荒漠竹節蟲的形態及生物學特性研究
2015-12-03 05:55:56陳新燕馬東建范兆田
環境昆蟲學報 2015年2期
陳新燕,史 雯,馬東建,范兆田,吳 衛*
(1.新疆大學生命科學與技術學院,烏魯木齊 830046;2.新疆大學理化測試中心,烏魯木齊 830046)
竹節蟲目昆蟲因其超高的擬態而廣為人知,目前有記錄的3 科500 屬3000種,均為植食性物種,多分布于炎熱潮濕的熱帶、亞熱帶地區,鮮少在地處溫帶內陸干旱區的中國新疆鮮有發現,目前發現該種為唯一一種在中國新疆的干旱、半干旱荒漠中分布的竹節蟲目昆蟲;其分布區域晝夜溫差大,夏季極其干旱、炎熱而冬季則寒冷而漫長,因此獲名荒漠竹節蟲(黃仁鑫等,2005)。國外對該種昆蟲僅有少量報道(БЕЙ-БИЕНКО,1964;Hennemann et al.,2008),在生物學特性方面尚無文獻記載。本文作者在2006年4月至2013年10月對荒漠昆蟲的野外考察期間,觀察并采集了一部分在新疆干旱荒漠環境中生存的荒漠竹節蟲,并分別于2006年5月至2008年12月,2013年4月至12月在室內進行人工飼養,對其形態學和生物學特性有了進一步的了解,現記述如下。
1 材料與方法
1.1 研究對象
荒漠竹節蟲隸屬于 Phasmatodea 目Diapheromeridae 科Sceptrophasma 屬(Brock & Seow-Choen,2000),實驗材料為2006年6月至2013年10月期間,在烏魯木齊市郊及準噶爾盆地荒漠南緣采集的所有成體、卵和幼體。
1.2 調查地點
1.2.1 雅瑪里克山
地理位置為 N43° 45' 44″-43° 47' 25″,E87°33' 16″-87° 34'56″,位于烏魯木齊市西側,約呈南北走向,平均海拔800 m,最高點青年峰海拔1391 m,山體地表面積約40 km2,長度約16 km。地質結構為油頁巖、片麻巖構成;植被以荒漠植被為主,植物群落構成較簡單,春秋季較濕潤,夏季干旱,冬季漫長而寒冷。1983-1996年統計表明,年均溫7.5℃,極端最高溫37.7℃,極端最低溫度-19.1℃,年降水量為266.9 mm,年蒸發量2731 mm,是總降水量的10.2 倍;平均相對濕度58%,屬中溫帶荒漠氣候。由于近年來的荒山綠化和封山育林,使荒山植被除了荒漠外,還有荒漠草原、真草原、草甸、沼澤的出現(葉衛英,2005)。
1.2.2 準噶爾盆地南緣
本地區代表著歐亞大陸腹地、溫帶內陸荒漠區。地帶性植被為典型的荒漠植被,如梭梭Haloxylon ammodendron、沙拐棗 Calligonum mongolicum、絹蒿Seriphidium korovinii 等;氣候類型為典型的大陸性干旱氣候,年均溫6.6℃,最高氣溫42.6℃,最低氣溫-41.6℃,年降水量164 mm,年蒸發量2000 mm 左右。本研究選擇102 團荒漠(E87°81'09″N44°36'49″)和阜康荒漠生態試驗站(E87°87'92″ N44°37'71″)2個區域進行調查。生境較為相似,但后者區域內植被保護較好,且有人工植被及試驗地分布,是研究的理想選地。
1.3 研究方法
1.3.1 野外觀察
分別在上述調查地點對荒漠竹節蟲成蟲及幼蟲的生態、行為、食性等進行觀察,于卵期在其棲息地的寄主植物上尋找自然產出的卵粒,標記并定期觀察,以確定其孵化時間。
