白細胞介素22誘導HaCaT細胞表達肝素結合表皮生長因子樣生長因子的作用機制探討
2015-11-29 11:19:42劉欣欣羅素菊鄭焱許文娟李穎劉全忠
中華皮膚科雜志 2015年3期
關鍵詞:信號
劉欣欣 羅素菊 鄭焱 許文娟 李穎 劉全忠
白細胞介素22誘導HaCaT細胞表達肝素結合表皮生長因子樣生長因子的作用機制探討
劉欣欣 羅素菊 鄭焱 許文娟 李穎 劉全忠
目的 探討白細胞介素22(IL-22)誘導HaCaT細胞表達肝素結合表皮生長因子樣生長因子(HBEGF)的相關作用機制。方法 用不同濃度的IL-22(12.5、25、50、100μg/L)干預處理HaCaT細胞,對照組選擇等量磷酸鹽緩沖液(PBS)處理。24 h后,提取HaCaT細胞總蛋白進行蛋白免疫印跡,檢測絲裂原活化蛋白激酶-細胞外信號調節激酶1/2(MAPK-ERK1/2)通路中磷酸化ERK1/2(P-ERK1/2)的表達和轉錄激活因子途徑(JAK/STAT)中磷酸化 JAK2(P-JAK2)和磷酸化 STAT3(P-STAT3)的表達。將HaCaT細胞分 4組,分別用 PBS、IL-22、MAPK-ERK1/2抑制劑PD98059、JAK2/STAT3通路抑制劑AG490與IL-22共同干預HaCaT細胞,24 h后,提取細胞總蛋白和總mRNA,分別用蛋白免疫印跡法和實時定量逆轉錄(RT)-PCR法檢測不同處理組HB-EGF蛋白和mRNA水平的改變。采用SPSS16.0軟件進行單因素方差分析檢驗組間差異,Bonferroni檢驗進行多重比較。結果 不同濃度的IL-22干預處理后,HaCaT細胞中P-ERK1/2、P-JAK2和P-STAT3蛋白表達均高于對照組(P<0.05)。HB-EGF蛋白水平(HB-EGF/內參照的灰度比值)在PD98059組和AG490組分別為0.183±0.020和0.199±0.011,與 IL-22組(0.924±0.032)相比,差異具有統計學意義(F值分別為37.700、36.400,均P<0.05)。HB-EGF mRNA水平在PD98059組和AG490組分別為1.034±0.072和0.989±0.038,與IL-22組(1.844±0.135)相比,差異具有統計學意義(F值分別為11.271、13.429,均P<0.05)。結論 IL-22可以引起HaCaT細胞中MAPK-ERK1/2和JAK2/STAT3這兩條信號通路激活。IL-22誘導HaCaT細胞產生HB-EGF蛋白的作用機制可能與MAPK-ERK1/2和JAK2/STAT3這兩條信號通路有關。
白細胞介素-22;成纖維細胞生長因子1;角蛋白細胞;絲裂原激活蛋白激酶類;Janus激酶2;STAT3轉錄因子;銀屑病;HaCaT細胞
目前研究認為,銀屑病的發病機制主要是由T細胞介導的以表皮角質形成細胞為靶點的免疫應答反應。在其發病過程中Th17、Th22細胞及其細胞因子發揮重要作用。白細胞介素22(IL-22)為Th22細胞分泌的主要細胞因子,參與調節機體免疫反應。國內外研究表明,銀屑病患者外周血及皮損中IL-22水平較健康人高,且IL-22水平的高低與銀屑病嚴重程度密切相關[1-3]。肝素結合表皮生長因子樣生長因子(HB-EGF)是表皮生長因子家族中的一員,可促進細胞的增殖,在銀屑病皮損中表達增高,并且其表達量與銀屑病病情相關[4],提示其在銀屑病表皮異常增殖中發揮重要作用。以往研究表明,細胞外調節蛋白激酶(ERK)和信號轉導與轉錄激活因子3(STAT3)這兩種信號通路蛋白在角質形成細胞增殖信號傳導中至關重要[5]。我們的研究表明,IL-22可在基因和蛋白水平促進HaCaT細胞HBEGF的表達(未發表)。在本研究中,我們檢測IL-22對絲裂原活化蛋白激酶(MAPK-ERK1/2)通路中磷酸化ERK1/2(P-ERK1/2)和轉錄激活因子途徑(JAK/STAT)中磷酸化JAK2(P-JAK2)和磷酸化STAT3(P-STAT3)的影響,并通過抑制其信號傳導檢測其是否影響HaCaT細胞HB-EGF的表達,探討其可能的作用機制。
