鴕鳥源鼠隱孢子蟲蟲種鑒定及動(dòng)物感染試驗(yàn)研究
2015-11-20 11:42:00劉利敏徐利納張素梅張龍現(xiàn)
中國人獸共患病學(xué)報(bào) 2015年5期
關(guān)鍵詞:小鼠
齊 萌,劉利敏,黃 磊,徐利納,張素梅,張龍現(xiàn)
隱孢子蟲病(Cryptosporidiosis)是由隱孢子蟲(Cryptosporidium)引起的一種重要的機(jī)會(huì)性人獸共患原蟲病,為全球性重要的食源性/水源性疾病之一[1]。隱孢子蟲已公認(rèn)的有效種有27個(gè),多數(shù)種類具有宿主特異性,其中鼠隱孢子蟲(Cryptosporidiummuris)為人獸共患蟲種,主要寄生于嚙齒類動(dòng)物胃腺粘膜上皮細(xì)胞,包括人在內(nèi)的其他多種哺乳動(dòng)物也有感染報(bào)道[1-4]。禽類中,曾有報(bào)道在茶色蟆口鴟和夜鷹糞便中檢測到C.muris,但是否為糞便被C.muris卵囊污染尚不確定[5]。也有調(diào)查在蛇類糞便中檢測到C.muris和泰澤隱孢子蟲(Cryptosporidiumtyzzeri),調(diào)查者認(rèn)為該兩種隱孢子蟲并非寄生于蛇消化道上皮細(xì)胞,而是蛇攝食隱孢子蟲陽性的嚙齒類動(dòng)物,卵囊通過糞便排出體外[6-7]。
非洲鴕鳥屬鳥綱鴕形目鴕鳥科,是現(xiàn)存體型最大的平胸鳥類,其主要感染貝氏隱孢子蟲(Cryptosporidiumbaileyi)、火雞隱孢子蟲(Cryptosporidiummeleagridis)、隱孢子蟲禽基因型II(avian genotype II)和一個(gè)隱孢子蟲未定種[8-9]。目前已有的研究發(fā)現(xiàn)C.muris僅感染哺乳動(dòng)物,本研究從非洲鴕鳥糞便中分離到一種隱孢子蟲卵囊,經(jīng)形態(tài)學(xué)觀察和分子生物學(xué)鑒定為C.muris。為了解鴕鳥源C.muris傳播能力及生物學(xué)特征,本試驗(yàn)用鴕鳥源C.muris接種羅曼雛雞、Balb/c鼠和蒙古沙鼠,觀察動(dòng)物感染后臨床癥狀和排卵囊規(guī)律,通過組織學(xué)觀察其宿主適應(yīng)性和寄生部位等方面,以期為隱孢子蟲的分類地位及生物學(xué)特性研究提供參考。
1 材料與方法
1.1 材料
1.1.1 卵囊來源C.muris卵囊分離自鄭州某鴕鳥養(yǎng)殖場10歲以上長期排卵囊的鴕鳥。
1.1.2 試驗(yàn)動(dòng)物與分組 購自鄭州牧業(yè)工程高等專科學(xué)校孵化場1日齡羅曼雛雞20只,10只為感染組,10只為不感染對照組;購自河南省試驗(yàn)動(dòng)物中心20日齡Balb/c鼠15只,10只為感染組,5只為不感染對照組;購自首都醫(yī)科大學(xué)15只20日齡蒙古沙鼠,10只為感染組,5只為不感染對照組。各試驗(yàn)動(dòng)物均于感染前連續(xù)糞檢3d,無腸道寄生蟲感染者用于本試驗(yàn)。感染組動(dòng)物每只接種卵囊劑量為1.0×106個(gè)/mL,不感染對照組每只經(jīng)口灌胃等體積蒸餾水。動(dòng)物接種卵囊后每天收集糞便檢查卵囊。
1.1.3 動(dòng)物組織樣品 感染成功的動(dòng)物于排卵囊高峰期分別剖殺,取其胃、十二指腸和空腸組織樣品,光鏡樣品置10%中性福爾馬林固定液中,常溫保存?zhèn)溆茫婄R樣品保存于pH=7.2的2.5%戊二醛溶液中固定,置4。C冰箱保存。
1.2 方法
1.2.1 卵囊檢查 采用飽和蔗糖溶液漂浮法和改良抗酸染色法檢查卵囊,用微分干涉差顯微鏡進(jìn)行拍照。
