毒殺南方根結線蟲的木霉種類鑒定及活性研究
2015-11-25 08:31:49韓冰潔王媛媛朱曉峰段玉璽陳立杰
植物保護 2015年2期
關鍵詞:綠色
焦 俊,韓冰潔,王媛媛,朱曉峰,段玉璽,陳立杰
(沈陽農業大學植物保護學院,北方線蟲研究所,沈陽 110866)
?
毒殺南方根結線蟲的木霉種類鑒定及活性研究
焦 俊,韓冰潔,王媛媛,朱曉峰,段玉璽,陳立杰*
(沈陽農業大學植物保護學院,北方線蟲研究所,沈陽 110866)
木霉是國際上研究與應用非常普遍的生防真菌之一,具有重要的科研價值和廣闊的應用前景。本研究采用形態學鑒定、ITS和β-tubulin序列分析法對4株具有殺線蟲活性的木霉菌株進行種類鑒定,結果顯示FT311和FT312為綠色木霉(Trichodermaviride),FT1937和FT85為哈茨木霉(T.harzianum)。利用貝氏皿浸沒法測定,4株木霉發酵濾液均能顯著降低南方根結線蟲卵的孵化和2齡幼蟲(J2)的活性,其中FT1937和FT85對卵孵化抑制率和J2致死率均達到85%~92%,而FT311能夠達到80%~85%,但FT312毒性較低,僅達40%~60%;2株哈茨木霉的活性好于2株綠色木霉;同時觀察到發酵液對線蟲卵和幼蟲體壁有消解作用、抑制和延遲卵孵化,并使幼蟲體內空泡化。本研究為根結線蟲病害的生防防治提供了更多的木霉資源信息。
木霉; 種類鑒定; 南方根結線蟲; 殺線蟲活性
根結線蟲(Meloidogynespp.)是茄科、葫蘆科和果樹等多種植物的重要病原,能夠危害植物的根部,造成根系發育受阻和腐爛,給農業生產造成巨大的經濟損失[1]。近年來,隨著設施蔬菜種植業的迅速發展,東北地區根結線蟲病害愈來愈嚴重,尤其在遼寧省,由南方根結線蟲(MeloidogyneincognitaChitwood)引致的各類蔬菜根結線蟲病正嚴重威脅著老棚區設施蔬菜的生產[2]。由于蔬菜的周年性生產和持續性采摘,高毒高殘留的化學殺線劑被限制使用,因此開發利用安全有效的殺線劑已經成為植物線蟲病害防治工作中亟待解決的問題。
木霉(Trichodermaspp.)隸屬于子囊菌門(Ascomycota)糞殼菌綱(Sordariomycetes)肉座菌目(Hypocreales)肉座菌科(Hypocreaceae)真菌,是世界上研究最多的有效生防真菌之一。但木霉的研究多集中于防控植物真菌病害,如用多種木霉防治水稻紋枯病[3],利用木霉制劑防治西瓜枯萎病等[4],一些研究明確了木霉的作用機制包括快速生長的競爭機制、重寄生作用以及抑菌活性等[5]。近年來也有研究發現木霉對植物線蟲有一定防控效果,如長枝木霉對根結線蟲的寄生作用[6],哈茨木霉防控爪哇根結線蟲[7]以及哈茨木霉發酵液對根結線蟲的致死作用[8]等。本文針對實驗室前期篩選獲得的幾株對植物線蟲有活性的木霉菌株,利用形態學和DNA信息鑒定到種,并檢測其對南方根結線蟲的活性,以期為蔬菜根結線蟲病防控提供新的生防真菌資源。