材料和方法
一、實驗材料
1.實驗細胞:HaCaT細胞系德國柏林自由大學本杰明·富蘭克林醫學中心惠贈,為我實驗室長期凍存細胞。
2.主要試劑:重組人IL-22(美國Protein Specialists 公 司),P-ERK1/2、P-JAK2、JAK2、PSTAT3、STAT3、辣根過氧化物酶標記(HRP)-羊抗兔的二抗(美國CST公司),β肌動蛋白、HRP-羊抗鼠的二抗、HRP-兔抗山羊二抗(北京中杉金橋公司),ECL發光顯色液、熒光Mix(美國Invitrogen公司),PD98059、AG490(美國 Millipore公司),DMEM培養基(美國Gibico公司),Thermo逆轉錄試劑盒(美國Thermo公司),HB-EGF抗體、HB-EGF蛋白ELISA試劑盒(美國RD公司)。HB-EGF及β肌動蛋白的引物由生工生物工程(上海)股份有限公司合成。
3.主要儀器:酶聯免疫檢測儀(美國Biotek公司),凝膠成像系統儀(英國Cambridge公司),實時定量PCR儀(美國Bio-Rad公司),CO2孵育箱(美國Forma Scientific公司)。
二、實驗方法
1.HaCaT細胞的復蘇與培養:將凍存的HaCaT細胞復蘇后,用含10%胎牛血清(FBS)的DMEM培養液,于5%CO2、37℃條件下培養。待HaCaT細胞90%融合后,用0.25%胰酶/乙二胺四乙酸(EDTA)消化進行傳代培養。
2.HaCaT細胞的干預處理:將經傳代培養后處于對數生長期的細胞消化后,以1×105/ml接種于6孔板,37℃、5%CO2孵育培養。待細胞70%~80%融合時,進行饑餓處理2~3 h后,更換新的培養液,分別用 12.5、25、50、100μg/L 重組人 IL-22 進行干預,對照組用等量磷酸鹽緩沖液(PBS)干預。繼續于37℃、5%CO2孵育培養 24 h。
3.蛋白免疫印跡檢測信號通路相關蛋白:干預處理后的HaCaT細胞,用預冷PBS洗3遍,每孔加300μl細胞裂解液和2μl蛋白酶抑制劑苯甲基磺酰氟(PMSF)。冰上裂解30min,用EP管收集細胞裂解液離心10min。測蛋白濃度后進行Western印跡。取上清,10%聚丙烯酰胺凝膠蛋白電泳,每孔上樣量 25μl,電泳 90min,穩流 300 mA 轉膜 40min,轉膜后的硝酸纖維素膜分別用兔抗人一抗(PERK1/2、P-JAK2、JAK2、P-STAT3、STAT3)4 ℃孵育過夜,后洗滌,用二抗(HRP標記的羊抗兔二抗)室溫孵育2 h,ECL暗室曝光顯色。檢測P-ERK1/2、PSTAT3、P-JAK2的表達情況。每組實驗重復3次。
4.細胞信號通路阻斷劑阻斷實驗:
(1)細胞干預處理:對接種培養的HaCaT細胞分為4 組:①陰性對照組:PBS;②IL-22 組:50μg/L IL-22;③PD98059 組:50μmol/L PD98059+50μg/L IL-22;④AG490 組:50μmol/L AG490+50μg/L IL-22。抑制劑的選擇及使用濃度參照文獻[6-7]。
(2)蛋白免疫印跡檢測HB-EGF蛋白水平的改變:干預培養24 h后,按上述方法提取各組細胞的總蛋白進行蛋白免疫印跡,檢測HB-EGF蛋白水平的改變。實驗重復3次。