1.2.2 卵囊收集與計(jì)數(shù) 采用不連續(xù)蔗糖密度梯度離心法收集卵囊。用血細(xì)胞計(jì)數(shù)板進(jìn)行卵囊計(jì)數(shù),計(jì)算每克糞便中的隱孢子蟲卵囊數(shù)(OPG)。
1.2.3 DNA提取 采用日本TOYOBO公司生產(chǎn)的Mag Extractor-Genome提取試劑盒 (購自北京夏斯生物技術(shù)有限公司),按照其說明書提取流程進(jìn)行操作。
1.2.4 PCR擴(kuò)增 18SrRNA基因引物設(shè)計(jì)和反應(yīng)程序參照Xiao等[10]的報(bào)道,引物由上海生工生物工程有限公司合成。
1.2.5 測序 由北京諾賽基因有限公司完成。
1.2.6 組織樣品觀察 光鏡樣品按常規(guī)石蠟組織切片樣品處理方法脫水、HE染色、顯微鏡觀察照相;電鏡樣品按常規(guī)掃描電鏡樣品處理方法脫水、置換乙醇、干燥和噴金,觀察照相。
2 結(jié) 果
2.1C.muris形態(tài)學(xué)觀察 飽和蔗糖溶液中卵囊呈長橢圓形,淡藍(lán)色,卵囊周圍顯黃綠色,部分卵囊囊壁凹陷。微分干涉差顯微鏡下,卵囊呈卵圓形或近圓形,測量76個(gè)卵囊,平均卵囊大小為(7.41±0.12)μm×(5.70±0.07)μm,卵囊指數(shù)為1.30。見圖1。

圖1 微分干涉顯微鏡下的鼠隱孢子蟲卵囊 (比例尺:10 μm)Fig.1 Cryptosporidium muris oocysts under differential interference contrast microscopy(Scale bars:10μm)
2.2 18SrRNA基因的PCR擴(kuò)增和測序 PCR成功擴(kuò)增出隱孢子蟲分離株的18SrRNA基因片段,大小為830~850bp。將PCR進(jìn)行測序,對其序列進(jìn)行Clustalx比對,與C.muris同源性為100%,鑒定出該隱孢子蟲分離株為C.muris。該序列GenBank登錄號(hào)為GQ227706。
2.3 動(dòng)物感染試驗(yàn) 雛雞接種鴕鳥源C.muris30d內(nèi),各組雛雞均無C.muris卵囊排出,精神狀態(tài)始終良好,剖檢其胃腸道均正常。C.muris卵囊經(jīng)灌胃接種Balb/c鼠,從感染后第9 d(9days post infection,DPI 9)有卵囊排出至DPI 46結(jié)束。潛隱期為8d,顯露期為37d。排卵囊高峰期在DPI 17-DPI 31,最大 OPG為20.5×104個(gè)。感染組與不感染對照組小鼠均無死亡,糞便性狀、食欲、毛質(zhì)和精神狀況良好。詳見圖2。

圖2 Balb/c鼠感染鼠隱孢子蟲排卵囊規(guī)律Fig.2 Development of oocysts shedding of Balb/c mice after inoculation with C.muris oocysts
C.muris卵囊接種蒙古沙鼠后,潛隱期為13 d,DPI 13發(fā)現(xiàn)卵囊排出,沙鼠持續(xù)排出卵囊時(shí)間至DPI 85結(jié)束,顯露期72d,排卵囊高峰期在DPI 31-DPI 64,最大OPG為41.5×104個(gè)。試驗(yàn)前后,感染組與不感染對照組蒙古沙鼠均無死亡,糞便性狀、食欲、毛質(zhì)和精神狀況良好。
2.4 組織粘膜涂片改良抗酸染色觀察 分別在Balb/c鼠和蒙古沙鼠感染隱孢子蟲排卵囊高峰期,各剖殺兩只,取胃腸道粘膜涂片,采用改良抗酸染色法進(jìn)行觀察,十二指腸、空腸、回腸、結(jié)腸和直腸各腸道粘膜涂片中均未發(fā)現(xiàn)C.muris卵囊,僅在鼠胃粘膜涂片中發(fā)現(xiàn)C.muris卵囊。在1 000倍光學(xué)顯微鏡下觀察,卵囊在藍(lán)綠色背景下呈玫瑰紅色,對比明顯。見圖3。