1 材料與方法
1.1 試驗材料
供試線蟲:南方根結線蟲(Meloidogyneincognita)2齡幼蟲J2。繁殖自北方線蟲研究所的溫室大棚,將接種南方根結線蟲的番茄幼苗培養40 d,待根結上產生卵囊后,將番茄根系取出,用水輕輕沖洗,置于1%的次氯酸鈉中消毒3 min,再用無菌水沖洗3 次,放入內有少量無菌水的培養皿,25 ℃恒溫箱中培養,每隔24 h收集一次新孵化的2齡幼蟲J2。
供試菌株:菌株FT311和FT312分離自廣西玉林市八角根圍土壤,FT85分離自吉林省撫松縣人參根際土壤,FT1937來自于實驗室前期保存的菌種。
將保存菌株從冷庫中取出,在PDA平板上進行菌株活化,25 ℃培養后在4 ℃恒溫箱中保存待用。將活化菌株接種于查氏培養基,于25 ℃150 r/min液體搖瓶發酵5 d。發酵液抽濾出菌絲,置于4 ℃恒溫箱中保存備用。經測定4種木霉菌發酵液pH為6.3~6.8之間,均呈弱酸性。
1.2 試驗方法
1.2.1 木霉的形態學鑒定
將菌株用接種針挑至PDA平板中央,25 ℃恒溫箱中培養,每隔12 h觀察菌落的顏色和形態。48 h后于顯微鏡下觀察和測量菌絲、分生孢子梗及分生孢子的形態與大小。依據Rifai和Bissett的方法進行分類鑒定[9-10]。
1.2.2 木霉的分子生物學鑒定
利用天根新型基因組DNA提取試劑盒提取4種木霉菌株的基因組DNA,-20 ℃保存備用。采用真菌rDNA-ITS通用引物ITS1和ITS4[11]和β-tubulin基因種間引物Bt2a和Bt2b[12]對菌株基因組DNA進行PCR擴增,引物序列如下:
ITS1:5′-TCCGTAGGTGAACCTGCGG-3′
ITS4:5′-TCCTCCGCTTATTGATATGC-3′
Bt2a:5′-GGTAACCAAATCGGTGCTGCTTTC-3′
Bt2b:5′-ACCCTCAGTGTAGTGACCCTTGGC-3′
①rDNA-ITS PCR擴增的反應體系:10×反應緩沖液(10×PCR buffer)2.5 μL,MgCl2(25 mmol/L)2.0 μL,dNTP (2.5 mmol/L)1.5 μL,Taq(5 U/μL)0.2 μL,ITS1(10 μmol/L)2.0 μL,ITS4(10 μmol/L)2.0 μL,模板2.0 μL,ddH2O 12.8 μL,總體積25.0 μL。rDNA-ITS基因PCR擴增反應的條件為:94 ℃預變性5 min;94 ℃變性30 s,51 ℃退火40 s,72 ℃延伸90 s,30個循環;72 ℃延伸7 min,最后于4 ℃保存。
