淡紫灰鏈霉菌除草活性物質對苘麻葉片生理生化的影響
2015-11-25 08:26:04魏松紅楊亞飛李平生藺麗文紀明山
植物保護 2015年2期
魏松紅,楊亞飛,李平生,藺麗文,東 琴,紀明山
(沈陽農業大學植物保護學院, 沈陽 110866)
?
研究報告
Research Reports
淡紫灰鏈霉菌除草活性物質對苘麻葉片生理生化的影響
魏松紅,楊亞飛,李平生,藺麗文,東 琴,紀明山
(沈陽農業大學植物保護學院, 沈陽 110866)
以苘麻成株葉片為材料,采用植物抗性生理分析測定技術研究了淡紫灰鏈霉菌除草活性物質D -13組分對苘麻葉片細胞膜透性、葉片內MAD含量、SOD、CAT和APX酶活性變化的影響,以及對膜系統的影響。結果表明,苘麻葉片經D -13組分處理后,細胞膜透性隨著D -13組分濃度的增加而增加,MDA含量在D -13組分處理后,隨著D -13組分濃度的增加而升高。在電鏡觀察下,葉片細胞的膜系統隨著D -13組分濃度的升高呈現出不同的受害情況。隨著D -13組分處理時間的延長,SOD、CAT、APX活性均出現了不同程度的下降,與對照相比,SOD、CAT、APX在40 h時活性達到最低,分別比對照活性降低超過了60%、30%和50%。可見,淡紫灰鏈霉菌除草活性物質可嚴重降低苘麻葉片的生理生化活性。
除草活性物質; 膜透性; MDA; 酶活性; 超微結構
苘麻(AbutilontheophrastiMedicus),是錦葵科(Malvaceae)苘麻屬(AbutilonMiller)一年生亞灌木狀草本,主要危害玉米、棉花、豆類、蔬菜等作物,荒地、路旁亦有生長[1]。由于其具有廣泛的適生性和較強的繁殖特性,已經成為普遍的危害物種。目前,化學除草劑在雜草防治中占有較大比重,但化學除草劑的大量使用也引發了一系列問題,諸如環境污染日趨嚴重、耐藥和抗藥雜草種群的上升、對非雜草生物的危害等。許多微生物寄主存在專一性,產生的毒素首先滲入宿主雜草,破壞其內部結構,進而使雜草產生病斑或枯萎,最終造成雜草的死亡。Kim等[2]報道不等彎孢霉產生的棘殼孢素pyrenocine A和pyrenocine B均可引起百慕大草坪的電解質滲漏和葉片尖端枯萎。從百日草鏈格孢(Alternariazinniae)內分離到的毒素zinniol,可抑制番茄、西瓜、柑橘和胡蘿卜種子萌發,引起西瓜、西葫蘆、燕麥等植物幼苗的萎蔫,可以干擾細胞的鈣素調節過程而使細胞死亡[3]。據不完全統計,被研究的生防真菌多達150余種[4]。本試驗篩選出對苘麻具除草活性的淡紫灰鏈霉菌(Streptomyceslavendulae)[5],其次級產物D -13組分可以使苘麻葉片萎蔫、干枯,研究D -13組分對苘麻葉片傷害的生理生化影響,能夠為利用淺紫灰鏈霉菌的代謝產物作為控制苘麻的生物源除草劑開發提供依據。
1 材料與方法
1.1 材料
苘麻(AbutilontheophrastiMedicus)采自沈陽農業大學實驗基地,對種子進行自然風干處理,裝入牛皮紙袋中,放置于陰涼干燥處備用。
淡紫灰鏈霉菌株從土壤中分離篩選獲得。將菌種活化,接種到改良的高氏一號培養基上,培養5 d。用正丁醇混合萃取、合并有機相。減壓濃縮,獲得粗提取物,提取物經一級、二級柱層析分離及生物測定得到活性組分D -13。
1.2 細胞膜透性的測定