1.3.2 室內飼養
除了野外觀察,還在室內進行人工飼養,方法如下:人工移植不同棲息地的寄主植物絹蒿至室內,與當地采集到的4 齡或5 齡若蟲一同罩在自制的紗籠中,歷經蛻皮、羽化、產卵、孵化等階段;五齡若蟲單獨飼養,以觀察其繁殖特性,用Olympus μ-mini D、尼康D50 和Olympus E-410等數碼相機進行戶外及室內影像記錄;用尼康SMZ-800 解剖顯微鏡和Lnoe-1403VT 掃描電子顯微鏡進行成體及卵的顯微結構觀察;使用HTC-1電子溫濕度計測量溫、濕度數據,標本保存于新疆大學生命科學與技術學院。
2 結果與分析
2.1 成蟲形態特征
身體細長、纖弱,樹枝狀,灰土色,表面光裸,完全無翅(圖1 A)。
頭部呈卵圓形,觸角粗短,明顯分為16 節,其上有稀疏的感覺毛,長度不超過前足腿節的1/3;柄節背腹方向扁平,長而寬大,背面有一縱向脊;梗節短,寬為長的1.2 倍;鞭節14 節,略扁平,第一鞭小節的長度是寬度的2.6 倍,第三和第五節鞭節的長和寬幾乎相等,末節端部圓,長是寬的2.2 倍;其余鞭節寬大于長。復眼橢圓而突出,復眼間具成對縱行的結節,無單眼。唇基前緣平直,額略凹陷。顳邊緣隆起,長度是復眼縱向直徑的5 倍;后頭平,向后變窄(圖1 B)。胸部細長較扁,無翅,前胸為前端略窄的梯形,比頭部稍窄,中胸是胸部最長的一節,長為寬的6 倍,中胸、后胸背板中央有一縱棱,與兩側邊的脊平行,前胸、中胸和后胸比例為1.0∶6.2∶4.6。足細長,前足股節基部彎曲,稍扁。中、后足脛節下方中棱達脛節端部。腿節∶脛節∶跗節長度比例為:前足:4.1∶4.3∶1.0;中足:4.0∶4.3∶1.0;后足:4.4∶5.0∶1.0。
腹部扁平,明顯分為10 節,第1 節較短,第2-7 節逐漸延長,第4-5 節最寬。雌體腹部第7節腹板末端具尖突,腹瓣光滑,尖矛形,不超過臀節基部;臀節有時凹陷。肛上板有3 條縱棱,兩側的不甚明顯,第8、9 節的背板中棱較平坦;尾須不分節(圖1:C,D,E)。
體長:6.2-7.6 mm(雌體)。
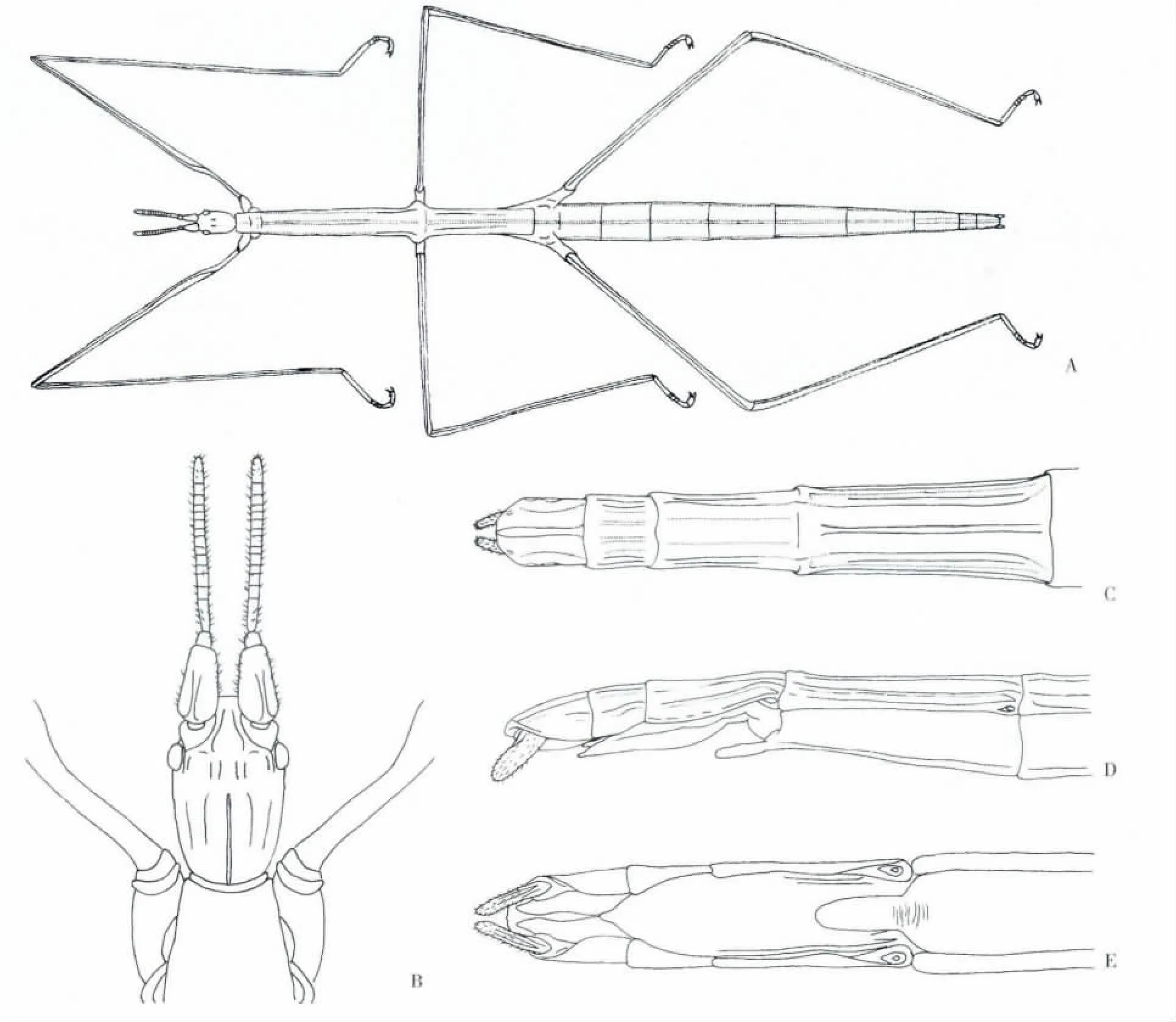
圖1 A,荒漠竹節蟲成蟲)♀;B,頭部背面觀(♀);C,腹部第7-10 體節背面觀(♀);D,腹部第7-10 體節側面觀(♀);E,腹部第8-10 體節腹面觀(♀)Fig.1 A,Sceptrophasma bituberculatum ♀;B,Dorsal view of head(♀);C,Dorsal view of VII-X abdominal segments(♀);D,Lateral view of VII-X abdominal segments(♀);E,Ventral view of VIII-X abdominal segments(♀)
2.2 分布及棲息地環境
據記載本種分布于哈薩克斯坦,土庫曼斯坦和中國新疆(БЕЙ-БИЕНКО,1964;蔡保靈,1987;Hennemann et al.,2008)。1980年至今的野外調查期間,僅在新疆伊犁河谷、天山低山山麓、準噶爾盆地荒漠僅發現了幾次。它們多生活于有枯枝的干旱、半干旱的荒漠植叢中(圖2),以其中一種絹蒿屬Serphidium 植物為寄主(野外采集時,荒漠竹節蟲的卵、若蟲和成蟲在其上均有發現)。據采集者稱,白茨屬Nitraria 植物亦可能是其寄主植物,初次室內喂養時,禾本科植物芨芨草Achnatherum splendens 也可被其取食。