②β-tubulin基因PCR擴增的反應體系:10×反應緩沖液(10×PCR buffer)2.5 μL,MgCl2(25 mmol/L)2.0 μL,dNTP (2.5 mmol/L)2.0 μL,Taq(5 U/μL)0.25 μL,Bt2a/CladAct-F3 (10 μmol/L)1.0 μL,Bt2b/CladAct-R3 (10 μmol/L)1.0 μL,模板2.0 μL,ddH2O 14.25 μL,總體積25.0 μL。β-tubulin基因PCR擴增反應的條件為:94 ℃預變性3 min;94 ℃變性30 s,61 ℃退火30 s,72 ℃延伸1 min,39個循環;72 ℃延伸10 min,最后于4 ℃保存。
擴增產物經1%瓊脂糖凝膠電泳檢測,引物合成及PCR產物的純化和序列測定委托上海生工生物工程技術服務有限公司完成。
ITS和β-tubulin序列結果利用MEGA軟件進行序列分析和比較。利用美國國家生物工程信息中心(NCBI,http:∥www.ncbi.nlm.nih.gov/BLAST)的BLAST網絡工具與GenBank數據庫中相關真菌菌株的序列進行最高同源性比對,并分別對ITS和β-tubulin測序結果進行基因序列數據分析,從DNA水平確定木霉屬的種類及遺傳進化關系。
1.2.3 木霉對南方根結線蟲卵囊中卵孵化的抑制作用
將消毒后的卵囊擠壓破碎,得到未孵化的線蟲卵。在每個貝氏小皿中放入100粒卵并分別加入4種木霉的發酵液各800 μL,3次重復,以無菌發酵培養液為對照。分別于24、48、72、96 h觀察線蟲卵的孵化情況,比較木霉發酵濾液對線蟲卵孵化抑制率的差異。
相對抑制率公式如下:
相對抑制率(%)=

1.2.4 木霉菌株發酵液對南方根結線蟲J2活性的測定
多媒體作為新興的科技產物,對小學生具有一定的吸引力,從而提高小學生的學習質量,提升教師的教學效果。多媒體采用音頻、視頻和教學相結合的方式,增加了課堂學習的趣味性和新穎性,激發了學生學習和求知的欲望,引起了學生濃厚的學習興趣,幫助學生更好地理解教學內容,推動了學生向更深層次的追求和發展,從而促進小學數學教學效果的提升。
取800 μL木霉發酵濾液加入內含約50條J2線蟲的貝氏小皿中,置于25 ℃恒溫箱中培養,3次重復,以無菌發酵培養液為對照,分別在12、24 h觀察J2死亡情況,若線蟲呈僵直狀態,使用0.5%NaClO刺激后無再次活動現象,則判定其為死亡[8]。
線蟲的校正死亡率公式如下:
校正死亡率(%)=

所獲數據采用Excel2013和SPSS Statistics軟件進行數據處理,Duncans新復極差法進行多重比較。
2 結果與分析
2.1 木霉的形態特征