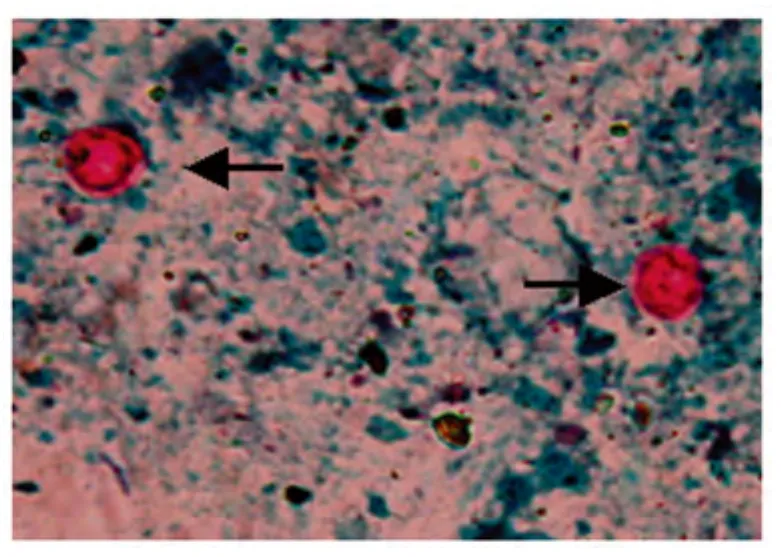
圖3 經(jīng)改良抗酸染色法染色后的鼠胃粘膜涂片中鼠隱孢子蟲卵囊(1 000×)Fig.3 C.muris oocyst in the mice gastric gland stained by modified acid fast stain(1 000×)
2.5 病理組織學(xué)觀察 剖檢可見,感染組Balb/c鼠和蒙古沙鼠的胃和對照組相比,均萎縮變小,內(nèi)容物少,胃壁變薄。通過對組織切片HE染色觀察,結(jié)果在胃腺小凹發(fā)現(xiàn)大量C.muris蟲體寄生,在胃腺內(nèi),可觀察到C.muris各發(fā)育階段,腸道組織內(nèi)未發(fā)現(xiàn)蟲體,見圖4。對照組未見異常。
2.6 電子掃描顯微鏡觀察 取感染組Balb/c鼠胃組織進(jìn)行電鏡觀察,在5 000倍掃描電鏡下,胃腺部位發(fā)現(xiàn)大量各階段C.muris蟲體寄生,帶蟲空泡嵌在胃腺粘膜內(nèi),蟲體在帶蟲空泡內(nèi)發(fā)育,Ⅰ型裂殖體8個(gè)裂殖子清晰可見,見圖5。
3 討 論
3.1 鴕鳥自然感染的隱孢子蟲蟲種 1994年,Gajadhar[11]報(bào)道鴕鳥可感染隱孢子蟲,顯微鏡觀察其卵囊形態(tài)不同于C.baileyi和C.meleagridis,分離的隱孢子蟲卵囊不能感染乳鼠、雛雞、火雞和鵪鶉,提出該鴕鳥源隱孢子蟲可能是一個(gè)新種。2006年,Meireles等[12]報(bào)道其發(fā)現(xiàn)的鴕鳥隱孢子蟲分離株與C.baileyi分子遺傳特征有差異,但需對其卵囊的形態(tài)特征和生物學(xué)特性進(jìn)行研究以確定其是否為新的隱孢子蟲種類/基因型。國內(nèi),孫銘飛等[13]對鴕鳥源隱孢子蟲分離株進(jìn)行生物學(xué)特性研究,結(jié)果顯示獲得的分離株均為C.baileyi,可感染雛雞和幼齡鴕鳥,但不能感染小鼠,主要寄生部位為泄殖腔和法氏囊。Wang等[14]報(bào)道鄭州地區(qū)鴕鳥隱孢子蟲感染率為11.7%,主要感染6月齡以下鴕鳥,基于18srRNA基因?qū)?3個(gè)隱孢子蟲分離株進(jìn)行鑒定,均為C.baileyi。本研究從形態(tài)學(xué)觀察,分子生物學(xué)方法和動(dòng)物感染試驗(yàn)證明分離的鴕鳥源隱孢子蟲為C.muris,且長期排卵囊,其在鴕鳥體內(nèi)的寄生部位有待于進(jìn)一步研究。
3.2C.muris人獸共患性C.muris具有較為廣泛的宿主譜,可感染人、小鼠、倉鼠、松鼠、田鼠、巖貍、雙峰駝、野生白山羊、犬、貓、山狗、非洲蹄兔、短尾猴和豬等多種哺乳動(dòng)物,在肯尼亞、泰國、法國、秘魯、印度尼西亞和埃及等國的免疫力低下人群中及健康人群中均有感染C.muris的報(bào)道[1-4]。先前研究認(rèn)為,C.muris只感染哺乳動(dòng)物,禽類曾有報(bào)道糞便中發(fā)現(xiàn)C.muris,但不清楚是否為通過卵囊機(jī)械傳播造成的[5]。基于多位點(diǎn)序列分型(MLST),本研究中的鴕鳥源C.muris亞型結(jié)構(gòu)(M5、M4、M6、M4)顯著不同于來自于捷克共和國、肯尼亞、埃及、秘魯、日本和中國的多種人源/動(dòng)物源C.muris亞型結(jié)構(gòu),表明鼠隱孢子蟲與宿主存在共進(jìn)化,其人獸共患性經(jīng)長期宿主適應(yīng)存在協(xié)同進(jìn)化[15-16]。