2.3 產卵及卵的外形結構
2006年6月初第一次觀察到了荒漠竹節蟲的卵:長橢圓形,具灰、褐色圖案,類似植物的種子,卵長4.9-5.5 mm,寬1.15-1.20mm,高1.0-1.2mm;卵蓋一端為前極,略向背面彎曲,U 形卵孔杯位于卵背面后1/3 處,外殼堅硬而脆,表面具網狀的紋飾,背面中線自卵孔杯之后一直延伸到卵囊末端。卵通常是單個排出,卵蓋朝上,被分散地黏附在寄主植物的枝條上,具有很好的隱蔽性。自然條件下,曾最多在一株寄主植物上發現3 枚卵。室內飼養統計表明,每只雌蟲產卵數為30-50 枚(圖3,4,5)。

圖2 野外的荒漠竹節蟲雌蟲Fig.2 A female Sceptrophasma bituberculatum in the field
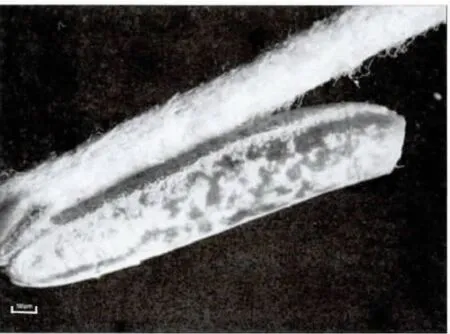
圖3 荒漠竹節蟲卵側面觀Fig.3 Lateral view of egg of Sceptrophasma bituberculatum
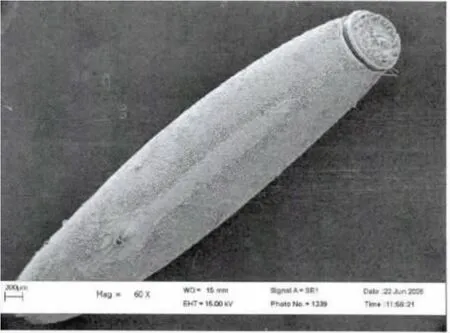
圖4 荒漠竹節蟲卵的亞顯微結構Fig.4 Dorsal view of egg of Sceptrophasma bituberculatum by SEM
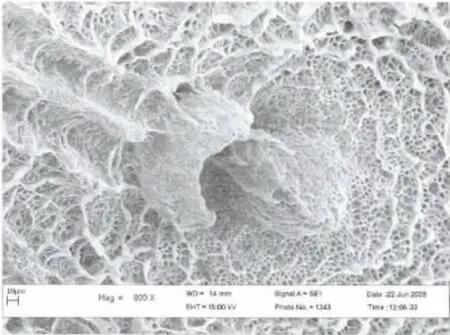
圖5 卵孔杯表面亞顯微結構Fig.5 Micropyle and sponge like surface of the egg by SEM
2.4 生活史
多年野外觀察和室內飼養的結果表明:新疆分布的荒漠竹節蟲一年一代,越冬個體常常出現在每年四月中旬(氣溫8-10℃持續數天后)至9月下旬(氣溫常保持在10-18℃),成蟲常于5月中旬出現,在野外可持續存活至6月下旬。荒漠竹節蟲具有逼真的擬態,成蟲不僅在野外很難找尋,室內籠養時也常常會被它們迷惑,四齡若蟲之前階段的幼體更加難以被發現,在野外僅在極其偶然的情況下采集到8 頭四齡以前的若蟲,因此對若蟲的研究主要來源于室內的飼養和觀察。產卵最早見于5月18日,野外采集的成蟲在人工飼養和食源充分的情況下,成蟲的壽命和產卵期明顯增長,可持續到7月中旬,卵約經47 d 開始孵化,孵化最早見于7月3日,孵化高峰期在7月下旬至8月中旬,室內卵的孵化可持續到11月中旬。一齡若蟲細小,難以被發現,野外觀察的數據是通過直接觀察和對產在已標記寄主植物上卵的定期觀察推論出來的:如卵殼完整,說明尚未孵化,如卵蓋張開或脫落,內部變空,說明若蟲已孵化。新生的若蟲體長11-14 mm,身體修長柔軟,淡黃綠色,擬態和生活行為方面與成蟲相似。人工飼養條件下孵化的若蟲齡期一般為17-22 d。野外生存的荒漠竹節蟲在經過了三次蛻皮以后便開始準備越冬(9月后捕捉到的若蟲均為4齡),而室內飼養的個體由于室溫較高,雖不冬眠,在4 齡后亦不再蛻皮,直至年底死亡。

表1 荒漠竹節蟲年生活史觀察(自然條件和人工飼養條件下)Table 1 Observation of generational life history of Sceptrophasma bituberculatum both in the field and indoors in Urumqi