(3)實時熒光定量RT-PCR檢測HB-EGF基因水平的改變:干預處理24 h后吸去培養液,用PBS洗2遍,用Trizol試劑提取細胞總RNA,用紫外分光光度計測吸光度(A260nm/A280nm),檢測RNA的純度及濃度。應用逆轉錄試劑盒合成cDNA。熒光定量PCR 反應體系:25μl,冰上加樣熒光定量 Mix12.5μl,上下游引物各 0.25μl,模板 cDNA 2μl,無 RNA 酶的雙蒸水10μl。按照試劑盒說明設定反應條件50℃2min促使尿嘧啶-DNA糖基化酶(UDG)激活:然后95℃,2min預變性,1個循環;95℃15 s變性,60℃1min退火延伸,40個循環。進行熔解曲線分析判定PCR反應的特異性。HB-EGF上游引物:5′-GGCTCCCTCCTGCATCTG-3′,下游引物:5′-AGGAT GGTTGTGTGGTCATAGGT-3′。β肌動蛋白上游引物:5′-CCTGGCACCCAGCACAAT-3′,下游引物:5′-CTGATCCACATCTGCTGGAA-3′。合成的cDNA進行實時定量PCR擴增。每個樣本設3個復孔。上述RT-PCR實驗重復3次。
(4)ELISA法檢測HB-EGF蛋白的表達:采用雙抗體夾心ELISA法,參照試劑盒使用說明書進行。用酶標儀在450 nm波長處測量各孔的吸光度。每個樣本重復3次。制作樣品濃度標準曲線,并據其計算各樣品的濃度。應用二喹啉甲酸(BCA)法測樣品總蛋白濃度。統計不同樣品HB-EGF蛋白濃度與總蛋白濃度的比值。
三、統計結果處理
結 果
一、IL-22干預后HaCaT細胞中P-ERK1/2、PSTAT3、P-JAK2蛋白的表達
免疫印跡結果顯示,12.5、25、50、100μg/L IL-22干預處理后,HaCaT細胞MAPK通路中P-ERK1/2表達量、JAK2/STAT3通路中P-STAT3表達量、PJAK2表達量均較對照組升高(表1),且與IL-22濃度有一定相關關系。3次重復實驗結果類似(圖1、2)。參照課題組前期研究及信號通路蛋白表達選擇50μg/L的IL-22進行信號通路阻斷實驗。
表1 IL-22干預后HaCaT細胞中P-ERK1/2、P-STAT3、P-JAK2蛋白的表達量(與β肌動蛋白灰度比值,±s)
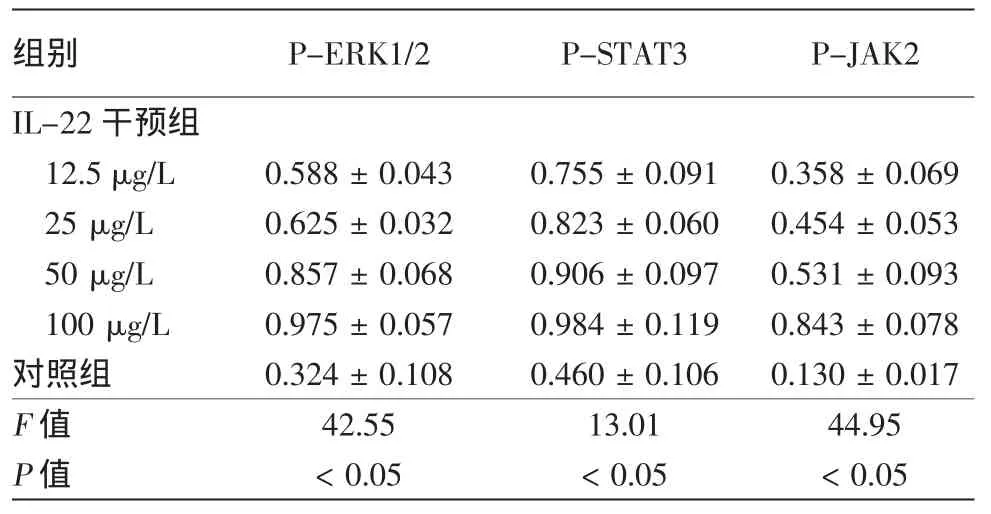
表1 IL-22干預后HaCaT細胞中P-ERK1/2、P-STAT3、P-JAK2蛋白的表達量(與β肌動蛋白灰度比值,±s)
注:n=3
組別 P-ERK1/2 P-STAT3 P-JAK2 IL-22干預組12.5μg/L 0.588±0.043 0.755±0.091 0.358±0.069 25μg/L 0.625±0.032 0.823±0.060 0.454±0.053 50μg/L 0.857±0.068 0.906±0.097 0.531±0.093 100μg/L 0.975±0.057 0.984±0.119 0.843±0.078對照組 0.324±0.108 0.460±0.106 0.130±0.017 F值 42.55 13.01 44.95 P值 <0.05 <0.05 <0.05
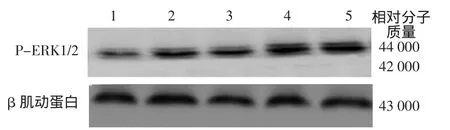
圖1 白細胞介素22(IL-22)對HaCaT細胞MAPK通路蛋白ERK及P-ERK表達的影響 1:對照組;2~5:分別為12.5、25、50、100μg/L IL-22 組

圖2 白細胞介素22(IL-22)對HaCaT細胞JAK2/STAT3通路蛋白表達的影響 1:對照組;2~5:分別為12.5、25、50、100μg/L IL-22組

圖3 部分信號通路阻斷HaCaT細胞中肝素結合表皮生長因子樣生長因子的表達 1:對照組;2:50μg/L IL-22;3:50μg/L IL-22+PD98059;4:50μg/L IL-22+AG490
二、通路阻斷實驗結果
1.加入信號通路阻斷劑后HB-EGF蛋白水平的改變:蛋白免疫印跡結果顯示,在加入信號通路阻斷劑后,HB-EGF蛋白表達量(HB-EGF/β肌動蛋白灰度比值)PD98059組為0.183±0.020,較IL-22組(0.924±0.032) 降低(F=37.700,P<0.05);AG490組(0.199±0.011)也較IL-22組降低(F=36.400,P<0.05)。結果見圖3。
