQ.Principle and Methods of Community Ecology[M].Chongqing:Publishing House of Scientific and Technical Documentation,1990,147-279.[趙志模,郭依泉.群落生態學原理與方法[M].重慶:文獻和科學技術出版社,1990,147-279]