2.5 孵化率
觀察期間,荒漠竹節蟲在人工飼養條件下共產卵550 粒,291 粒孵化,孵化率為52.91%。考慮未孵化的卵存在滯育期,但經過觀察,未孵化的卵直到來年的7、8月依舊沒有絲毫孵化的跡象。對一些當年和前一年沒有孵化的卵在顯微鏡下解剖觀察,發現雖然有些卵即將完成胚胎發育,并且可以明顯看出幼體結構,卻沒有絲毫生命跡象,并且散發著腐爛的味道(圖6)。另外一些則根本沒有發育,卵內物質呈干涸凝固狀,約占原體積的1/2。
2.6 蛻皮

圖6 幾乎完全發育的死亡胚胎Fig.6 An almost fully developed dead embryo from unhatched egg
竹節蟲目的昆蟲為半變態發育,荒漠竹節蟲需經過五次蛻皮羽化為成蟲。人工紗籠中飼養的荒漠竹節蟲每個齡期持續17-22 d,1-5 齡蟲體體長分別為:10-14mm(圖7),19-21mm,25-28 mm,32-39 mm 和46-54 mm。每次蛻皮前蟲體明顯膨脹,蛻皮時若蟲緊緊攀附在寄主植物的枝干上,頭部朝下彎向胸部,蛻皮從頭部開始,然后是胸部,腹部,自然狀態下蛻皮常常發生在早晨,歷時約30 min,條件不適時可能發生在白天,蛻皮時間也會相應延長,如果受到驚擾可能還會終止并導致死亡。人工飼養初期,在塑料罐中飼養的若蟲因沒有提供枝條可攀附,曾觀察到它們腹面向上靜止躺在罐底,但第二天卻發現它們再次“復活”,且身體明顯變長并完成了一次蛻皮。若蟲一般不取食蟲蛻。

圖7 荒漠竹節蟲剛孵化的若蟲Fig.7 A newly hatched nymph of Sceptrophasma bituberculatum
2.7 再生
荒漠竹節蟲有附肢再生的現象。飼養觀察期間,一頭剛孵化的若蟲在第二天便失去了右側前足,經過17 d,發生第一次蛻皮以后,斷損的足再生,包括腿節、脛節和跗節(僅有4個跗分節),腿節短而直,脛節和跗節向背方盤曲一圈余,爪呈芽狀,缺角質尖端(圖8);但該若蟲5 d后死亡,沒能繼續觀察到再生附肢的生長狀況。而觀察中,在五齡若蟲階段,附肢有斷損,隨著最后一次蛻皮,斷損處并沒有新的肢芽長出。

圖8 二齡若蟲再生的前足Fig.8 Regenerated fore leg of a second instar nymph
2.8 防御
荒漠竹節蟲的自我防御行為較為溫和,荒漠竹節蟲在日間一直保持前足外伸的姿勢,呈其寄主植物干枯的枝干狀,與寄主植株融為一體,很難被發現,一旦有風吹草動,它們還會在枝干上來回搖曳模仿樹枝的擺動;黃昏時分荒漠竹節蟲比較活躍,有較強的取食行為,食量較大;受到外界驚擾時,便會立即逃向植叢深處,同時腹部向背方卷曲(模仿蝎的行為,其生境中常有蝎的分布)作為最常見的防御行為。
此外,舍棄附肢的自殘行為應該也是它們逃脫敵害的一種防御方式,在戶外采集時常可見到斷足的蟲體。經過觀察,低齡若蟲缺失的附肢有再生現象。沒有假死和放射化學物質的防御行為。
2.9 越冬
與其他的竹節蟲目的昆蟲不同,荒漠竹節蟲產卵后沒有滯育現象,很快開始個體發育,多以四齡若蟲度過寒冷的冬天。在野外采集中,2006年4月16日所采集的標本為四齡若蟲,體長32 mm,當時室外溫度達到8-10℃,2007年9月27日和2009年10月3日采集的標本依舊是四齡若蟲(9月27日采集標本長度39 mm),當時室外溫度為10-18℃,沒有觀察到具體的越冬行為和場所。