選取生長一致的露白苘麻種子在溫室內培育,至幼苗長出3~4對葉時,取幼苗倒數第3對葉片。用流水沖洗30 min,再用蒸餾水清洗3次,濾紙吸干水分。切成0.5 cm×0.5 cm方塊,稱取0.5 g,浸入盛有濃度50、100、500、1 000 μg/mL的D -13組分溶液的試管中,真空減壓處理30 min,28 ℃光照下2 h,每個處理3次重復。取出葉片,用蒸餾水沖洗3次,用濾紙吸干葉片表面水分,置于盛有10 mL蒸餾水的20 mL試管中,用濾網使葉片完全浸入水中,于8、16、24、32和40 h后,在室溫下用電導儀測定葉組織浸出液的電導率,將各處理材料煮沸15 min,冷卻至室溫后測定葉片組織細胞膜徹底破壞后的最大電導率值[6],計算D -13組分對葉片組織細胞膜的傷害率。傷害率用相對電導率來表示。
1.3 丙二醛(MDA)含量的測定
葉片處理同1.2,用蒸餾水沖洗3次,用濾紙吸干葉片表面的水分,加入5 mL磷酸緩沖液(pH7.8,內含1% PVP),冰浴研磨成勻漿,于2 500 r/min離心10 min,取上清液2 mL與2 mL硫代巴比妥酸溶液(內含20%三氯乙酸,0.5%硫代巴比妥酸)混合,100 ℃水浴30 min,冷卻后于2 500 r/min離心10 min,取上清液分別在450、532和600 nm處測消光度值A450、A532和A600,按公式Y532=-0.001 98+0.088A450,MDA的消光度值D=A532-A600-Y532,求出樣品的MDA含量[6]。
1.4 細胞葉綠體超微結構的觀察
幼苗長出3~4對葉時,取幼苗倒數第3對葉片葉脈附近葉肉組織,切成1 mm×3 mm長條。2.5%戊二醛固定2 h,蒸餾水清洗3次,1%鋨酸固定2 h,PBS(二鉀砷酸鈉緩沖液配制)清洗2 h。50%、70%乙醇脫水各0.5 h,80%、90%的丙酮脫水各0.5 h,100%的丙酮脫水3次各10 min。Epon-812#環氧樹脂滲透過夜,Epon-812#環氧樹脂包埋。在35、45、60 ℃條件下依次聚合24 h,LKB-5超薄切片儀切片。醋酸雙氧鈾,醋酸鈾與檸檬酸鉛雙重染色[7]。H-7650型透射電鏡觀察。
1.5 酶活性的測定
超氧化物歧化酶(superoxide dismutase,SOD)活性測定[6]:取長勢均勻,生長至3~4對葉的苘麻葉片0.5 g于預冷的研缽中,加1 mL預冷的磷酸緩沖液在冰浴上研磨成勻漿,加緩沖液使終體積為5 mL。在4 ℃條件下10 000 r/min離心20 min,上清液即為SOD粗提液。取2.65 mL SOD反應液(1.5 mL 0.05 mol/L 磷酸緩沖液pH 7.8、0.3 mL 130 mmol/L甲硫氨酸溶液、0.3 mL 750 μmol/L氮藍四唑溶液、0.3 mL 100 μmol/L EDTA-Na2溶液、0.25 mL蒸餾水)于10 mL試管中,加入50 μL酶提取液,再加入300 μL核黃素溶液,混勻(以pH7.8磷酸緩沖液代替酶液做對照)。在4 000 lx日光下反應20 min,黑暗終止反應,于560 nm下測吸光值。SOD活性以1 g鮮重樣品1 min抑制氮藍四唑光化還原的50%(即酶單位)來表示。
過氧化氫酶(catalase,CAT)酶活性測定[8]:取各處理的苘麻葉片0.5 g置研缽中,加2~3 mL 4 ℃下預冷的pH=7.8的磷酸鹽緩沖液和少量石英砂研磨成勻漿后,轉入25 mL容量瓶中,并用緩沖液沖洗研缽數次,合并緩沖洗液,并定容到刻度。混合均勻,將容量瓶置5 ℃靜置10 min,取上部澄清液在4 000 r/min下離心15 min,上清液即為過氧化氫酶粗提液,5 ℃下保存備用。取2.5 mL酶液于50 mL三角瓶中(對照加煮死酶液2.5 mL),再加入2.5 mL 0.1 mol/L H2O2,同時計時,于30 ℃恒溫水浴中保溫10 min,立即加入10% H2SO42.5 mL用0.1 mol/L KMnO4標準溶液滴定,至出現粉紅色(在30 min內不消失)為終點。CAT酶活性用1 g鮮重樣品1 min內分解H2O2的毫克數表示。