FT85與FT1937菌株在PDA培養基上生長迅速,菌落呈圓形,輪環狀排列,初在邊緣產生白色菌絲,漸變為綠色,最后變為暗綠色。氣生菌絲較繁茂,絨氈狀,產孢量大,氣味不明顯。初生分支與主支常呈直角或略向上彎曲。瓶梗燒瓶形,形成3~4個瓶梗輪生體,偶對生;FT1937菌株與FT85相比,在平板上的顏色相對更深,呈現深綠色,分生孢子梗分支較多,產孢量大。鑒于分生孢子梗和菌落等典型形態特征,初步鑒定FT85與FT1937為哈茨木霉(TrichodermaharzianumRifai,1991)。
FT311與FT312菌株在PDA培養基上生長較快,菌絲初期為白色,后期轉變為綠色,背面無色,氣味不明顯。菌絲無色透明,分生孢子梗由菌絲直立伸出,夾角近似直角,末端有小梗,分生孢子為圓形或橢圓形;FT312菌株與FT311相比,生長相對較緩,最后菌落上不形成致密菌絲層,后期綠色不明顯,分生孢子量較少。鑒于分生孢子梗和菌落等典型形態特征,初步鑒定FT311與FT312為綠色木霉(TrichodermaviridePers,1794)。
由于木霉屬真菌在形態上差別比較難于界定,尤其是分生孢子梗和分生孢子進行形態觀察時差別較小,培養性狀觀察較為繁瑣,產生的不確定性因素較多,因此借助分子生物學技術進行DNA指紋識別鑒定將更為快速與準確。
2.2 木霉的DNA鑒定結果
將4株木霉的DNA提取后,利用ITS和β-tubulin通用引物進行PCR擴增,分別獲得580 bp左右和310 bp左右的片段(圖1和圖2),而后測序獲得其相關基因序列。
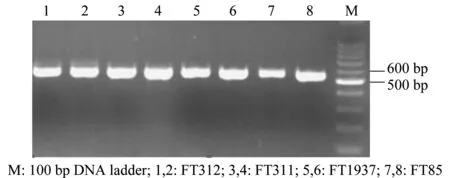
圖1 ITS序列PCR擴增產物電泳圖Fig.1 Electrophoresis of the PCR products of ITS
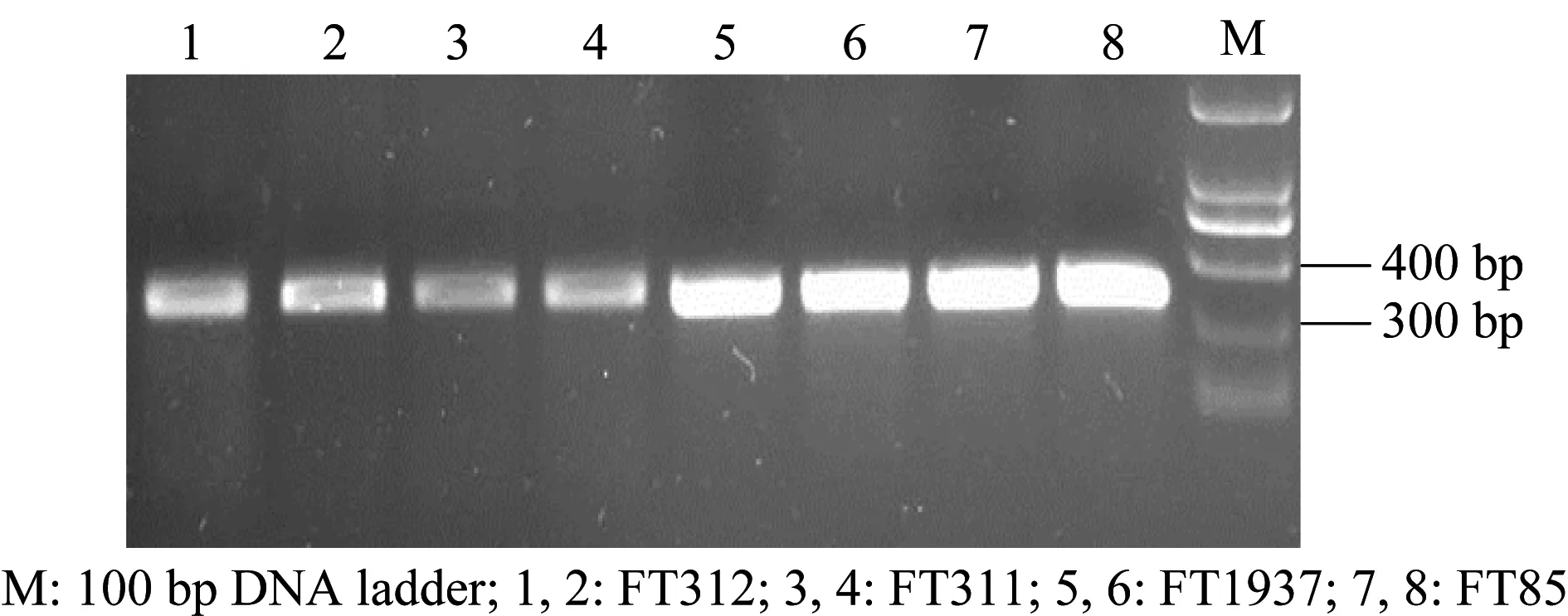
圖2 β-tubulin序列PCR擴增產物電泳圖Fig.2 Electrophoresis of the PCR products of β-tubulin
將得到的基因序列與GenBank數據庫中的已知木霉屬進行同源性搜索BLAST比對(表1),從DNA遺傳信息方面進一步確定種類鑒定的準確性。
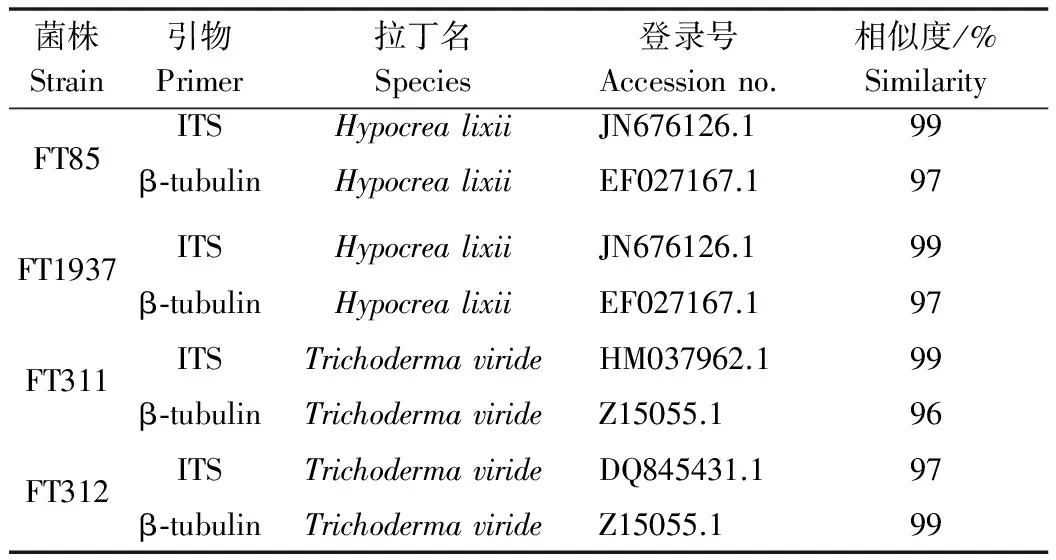
表1 木霉菌株的ITS和β-tubulin基因測序結果比對分析
FT85和FT1937菌株的ITS和β-tubulin基因測序結果顯示與哈茨木霉的有性型Hypocrealixii相似度最高,系統進化樹分析FT85和FT1937也被分為一組,結合其形態學鑒定,表型特征符合哈茨木霉的一般性狀,最終鑒定兩個菌株為哈茨木霉(Trichodermaharzianum)。