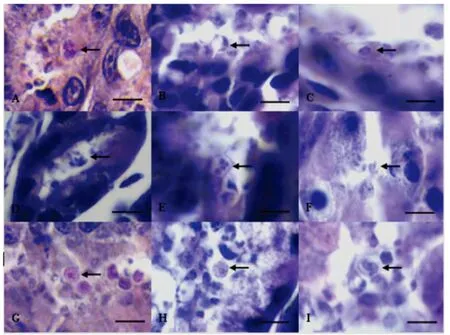
圖4 蒙古沙鼠胃腺中鼠隱孢子蟲各階段蟲體形態(tài)(1 000×)Fig.4 Morphological characterization of C.muris developed in Mongolian gerbils gastric gland(1 000×)

圖5 掃描電鏡觀察Balb/c鼠胃腺寄生的鼠隱孢子蟲(5 000×)Fig.5 C.muris developed in Balb/c mice gastric gland observed by SEM (5 000×)
3.3C.muris動(dòng)物感染試驗(yàn)C.muris由Tyzzer于1907年首次在試驗(yàn)鼠胃內(nèi)發(fā)現(xiàn)并命名,隨后研究發(fā)現(xiàn)C.muris所有發(fā)育階段均在胃腺完成[17]。日本研究人員先后用家鼠源C.muris分離株(RN66株)進(jìn)行動(dòng)物感染試驗(yàn),但結(jié)果存在較大差異。1989年,Iseki[18]等報(bào)道 RN66株卵囊感染小鼠、貓、豚鼠、兔子和犬,試驗(yàn)動(dòng)物個(gè)體接種卵囊量均為1.0×106個(gè),卵囊接種3周齡的SPF小鼠后,潛隱期為5 d,顯露期為DPI 34-75,高峰期為DPI 16-26,在高峰期每只小鼠每天排卵囊量達(dá)11×106~46×106個(gè),寄生部位為胃腺。1999年,Taylor等[19]等分別用400個(gè)、2.0×104個(gè)和1.0×106個(gè) RN66株卵囊感染6周齡小鼠,卵囊潛隱期為10~18d,排卵囊持續(xù)至DPI 89,不同劑量感染組小鼠均未表現(xiàn)臨床癥狀,感染組小鼠和不感染對照組體重變化無統(tǒng)計(jì)學(xué)差異;剖檢可見,胃腺腫大增生,充血,組織學(xué)觀察發(fā)現(xiàn)蟲體寄生于胃腺,細(xì)胞壞死,粘膜固有層嗜酸性粒細(xì)胞浸潤、變形。2001年,Toshihiro等[20]用低劑量RN66株卵囊對3周齡ICR小鼠進(jìn)行感染試驗(yàn),25只小鼠接種24個(gè)卵囊,僅1只發(fā)現(xiàn)有卵囊排出,潛隱期為21d;25只小鼠接種240個(gè)卵囊,19只感染成功,潛隱期為14~17d;50只小鼠每天接種24個(gè)卵囊,連續(xù)接種10d,4只感染成功,潛隱期為14-19d;而5只接種2.4×106個(gè)卵囊的小鼠均感染成功,潛隱期為6~7d。上述研究表明,使用同一個(gè)C.muris分離株進(jìn)行動(dòng)物感染試驗(yàn),不同的試驗(yàn)動(dòng)物和不同的感染量均可使其感染力、潛隱期及顯露期不同。
不同C.muris分離株存在宿主特異性。自然條件下,野生田鼠可感染C.muris,然而 Modry等[21]用C.muris卵囊人工接種5種田鼠,每只接種1.0×105個(gè)卵囊,結(jié)果發(fā)現(xiàn)均不感染。同樣,非人靈長類可自然感染C.muris,而近來一個(gè)研究發(fā)現(xiàn),免疫力正常和免疫抑制的食蟹猴接種1.0×107個(gè) RN66株卵囊,均未發(fā)現(xiàn)感染。Kvac等[23-24]人工感染試驗(yàn)顯示,家鼠源C.muris不能感染8周齡仔豬,但可感染蒙古沙鼠,接種7日齡和8周齡蒙古沙鼠,每只接種1.0×106個(gè)卵囊,潛隱期分別為18d和21d,前者顯露期大于90d,排卵囊最大OPG值為12×106,后者顯露期為93~100d,個(gè)體每日排卵囊最大OPG值為1.7×106,試驗(yàn)過程中,均未出現(xiàn)臨床癥狀。