2.加入信號通路阻斷劑后HB-EGF mRNA水平的變化:加入信號通路阻斷劑后HB-EGF mRNA表達水平均較IL-22組減少。PD98059組HB-EGF mRNA表達水平(2-ΔΔCt)為1.034±0.072,較IL-22組(1.844±0.135)降低(F=11.271,P<0.05);同時AG490組(0.989±0.038)較IL-22組亦降低(F=13.429,P<0.05)。
三、不同干預處理組HB-EGF蛋白的濃度與總蛋白濃度的比值
在加入信號通路抑制劑后,HB-EGF蛋白表達均較IL-22組降低。HB-EGF與總蛋白濃度比值在對照組為(0.816±0.152)× 10-3,PD98059 組為(0.852±0.159)×10-3,AG490組為(1.049±0.166)×10-3,后兩組與 IL-22組(2.067±0.160)× 10-3比較均降低,F值分別為20.933、5.414,均P<0.05。
討 論
本實驗研究結果顯示,IL-22可誘導HaCaT細胞MAPK信號通路中P-ERK1/2蛋白表達增加,加入MAPK-ERK1/2通路特異性抑制劑PD98059后,HB-EGF蛋白和mRNA的表達較未加抑制劑組明顯降低。提示MAPK-ERK1/2信號通路在IL-22促進HB-EGF的表達中發揮重要作用。同樣濃度IL-22干預后,JAK2/STAT3信號通路中P-JAK2和P-STAT3蛋白水平均明顯升高。加入JAK2/STAT3通路抑制劑AG490后,HB-EGF無論從蛋白水平還是基因水平均較無抑制組降低。
有研究顯示,IL-22可通過促進HaCaT細胞PERK1/2和P-STAT3升高進而引起角蛋白17升高[8]。HB-EGF可促進角質形成細胞的增殖和遷移。研究表明,HB-EGF在正常皮膚表皮中不表達或表達量很低,而在銀屑病患者皮損中表達升高,其表達水平與銀屑病的嚴重程度有一定的相關性,提示其在銀屑病表皮異常增殖中發揮重要作用[9]。IL-22受體廣泛分布于免疫細胞中,參與調控角質形成細胞、內皮細胞、成纖維細胞等其他組織細胞的增殖和分化,尤其在角質形成細胞的增殖、分化及免疫中起重要作用[10]。在銀屑病皮損中IL-22受體較正常皮膚增多,提示IL-22受體參與銀屑病的表皮異常增殖[11]。因此我們推測IL-22通過與HaCaT細胞表面IL-22受體結合引起細胞內通路激活。
銀屑病皮損的棘層和基底層角質形成細胞中存在ERK1/2的磷酸化,并且與角質形成細胞的過度增殖相關[12-13]。另外,ERK1/2的磷酸化可促進T細胞活化、增殖和抗凋亡作用[14]。研究表明,在銀屑病患者的皮損中STAT3表達明顯升高[15-16]。在銀屑病患者機體內STAT3參與Th17細胞信號傳導過程,與角質形成細胞過度增殖和炎性浸潤有關[17]。
綜上所述,IL-22可使HaCaT細胞HB-EGF、PERK1/2蛋白、P-JAK2和P-STAT3蛋白水平升高。MAPK-ERK1/2通路特異性抑制劑PD98059和JAK2/STAT3通路抑制劑AG490可明顯抑制HBEGF的表達,提示IL-22可能通過MAPK-ERK1/2和JAK2/STAT3這兩條通路誘導HB-EGF的表達。提示阻斷IL-22信號通路傳導可減少HB-EGF的表達,進一步研究可能為治療銀屑病提供一個新的治療途徑。
[1]Kagami S,Rizzo HL,Lee JJ,et al.Circulating Th17,Th22,and Th1 cells are increased in psoriasis[J].J Invest Dermatol,2010,130(5):1373-1383.