FT311和FT312菌株的ITS和β-tubulin基因測序結果顯示與綠色木霉相似度最高,系統進化樹分析FT311和FT312被分為一組,結合其形態學鑒定,表型特征符合綠色木霉的一般性狀,最終鑒定兩個菌株為綠色木霉(Trichodermaviride)。
根據4株木霉菌株的ITS和β-tubulin基因序列比對,利用MEGA軟件進行序列分析,結果(圖3)表明,FT85和FT1937菌株被分為一組,其中ITS序列與哈茨木霉(EF568084.1)同源性達到99%,但β-tubulin序列與哈茨木霉(EF027167.1)的同源性較低,僅為79%。菌株FT311和FT312被分為一組,其同源性很高,ITS序列與綠色木霉(DQ315457.1)同源性達到98%,β-tubulin序列與綠色木霉(Z15055.1)同源性達到100%。

圖3 4株木霉的基因序列遺傳聚類分析Fig.3 Phylogenetic analysis of gene sequences of Trichoderma
2.3 木霉菌株發酵液對南方根結線蟲卵孵化的影響
在木霉發酵濾液對南方根結線蟲卵孵化抑制率的測定中,處理24 h時線蟲的卵幾乎沒有孵化,從48 h開始到96 h的測試中,4株木霉均能顯著降低南方根結線蟲卵的孵化。其中菌株FT1937和FT85屬于哈茨木霉,對卵孵化的抑制效果非常明顯,48 h處理抑制率最高達92%,96 h處理抑制率降低,但也能達到80%以上。菌株FT311屬于綠色木霉,也能穩定地將卵孵化抑制率維持在80%~85%水平,但綠色木霉FT312的抑制效果一般,僅能維持在55%~60%水平(圖4)。此外,與對照相比,試驗中觀察到經木霉發酵液處理的線蟲存在卵被消解、線蟲卵孵化被延遲的現象,即使孵化出的幼蟲也有部分呈僵直狀死亡。
2.4 木霉菌株發酵液對南方根結線蟲2齡幼蟲的毒力測定
發酵液浸沒南方根結線蟲J2處理12 h,FT1937菌株對J2的毒殺作用表現突出,能達到40%的致死率;24 h后哈茨木霉FT85和FT1937菌株的致死效果相一致,達到90%左右,綠色木霉FT311菌株對J2的致死率也達到80%,這3株木霉菌株對J2都有較好的毒殺效果。但綠色木霉FT312菌株對J2致死效果不突出,24 h處理致死率僅達到30%,其對卵孵化的抑制效果也不明顯,說明不是所有的木霉菌株都對線蟲有很高的活性(圖5)。觀察到死亡線蟲體內產生空泡,體壁變薄,有的蟲體有消解現象,與正常線蟲具有明顯的差異。
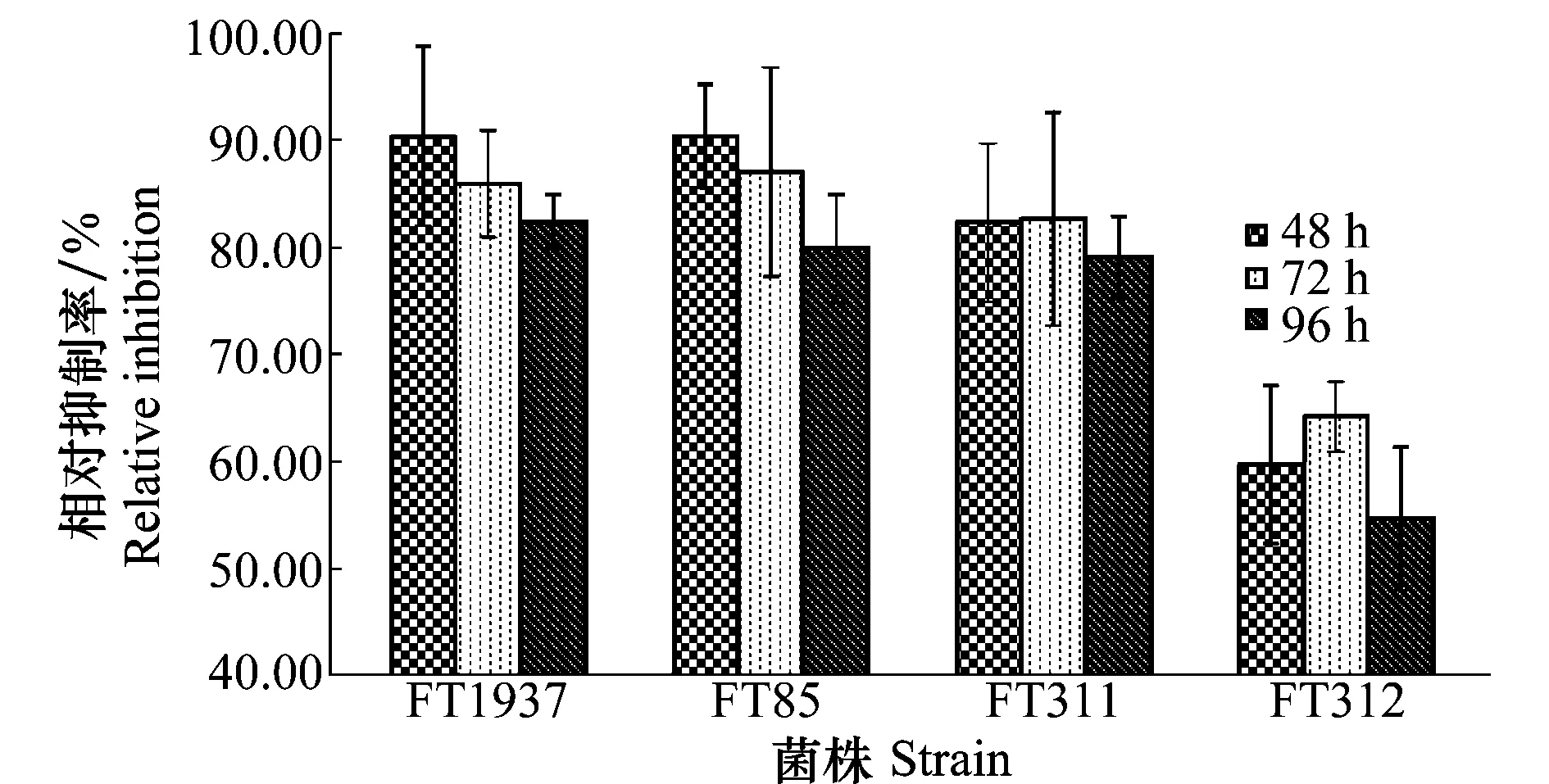
圖4 木霉對南方根結線蟲卵孵化的抑制作用Fig.4 Inhibition of Trichoderma against egg hatching of Meloidogyne incognita