抗壞血酸過氧化物酶(ascorbate peroxidase,APX)活性的測定:將各處理的苘麻葉片0.5 g加入提取液,研磨勻漿,15 000 r/min離心15 min,上清液定容至5 mL,取部分上清液經適當稀釋后用于酶活測定。在3 mL的反應體系中,加入50 mmol/L pH=7.0 PBS 1.8 mL,15 mmol/L AsA 0.1 mL,酶液0.1 mL,0.3 mmol/L H2O21 mL(以不加酶液為對照),記錄A290變化。以1 min內A290變0.01定義為1個酶活性單位(U)。
2 結果與分析
2.1 D -13組分對苘麻葉片細胞膜透性的影響
D -13組分對苘麻葉片細胞膜透性的影響見圖1。浸出液的相對電導率隨D -13組分濃度和處理時間的變化而增強。D -13組分溶液濃度在50~100 μg/mL的范圍內時,隨著時間的延長漲幅較小,曲線較為平緩。當D -13組分濃度超過100 μg/mL,相對電導率顯著增加時間出現在24 h左右。當D -13組分濃度達到500 μg/mL時,細胞液滲出率比對照增加35%。結果表明D -13組分對苘麻葉片細胞膜透性有影響,能夠明顯地引起葉片組織電解質的外滲。
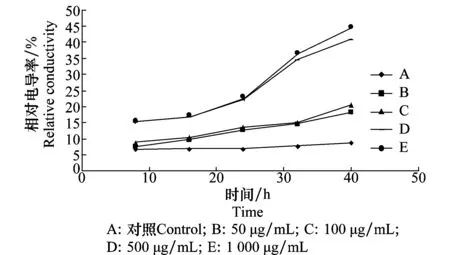
圖1 D -13組分對苘麻葉片細胞膜透性的影響Fig.1 Influences of D -13 on the permeability of cell membrane of A. theophrasti leaves
2.2 D -13組分對細胞內丙二醛(MDA)含量的影響
用D -13組分處理苘麻葉片后,細胞內MDA的含量見圖2,結果表明D -13組分處理能增加葉片組織MDA的含量,當D -13組分濃度在100 μg/mL以下時,MDA的含量上升不明顯,而D -13組分濃度在100 μg/mL以上時,MDA的含量隨D -13組分濃度的增加而上升,當D -13組分濃度達到1000 μg/mL時MDA的含量比對照增加237%。試驗結果表明D -13組分能引起苘麻葉片細胞組織內膜脂過氧化作用,導致MDA的大量滲漏。

圖2 D -13組分對苘麻葉片細胞內MDA含量的影響Fig.2 Influences of D -13 on MDA content of A. theophrasti leaves
2.3 D -13組分對苘麻葉片細胞葉綠體超微結構的影響
未經D -13組分處理的苘麻葉片細胞葉綠體分布均勻(圖3a),排列整齊,多成不規則橢圓形,基質分布均勻,膜結構清晰,基粒類囊體排列整齊緊密。經100 μg/mL D -13組分處理后,葉片的超微結構發生變化(圖3b),壁間偶有不明物質,葉綠體發生扭曲變形,基粒片層排列正常。經500 μg/mL D -13組分處理后(圖3c),可見細胞質壁分離嚴重,葉綠體膜變形受損變得模糊不清,偶有空泡化,大部分葉綠體的基質片層排列紊亂,且嗜鋨顆粒增多。經1 000 μg/mL D -13組分處理后(圖3d),葉綠體空泡化加劇,葉綠體膨脹較嚴重,基粒片層排列不規則且腫脹分解,嗜鋨顆粒進一步增多。未經D -13組分處理的苘麻葉片細胞線粒體、質膜結構正常(圖3e),線粒體基質、脊清晰可見,質膜雙層結構清晰。經500 μg/mL D -13組分處理后(圖3f~g)線粒體變形、脊數量減少,質膜出現局部破裂,內含物外流。
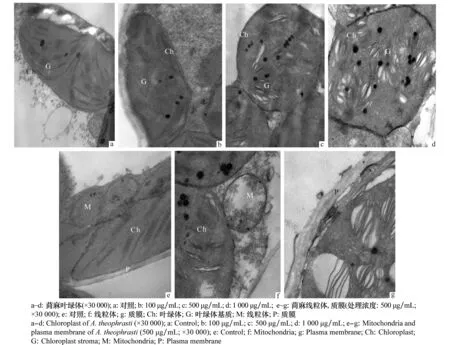