2.10 雌雄比例及繁殖方式
在對荒漠竹節蟲的觀察研究中,共采集標本54 頭,均為雌體,未見若蟲經室內單獨飼養羽化,產出的卵可孵化為若蟲,可確定孤雌生殖為其繁殖方式之一。
2.11 現狀
過去20年的野外調查表明,目前在新疆北部的荒漠竹節蟲呈點狀分散分布。盡管在某些地區很難找到1 頭蟲體,但在合適的生境分布區中依舊存在著一定的種群,只要發現1 頭,便會在它周圍發現其他蟲體,例如,2006年5月1日在一處蒿屬植物叢生的山谷中,發現了12 頭5 齡若蟲。但是由于該物種無翅且活動性差,環境的變化和人類活動的影響極易使其種群數量受到威脅,隨后2年在上述地點采集的個體就越來越少;同樣的情形也發生在準噶爾盆地南緣的荒漠上。
隨著近幾年城市建設的劇烈膨脹及人工綠化力度的加大,荒漠竹節蟲的自然生境受到了嚴重的破壞和威脅,分布地也越來越少,僅2006年至2013年間就有3個分布點消失。
3 結論與討論
鑒于竹節蟲目的昆蟲多分布在熱帶、亞熱帶地區,同時有Whiting et al.(2003)的系統研究結果,我們推測分布在中亞干旱荒漠的荒漠竹節蟲是一個古老的物種,它們所分布的區域曾經是類似于熱帶炎熱潮濕的環境,隨著氣候的改變,荒漠竹節蟲也隨環境的變化發生了一定的適應性改變。
除了和寄主植物枝干逼真的擬態以外,荒漠竹節蟲還具有許多荒漠昆蟲相同的適應性特征,如體表較為堅硬、體色灰色或土黃褐色。同樣,荒漠竹節蟲所產的卵形態似植物的種子,卵殼的結構可以很好地在干旱的環境中保留水分,同時又透氣保證了胚胎發育所需氧氣(吳衛和范兆田,2010)。
據Bedford(1976,1978)研究,生活在熱帶的竹節蟲目昆蟲種類常不發生滯育,但卵的發育時間差異極大——即使是同一物種的同一只雌體在24 h 內產的卵也是如此,干燥的環境還會使Carausius morosus 卵的發育延長;生活在溫帶地區的種類因成體受光周期影響較大,卵的發育通常在冬季會發生滯育或發育延遲,如Bacillus rossius,長光照周期時產的卵不經歷滯育,短光照時所產的卵需經歷滯育。本文中荒漠竹節蟲分布在溫帶內陸干旱區,產卵于長日照期(5月下旬至7月上旬),卵的發育沒有滯育期,產卵后一個半月左右便開始孵化,與文獻記載的其它需經歷數月至2 a滯育的種類(陳樹椿,2008)有很大差異,這種孵化方式應該也是它在進化中對干旱荒漠生境的一種適應,如同荒漠中的短命植物,如此可以最大程度地利用卵內的水分,防止因水分散失而導致的發育失敗;對未孵化卵的解剖表明,其中一部分卵的卵質干涸,這可能是其孵化失敗的主要原因,而另一部分卵發育失敗則是因為受到了微生物的感染。人工飼養條件下溫濕度比較恒定,實驗記錄卵的孵化時間甚至可持續至12月初,說明溫度和濕度對胚胎發育有影響。荒漠竹節蟲的孵化是在一年中最炎熱、干旱的季節,寄主植物幾近干枯,食源狀況不利于其若蟲的生長,但在生存策略上講,比起會因滯育而導致卵內水分散失最終不能孵化,應屬權宜之計了。
本文觀察到荒漠竹節蟲以四齡若蟲越冬。新疆冬季寒冷,尤其是荒漠,常見昆蟲通常會鉆入沙土中或石塊下方越冬,但竹節蟲體型細長,難以做到,結合其生境,推測它們多隱蔽在寄主植物的枝叢中或者沙石土縫中來度過漫長的冬季,荒漠竹節蟲越冬的機制仍需進一步觀察研究。