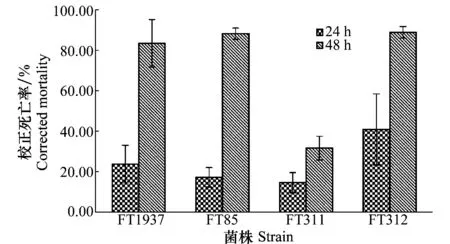
圖5 木霉菌株發酵液對南方根結線蟲J2的致死作用Fig.5 Effects of the fermentation broth of Trichoderma strains on J2 of Meloidogyne incognita
3 討論
自木霉被發現對植物病原菌有抑制作用以來,一直作為一種重要的植物病害生物防治因子而備受關注,用其防治水稻、番茄、黃瓜等的真菌病害已獲得顯著成果。近年來,人們對木霉在植物線蟲學領域的生防應用、生防機制等方面做了一些嘗試和研究,已發現多種木霉如深綠木霉(T.atrovirideP.Karsten)、哈茨木霉(T.harzianum)、絨毛木霉(T.tomentosumBissett)、綠木霉(T.virensMiller,Giddens & Foster)和棘孢木霉(T.asperellumSamuels,Lieckfeldt & Nirenberg)等均對根結線蟲具有一定的抑制作用[13-14],其中哈茨木霉和深綠木霉因其廣譜而高效的殺菌和殺線蟲活性而成為研究的熱點[15-16]。
本文對前期獲得的4株具有殺線蟲活性的木霉進行形態學和基因序列鑒定,結果顯示其中2株是哈茨木霉,另外2株是綠色木霉,而2株哈茨木霉的發酵液對南方根結線蟲卵孵化抑制作用和對J2毒性要好于綠色木霉,其中綠色木霉FT312菌株對線蟲的毒力作用較弱。說明木霉屬中不是所有種類都對植物線蟲有很強毒性,因此大量篩選獲得能夠穩定遺傳的對線蟲有作用的菌株很有必要。近幾年關于木霉對植物線蟲的作用研究進展比較迅速,已發現14種木霉對植物線蟲具有防控效果[17]。
Szabó等[18]研究表明哈茨木霉的幾丁質酶、絲氨酸蛋白酶和天冬氨酸蛋白酶等基因協同表達,在對線蟲卵寄生過程中都起重要作用。筆者在試驗過程中也觀察到南方根結線蟲的卵被消解和延遲孵化現象,部分幼蟲即使孵化出來也僵直死亡;另外2齡幼蟲被毒殺致死過程中,線蟲體壁變薄、體內產生空泡,這些現象可能是木霉的次生代謝產物中某些胞外蛋白酶的作用。Szabó等[19]研究顯示幾丁質酶和蛋白酶對線蟲的卵寄生過程中降解卵殼中幾丁質等有重要作用。
目前木霉殺線蟲的作用機理研究還較少,具體其發酵液中產生何種代謝產物起何種作用,都需要進行深入研究。隨著基因組學和蛋白質組學在農業領域的應用,木霉對植物線蟲病害的生物防治機理與應用研究將與日俱增。尤其木霉本身的競爭優勢,加之對線蟲的活性代謝產物研究深入開展,會強力推進木霉防控植物病害尤其是線蟲病害的研究與開發,并將能夠深入探討木霉與線蟲和植物的復雜互作機制。