Melicherova等[25]用兩種實(shí)驗(yàn)鼠對C.muris進(jìn)行了生活史研究,分別接種Balb/c鼠和白鼻乳鼠1.0×108個(gè)C.muris分離株(TS03株),兩者潛隱期和顯露期存在較大差異,Balb/c鼠潛隱期和顯露期分別為7.5~10d和10~15d,而白鼻乳鼠潛隱期長達(dá)18~21d,顯露期呈慢性長期感染。內(nèi)生發(fā)育過程研究顯示,兩種實(shí)驗(yàn)鼠均于接種C.muris卵囊后5d形成厚壁形卵囊,在Balb/c鼠胃內(nèi)薄壁型卵囊和卵囊最早形成于DPI 5和DPI 7.5,而顯著不同的是,在白鼻乳鼠胃內(nèi)薄壁型卵囊和卵囊最早形成于DPI 14和DPI 18,進(jìn)一步表明不同源C.muris分離株存在宿主差異并與宿主共進(jìn)化。
本試驗(yàn)采用鴕鳥源C.muris感染雛雞、Balb/c鼠和蒙古沙鼠,感染量均為1.0×106個(gè)卵囊,發(fā)現(xiàn)雛雞不能感染C.muris,Balb/c鼠和蒙古沙鼠均可感染成功。其中Balb/c鼠感染C.muris卵囊后,潛隱期8d,顯露期37d,排卵囊高峰期在DPI 17-DPI 31;蒙古沙鼠的潛隱期為12d,顯露期72d,排卵囊高峰期在DPI 31-DPI 64;兩種實(shí)驗(yàn)鼠的感染差異同Melicherova等[25]的研究,但接種鴕鳥源C.muris分離株的Balb/c鼠顯露期明顯長于接種TS03株的Balb/c鼠,表明鴕鳥源C.muris分離株與TS03株存在不同感染情況。本試驗(yàn)中感染成功的動(dòng)物試驗(yàn)前后精神狀態(tài)良好,無臨床癥狀,組織學(xué)和電鏡掃描觀察,僅在胃內(nèi)發(fā)現(xiàn)大量C.muris蟲體寄生,并可見其各發(fā)育階段,與前人研究相一致。結(jié)果表明鴕鳥源C.muris與鼠源C.muris感染實(shí)驗(yàn)鼠后在其排卵囊規(guī)律各有差異,臨床表現(xiàn)、寄生部位、內(nèi)生發(fā)育過程相同,存在人獸共患風(fēng)險(xiǎn),但其如何寄生于鴕鳥及其在鴕鳥體內(nèi)寄生部位和繁殖發(fā)育階段有待進(jìn)一步研究。
[1]Xiao L.Molecular epidemiology of cryptosporidiosis:An update[J].Exp Parasitol,2010,124(1):80-89.DOI:10.1016/j.exppara.2009.03.018
[2]Ren X,Zhao J,Zhang L,et al.Cryptosporidiumtyzzerin.sp.(Apicomplexa:Cryptosporidiidae)in domestic mice(Musmusculus)[J].Exp Parasitol,2012,130(3):274-281.DOI:10.1016/j.exppara.2011.07.012
[3]Kvac M,Hofmannova L,Hlaskova L,et al.Cryptosporidium erinacein.sp.(Apicomplexa:Cryptosporidiidae)in hedgehogs[J].Vet Parasitol,2014,201(1-2):9-17.DOI:10.1016/j.vetpar.2014.01.014