[2]Sabat R,Wolk K.Research in practice:IL-22 and IL-20:significance for epithelial homeostasis and psoriasis pathogenesis[J].J Dtsch Dermatol Ges,2011,9(7):518-523.
[3]史玉玲,徐曉光,畢新嶺,等.白介素22及其相關因子在銀屑病患者血清及外周血單一核細胞中的表達[J].中華皮膚科雜志,2011,44(4):238-240.
[4]Yoshida A,Kanno H,Watabe D,et al.The role of heparin-binding EGF-like growth factor and amphiregulin in the epidermal proliferation of psoriasis in cooperation with TNFα [J].Arch Dermatol Res,2008,300(1):37-45.
[5]Wang Z,Jin C,Li H,et al.GPR48-Induced keratinocyte proliferation occurs through HB-EGF mediated EGFR transactivation[J].FEBS Lett,2010,584(18):4057-4062.
[6]Lee H J,Lee H J,Sohn EJ,et al.Inhibition of connexin 26/43 and extracellular-regulated kinase protein plays a critical role in melatonin facilitated gap junctional intercellular communication in hydrogen peroxide-treated HaCaT keratinocyte cells[J/OL].Evid Based Complement Alternat Med,2012,2012:589365[2013-09-06].http://www.hindawi.com/journals/ecam/2012/589365/.
[7]Boissel JP,Ohly D,Bros M,et al.The neuronal nitric oxide synthase is upregulated in mouse skin repair and in response to epidermal growth factor in human HaCaT keratinocytes[J].J Invest Dermatol,2004,123(1):132-139.
[8]Zhang W,Dang E,Shi X,et al.The pro-inflammatory cytokine IL-22 up-regulates keratin 17 expression in keratinocytes via STAT3 and ERK1/2[J/OL].PLoS One,2012,7(7):e40797[2013-06-09].http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0040797.
[9]Zheng Y,Peng Z,Wang Y,et al.Alteration and significance of heparin-bindingepidermal-growth-factor-like growth factorin psoriatic epidermis[J].Dermatology,2003,207(1):22-27.
[10]Sonnenberg GF,Fouser LA,Artis D.Functional biology of the IL-22-IL-22R pathway in regulating immunity and inflammation at barrier surfaces[J].Adv Immunol,2010,107:1-29.
[11]Tohyama M,Hanakawa Y,Shirakata Y,et al.IL-17 and IL-22 mediate IL-20 subfamily cytokine production in cultured keratinocytes via increased IL-22 receptor expression [J].Eur J Immunol,2009,39(10):2779-2788.
[12]Takahashi H,Ibe M,Nakamura S,et al.Extracellular regulated kinase and c-Jun N-terminal kinase are activated in psoriatic involved epidermis[J].J Dermatol Sci,2002,30(2):94-99.
[13]Haase I,Hobbs RM,Romero MR,et al.A role for mitogen-activated protein kinase activation by integrins in the pathogenesis of psoriasis[J].J Clin Invest,2001,108(4):527-536.