[1] 段玉璽.植物線蟲學[M].北京:科學出版社,2011:1-9.
[2] 趙磊,段玉璽,白春明,等.遼寧省根結線蟲發生種類與分布[J].湖北農業科學,2010,49(4):865-868.
[3] Srivastava R K,Singh R K,Prasad R D.Relative antagonistic effect of different isolates ofTrichodermavirideandTrichodermaharzianumagainstRhizoctoniasolani[J].National Academy Science Letters,2012,35(1):49-52.
[4] 張麗榮,康萍芝,杜玉寧,等.木霉制劑對西瓜枯萎病的田間防治效果[J].北方園藝,2011(17):148-149.
[5] 孫虎,楊麗榮,全鑫,等.木霉生防機制及應用的研究進展[J].中國農學通報,2011,27(3):242-246.
[6] 張樹武,徐秉良,薛應鈺,等.長枝木霉對南方根結線蟲致死和寄生作用的顯微觀察及測定[J].植物保護, 2013, 39(4): 46-51.
[7] Murslain M,Javed N,Khan S A,et al.Efficacy of moringa leaves andTrichodermaharzianumon the invasion and development ofMeloidogynejavanica[J].Pakistan Journal of Phytopathology,2013,25(1):59-64.
[8] 段玉璽,靳瑩瑩,王勝君,等.生防菌株Snef85的鑒定及其發酵液對不同種類線蟲的毒力[J].植物保護學報,2008,35(2):132-136.
[9] Rifai M A.A revision of the genusTrichoderma[J].Mycological Papers,1969,116:1-56.
[10]Bissett J.A revision of the genusTrichoderma.III.SectionPachybasium[J]. Canadian Journal of Botany, 1991, 69: 2373-2417.
[11]White T J,Bruns T,Lee S,et al.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics [J].PCR protocols:a guide to methods and applications. New York: Academic Press,1990:315-322.