孤雌生殖是昆蟲中較為常見的生殖方式,也是昆蟲適應惡劣環境的一種表現(王成業,2011)。文獻記載荒漠竹節蟲有雄體(Бей-Биенко,1964;陳樹椿和何允恒,2008;Hennemann et al.,2008)。而在我們歷年來野外采集的54 頭標本中(四齡若蟲9 頭,五齡成蟲45頭)未見雄蟲,將五齡蟲體單獨室內飼養至產卵表明孤雌生殖是其繁殖方式之一;具體在何時出現雄體、并且發生兩性生殖尚需進一步研究。
因荒漠竹節蟲擬態性很強、野外難以發現,本研究結果是建立在室內外共同觀察的結果之上。觀察表明:溫度、濕度和食物對荒漠竹節蟲的生存影響很大。其中,室外溫差表現為日夜溫差(一般大于10℃)、季節溫差(極差可達55-80℃);夏季日間荒漠濕度常常低于10%,而室內則始終為30%左右;荒漠竹節蟲對食物的要求也較高,例如,對從阜康荒漠和雅馬里克山采集回來的竹節蟲同時投喂兩個采集地的蒿,它們都明顯只取食自己采集地的。因此,室內飼養的低齡幼蟲未能進入下一齡期就死亡的數量較多;從野外采集帶回室內飼養的四齡若蟲因室內溫度等原因不能進入越冬狀態,但也不會繼續發生蛻皮最終死亡;但同時室內飼養條件下,成蟲壽命較野外延長很多,產卵時間也相應延長;卵的孵化可隨之持續。
荒漠竹節蟲在新疆是體長最長的昆蟲,由于數量很少,在林、牧業上雖沒有十分重要的經濟意義,但仍然是一種珍稀的值得收藏的昆蟲,尤其在昆蟲對荒漠環境的適應和進化研究上是一種很好的研究材料。在此我們僅對荒漠竹節蟲的一般形態特征和生物學特征進行了研究,今后將對它在荒漠環境中的適應性及遺傳多樣性方面進一步研究,為對該物種的多樣性保護提供參考。
References)
Bedford GO.Biology and ecology of the Phasmatodea[J].Annual Review of Entomology,1978,23(1):125-149.
Bedford GO.Description and development of the eggs of two stick insects(Phasmatodea:Phasmatidae)from New Britain[J].Australian Journal of Entomology Society,1976,15(4):389-393.
Cai BL.A List of Chinese phasmids with the key to subfamilies[C].In:Contributions From Tianjin Natrual Museum,1987,4:26-30.[蔡保靈.中國竹節蟲名目及亞科檢索表[C].見:天津自然博物館論文集,1987,4:26-30]
Chen PC,Chen SC.Identification,biology and control of the important species of phasmatodea in China[J].Journal of Beijing Forestry University,1997,19(4):70-75.[陳培昶,陳樹椿.中國重要竹節蟲的鑒別、生物學及其防治[J].北京林業大學學報,1997,19(4):70-75]