圖3 D -13組分對苘麻葉片細胞超微結構的影響Fig.3 Influences of D -13 on cell ultrastructure of A. theophrasti leaves
2.4 D -13組分對苘麻葉片酶活性的影響
D -13組分對苘麻葉片中SOD、CAT和APX酶活性的影響分別見圖4~6,結果表明濃度在500 μg/mL時,苘麻葉片中CAT、APX和SOD 3種抗氧化酶活性的變化大小和變化趨勢各不相同。作為植物抗氧化系統第一道防線的SOD,處理16 h時對其活性沒有明顯的影響,活性僅略有降低。隨著處理時間的延長,SOD活性迅速下降,40 h時降低超過60%。CAT活性在D -13組分處理的過程中其檢測值緩慢降低,前期CAT活性沒有太大波動,但在24 h時CAT活性大為降低,比對照降低32%。APX活性在處理16 h后略有升高,隨著處理時間的延長,APX的活性大幅下降,處理時間為40 h時活性大幅度降低,比對照降低51%。

圖4 D -13組分對苘麻葉片超氧化物歧化酶活性的影響Fig.4 Influences of D -13 on SOD activity of A. theophrasti leaves

圖5 D -13組分對苘麻葉片過氧化氫酶活性的影響Fig.5 Influences of D -13 on CAT activity of A. theophrasti leaves

圖6 D -13組分對苘麻葉片抗壞血酸過氧化物酶活性的影響Fig.6 Influences of D -13 on APX activity of A.theophrasti leaves
3 結論與討論
活性物質主要的作用方式有:破壞寄主細胞膜的結構、影響寄主體內正常的生理代謝等[9]。淡紫灰鏈霉菌D -13組分是病原生物代謝過程中產生的,在生理濃度范圍內干擾植物正常生理功能的化合物,可以引起苘麻葉片萎蔫、壞死。

細胞超微結構改變是植物一系列生理活動異常的細胞學基礎,研究植物受D -13組分毒害后細胞超微結構的變化,可以從細胞水平上揭示D -13組分毒害植物的機理[11],張云霞等[12]報道,用蓮子草假隔鏈格孢毒素處理空心蓮子草葉片后,葉肉細胞超微結構發生改變,具體表現為葉綠體、質膜、線粒體等細胞器結構受到傷害。用D -13組分處理苘麻葉片后葉肉細胞超微結構發生改變,表現為葉綠體、線粒體、質膜等細胞器結構受到傷害。這些結果表明D -13組分對苘麻葉片細胞膜結構具有破壞作用。
SOD是植物體內第一道防線[13],試驗中SOD活性在低D -13組分濃度處理下只稍有波動,但隨著D -13組分濃度增大,SOD活性明顯受到抑制。CAT是清除高等植物葉綠體內H2O2的重要防御酶[14],但在應對D -13組分處理中酶活性持續下降,這可能是D -13組分作用于葉綠體時產生過多的活性氧,活性氧破壞了膜系統,進而抑制了CAT的活性。APX酶在處理初期雖有短暫上升,但隨著時間延長酶活性也大幅降低。這一結果與姜述君等[15](用狹卵鏈格孢菌毒素細交鏈孢菌酮酸處理稗草)及李榮金和強勝[10](用百日草鏈格孢菌毒素處理加拿大一枝黃花葉片)所做研究葉片細胞生理生化特性的變化趨勢相似。
綜上所述,D -13組分對苘麻葉片細胞膜系統和抗氧化酶系統傷害的順序為:D -13組分處理下,苘麻葉片細胞內產生的活性氧遠大于抗氧化酶系統的清除能力,活性氧積累,進而破壞膜系統,膜透性增大,胞質外滲(表現為受害葉片呈水浸狀),胞內正常的生理代謝被打亂,最終導致細胞受到不可逆傷害,所以D -13組分對苘麻傷害的實質是對苘麻膜系統的破壞。
[1] 李揚漢.中國雜草志[M].北京:中國農業出版社,1998:694-695.