[12]Glass N L,Donaldson G C.Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes[J].Applied and Environmental Microbiology,1995,61(4):1323-1330.
[13]Daragó A,Szabó M,Hrács K,et al.In vitro investigations on the biological control ofXiphinemaindexwithTrichodermaspecies [J].Helminthologia,2013,50(2):132-137.
[14]Affokpon A,Coyne D L,Htay C C,et al.Biocontrol potential of nativeTrichodermaisolates against root-knot nematodes in west African vegetable production systems [J].Soil Biology and Biochemistry,2011,43:600-608.
[15]Sharon E,Bar-Eyal M,Chet I,et al.Biological control of the root-knot NematodeMeloidogynejavanicabyTrichodermaharzianum[J].Phytopathology,2001,91(7):687-693.
[16]Spiegel Y,Chet I.Evaluation ofTrichodermaspp. as a biocontrol agent against soilborne fungi and plant-parasitic nematodes in Israel [J].Integrated Pest Management Reviews,1998,3(3):169-175.
[17]Sahebani N,Hadavi N.Biological control of the root-knot nematodeMeloidogynejavanicabyTrichodermaharzianum[J].Soil Biology and Biochemistry,2008,40:2016-2020.
[18]Szabó M,Urbán P,Virányi F,et al.Comparative gene expression profiles ofTrichodermaharzianumproteases during in vitro nematode egg-parasitism[J].Biological Control,2013,67(3):337-343.
[19]Szabó M,Csepregi K,Gálber M,et al.Control plant-parasitic nematodes withTrichodermaspecies and nematode-trapping fungi:The role of chi18-5 and chi18-12 genes in nematode egg-parasitism [J].Biological Control,2012,63:121-128.
Identification of Trichoderma and its toxicity to Meloidogyne incognita
Jiao Jun,Han Bingjie,Wang Yuanyuan,Zhu Xiaofeng,Duan Yuxi,Chen Lijie
(College of Plant Protection,Shenyang Agricultural University,Nematology Institute of Northern China, Shenyang 110866, China)
Trichodermais a famous biocontrol fungus in the world. Many species of the genus play an important role in controlling plant diseases, and have important scientific value and wide application prospects. In order to identifyTrichoderma, the morphology, ITS andβ-tubulinsequences were used to investigate the four strains, FT311, FT312, FT1937 and FT85, in this study. The four strains had been screened previously as biological nematicidal factors against plant nematodes in our laboratory. The results showed that FT311 and FT312 wereTrichodermaviride, and FT1937 and FT85 wereT.harzianum. The four strains could significantly inhibit the egg hatchability and were highly toxic to the secondary juveniles (J2) ofMeloidogyneincognitawith Bayesian dish immersion method. FT1937 and FT85 had high toxicity toM.incognitawith a J2corrected mortality of about 85%-92%, and FT311 had a J2corrected mortality of about 80%-85%, but FT312 had low toxicity, with a corrected mortality of about 40%-60%. Also, the cuticle of juveniles and shell of eggs were digested, eggs hatching delayed, and the intestinal cavity of larvae showed vacuolation. Furthermore, two strains ofT.harzianumwere better thanT.viridein their activity toM.incognita. The study provides more information for the control of the root knot nematode disease withTrichodermain our country.
Trichoderma; identification;Meloidogyneincognita; nematicidal activity
2014-03-05
2014-05-21
公益性行業(農業)科研專項(201103018);國家自然科學基金(31171823);沈陽農業大學天柱山學者資助計劃
S 476
A
10.3969/j.issn.0529-1542.2015.02.011
* 通信作者 E-mail: chenlijie0210@163.com
猜你喜歡
品牌研究(2022年29期)2022-10-15 08:01:00
品牌研究(2022年26期)2022-09-19 05:54:46
品牌研究(2021年31期)2022-01-25 06:41:46
品牌研究(2021年36期)2022-01-21 09:29:28
品牌研究(2021年34期)2022-01-18 10:54:04
品牌研究(2021年35期)2022-01-18 08:45:18
品牌研究(2021年20期)2022-01-11 04:07:56
品牌研究(2021年27期)2022-01-04 06:41:14
品牌研究(2021年17期)2021-12-09 05:06:54
海峽姐妹(2019年3期)2019-06-18 10:37:10