[14]Laatikainen LE,Incoronato M,Castellone MD,et al.SOD3 decreases ischemic injury derived apoptosis through phosphorylation of Erk1/2,Akt,and FoxO3a[J/OL].PLoS One,2011,6(8):e24456[2013-06-14].http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0024456.
[15]Sano S,Chan KS,Carbajal S,et al.Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model[J].Nat Med,2005,11(1):43-49.
[16]Andrés RM,Hald A,Johansen C,et al.Studies of Jak/STAT3 expression and signalling in psoriasis identifies STAT3-Ser727 phosphorylation as a modulator of transcriptional activity [J].Exp Dermatol,2013,22(5):323-328.
[17]LiuH,Moroi Y,Yasumoto S,et al.Immunohistochemical localization of activated Stat3 and hTERT protein in psoriasis vulgaris[J].Eur J Dermatol,2006,16(2):205-207.
2014-04-24)
(本文編輯:尚淑賢)
Mechanisms underlying interleukin-22-induced expression of heparin-binding epidermal growth factor-like growth factor in HaCaT cells
Liu Xinxin*,Luo Suju,Zheng Yan,Xu Wenjuan,Li Ying,Liu Quanzhong.*Department of Dermatology,Tianjin Medical University General Hospital,Tianjin 300052,China
Luo Suju,Email:luosuju2005@163.com
ObjectiveTo investigate the mechanisms underlying interleukin-22(IL-22)-induced expression of heparin-binding epidermal growth factor-like growth factor(HB-EGF)in HaCaT cells.MethodsSome HaCaT cells were divided into several inverention groups treated with IL-22 at concentrations of 12.5,25,50,100μg/L,respectively and a control group treated with phosphate buffer saline (PBS).After 24-hour culture,total proteins were extracted from the HaCaT cells,and Western blot was performed to measure the expression of phosphorylated extracellular signalregulated kinase 1/2 (P-ERK1/2)in the mitogen-activated protein kinase (MAPK)-ERK1/2 pathway,as well as phosphorylated-JAK2(P-JAK2)and phosphorylated-signal transducer and activator of transcription 3 (P-STAT3)in the JAK2/STAT3 pathway.In a blocking experiment,some HaCaT cells were divided into 4 groups to be treated with PBS,IL-22,PD98059 (an inhibitor of MAPK-ERK1/2)combined with IL-22 (PD98059 group),AG490 (an inhibitor of JAK2/STAT3)combined with IL-22 (AG490 group),respectively.After 24-hour treatment,total proteins and mRNAs were extracted from the HaCaT cells followed by Western blot and real-time quantitative reverse transcription-PCR for the measurement of protein and mRNA expressions of HB-EGF respectively.Statistical analysis was carried out with the software SPSS 16.0 by one-way analysis of variance (ANOVA)for intergroup comparisons and by Bonferroni′s test for multiple comparisons.ResultsAfter treatment with IL-22 at the above 4 concentrations,the expressions of P-ERK1/2,P-JAK2 and P-STAT3 in HaCaT cells were all increased compared with the control group(allP<0.05).The protein and mRNA expression levels(expressed as the HB-EGF/β-actin ratio and 2-△△Ctrespectively)of HB-EGF were both significantly decreased in the PD98059 group and AG490 group than in the IL-22 group(protein:0.183±0.020 and 0.199±0.011 vs.0.924±0.032,F=37.700,36.400,respectively,bothP<0.05;mRNA:1.034±0.072 and 0.989±0.038 vs.1.844±0.135,F=11.271,13.429,respectively,bothP<0.05).ConclusionsIL-22 can activate the MAPK-ERK1/2 and JAK2/STAT3 signaling pathways in HaCaT cells,which may contribute to IL-22-induced expression of HB-EGF in HaCaT cells.
Interleukin-22;Fibroblast growth factor 1;Keratinocytes;Mitogen-activated protein kinases;Janus kinase 2;STAT3 transcription factor;Psoriasis;HaCaT cells
10.3760/cma.j.issn.0412-4030.2015.03.009
國家自然科學基金(81201231、81371732)
300052天津醫科大學總醫院皮膚科(劉欣欣、羅素菊、許文娟、劉全忠),肺癌研究所(李穎);西安交通大學第二醫院皮膚科(鄭焱)
羅素菊,Email:luosuju2005@163.com
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