[2] Kim J C, Choi G, Heung T, et al. Pathogenicity and pyrenocine production ofCurvulariainaequalisisolated from zoysia grass [J]. Plant Disease, 2000, 84: 684-688.
[3] 李榮金. 百日草鏈格孢菌毒素的生產、分離、純化和鑒定及其作為微生物源除草劑潛力的研究[D].南京:南京農業大學,2005.
[4] 唐樹戈, 牟林, 鄭其格, 等. 玉米彎孢葉斑病菌毒素穩定性的研究[J]. 植物保護, 2009, 35(1): 111-113.
[5] 付丹妮. 具除草活性放線菌篩選及其活性物質研究[D]. 沈陽: 沈陽農業大學, 2011.
[6] 李合生. 植物生理生化實驗原理和技術[M]. 北京: 高等教育出版社, 2000.
[7] 宇克莉,孟慶敏,鄒金華.鎘對玉米幼苗生長、葉綠素含量及細胞超微結構的影響[J].華北農學報,2010,25(3):118-123.
[8] Amalo K, Chen G X, Asade K. Separate assays specific for ascorbate peroxidase and guaiacol peroxidase and for the chloroplastic and cytosolic isozymes of ascorbate peroxidase implants[J]. Plant and Cell Physiology, 1994, 35: 497-504.
[9] 萬佐璽, 朱晶晶, 強勝. 鏈格孢菌毒素對紫莖澤蘭的致病機理[J]. 植物資源與環境學報, 2001, 10(3): 47-50.[10]李榮金,強勝.百日草鏈格孢菌毒素對加拿大一枝黃花葉片傷害的生理生化研究[J].西北植物學報,2006,26(5):995-1000.
[11]張靜, 朱為民. 低溫脅迫下番茄細胞超微結構的變化[J]. 河南農業科學, 2012, 41(2): 108-110, 114.
[12]張云霞, 范蘭蘭, 施祖榮, 等. 蓮子草假隔鏈格孢毒素對空心蓮子草葉片和根尖組織超微結構的影響[J]. 華中農業大學學報, 2011, 30(1): 84-88.
[13]Gao Junjie, Li Tao, Yu Xianchang. Gene expression and activities of SOD in cucumber seedlings were related with concentrations of Mn2+, Cu2+, or Zn2+under low temperature stress[J]. Agricultural Sciences in China, 2009,8(6): 678-684.
[14]Chen Shiguo, Yin Chunyan, J?rg Reto, et al. Reactive oxygen species from chloroplasts contribute to 3-acetyl-5-isopropyltetramic acid-induced leaf necrosis ofArabidopsisthaliana[J]. Plant Physiology and Biochemistry, 2012, 52: 38-51.
[15]姜述君, 張國慶, 于涵, 等. 狹卵鏈格孢菌毒素細交鏈孢菌酮酸對稗草細胞膜脂質過氧化作用的影響[J]. 植物保護學報, 2011, 38(2): 178-182.
Physiological effects of the substances with herbicidal activity fromStreptomyceslavendulaeonAbutilontheophrastileaves
Wei Songhong, Yang Yafei, Li Pingsheng, Lin Liwen, Dong Qin, Ji Mingshan
(College of Plant Protection, Shenyang Agricultural University, Shenyang 110866, China)
The effects of the substances with herbicidal activity, D -13 components, fromStreptomyceslavendulaeon membrane permeability, MDA content, SOD, CAT and APX activities in cells ofAbutilontheophrastiMedicus leaves were studied through analyzing the resistance physiology. The results showed that the cell membrane permeability and the MDA content increased with the increase of D -13 concentration. Under the electron microscope, leaf cell membrane system showed different symptoms with the increase of D -13 concentration. SOD, CAT and APX activities declined to different degrees with the extension of processing time of D -13 components. Compared with the controls, SOD, CAT and AXP activities were the lowest at 40 h, decreasing more than 60%, 30% and 50%, respectively, compared with the control. It is concluded that D -13 components could effectively depressed the physiological activities ofA.theophrastileaves.
substance with herbicidal activity; membrane permeability; MDA; enzyme activity; ultramicrostructure
2014-02-25
2014-05-11
公益性行業(農業)科研專項(201303031)
S 451,S 476
A
10.3969/j.issn.0529-1542.2015.02.004
聯系方式 E-mail: songhongw125@163.com