Chen SC,He YH.Phasmatodea of China[M].Beijing:China Forestry Publishing House,2008,1-476.[陳樹椿,何允恒.中國蟲脩目昆蟲[M].北京:中國林業出版社,2008,1-476]
Hennemann FH,Conle OV,Zhang WW.Catalogue of the stick and leaf-insects(Phasmatodea)of China,with a faunistic analysis,review of recent ecological and biological studies and bibliography(Insecta:Orthoptera:Phasmatodea)[J].Zootaxa,2008,1735:1-77.
Huang RX,Wu W,Mao XF,et al.The Fauna of the Desert Insects of Xinjiang and its Formation and Evolution[M].Urumqi:Xinjiang Science and Technology Publishing House,2005,1-232.[黃人鑫,吳衛,毛新芳,等.新疆荒漠昆蟲區系及其形成與演變[M].烏魯木齊:新疆科技出版社,2005,1-232]
Whiting MF,Bradler S,Maxwell T.Loss and recovery of wings in stick insects[J].Nature,2003,421(6920):264-267.
Schwander T,Crespi BJ.Multiple direct transitions from sexual reproduction to apomictic parthenogenesis in Timema stick insects[J].Evolution,2009,63(1):84-103.
Wang CY.Genetic mechanism and evolutionary significance of the origin of parthenogenetic insects[J].Zoological Research,2011,32(6):689-695.[王成業.昆蟲孤雌生殖起源的遺傳機制和進化意義[J].動物學研究,2011,32(6):689-695]
Бей-Биенко ГЯ,1964.Определитель Насекомых Европейской Части Ссср[M].В Пяти Томах Ι.Издательство《Наука》.Москва-Ленинград:203-204.
Ye WY,Yin LK,Qian Yi et al.Study on the vegetation of Yama-like Mountain and plant diversity characteristic[J].Academic Journal of Xinjiang Agriculture University,2005,28(1):44-48.[葉衛英,尹林克,錢翌,等.烏魯木齊市周邊荒山植物群落類型及其物種多樣性[J].新疆農業大學學報,2005,28(1):44-48]