[4]Pavlasek I,Ryan U.The first finding of a natural infection ofCryptosporidiummurisin a cat[J].Vet Parasitol,2007,144(3-4):349-352.
[5]Ng J,Pavlasek I,Ryan U.Identification of novelCryptosporidiumgenotypes from avian hosts[J].Appl Environ Microbiol,2006,72(12):7548-7553.
[6]Richter B,Nedorost N,Maderner A,et al.Detection ofCryptosporidiumspecies in feces or gastric contents from snakes and lizards as determined by polymerase chain reaction analysis and partial sequencing of the 18Sribosomal RNA gene[J].J Vet Diagn Invest, 2011, 23 (3): 430-435. DOI: 10.1177/1040638711403415
[7]da Silva DC,Paiva PR,Nakamura AA,et al.The detection ofCryptosporidiumserpentisin snake fecal samples by real-time PCR[J].Vet Parasitol,2014,204(3-4):134-138.DOI:10.1016/j.vetpar.2014.05.012
[8]Fayer R.Taxonomy and species delimitation inCryptosporidium[J].Exp Parasitol,2010,124(1):90-97.DOI:10.1016/j.exppara.2009.03.005
[9]Nguyen ST,F(xiàn)ukuda Y,Tada C,et al.Prevalence and molecular characterization ofCryptosporidiumin ostriches(Struthiocamelus)on a farm in central Vietnam[J].Exp Parasitol,2013,133(1):8-11.DOI:10.1016/j.exppara.2012.10.010
[10]Xiao L,Escalante H,Yang CF,et al.Phylogenetic analysis ofCryptosporidiumparasites based on the small-subunit rRNA gene locus[J].Appl Environ Microbiol,1999,65(4):1578-1583.
[11]Gajadhar AA.Host specificity studies and oocyst description of aCryptosporidiumsp.isolated from ostriches[J].Parasitol Res,1994,80(4):316-319.
[12]Meireles MV,Soares RM,dos Santos MM,et al.Biological studies and molecular characterization of aCryptosporidiumisolate from ostriches(Struthiocamelus)[J].J Parasitol,2006,92(3):623-626.
[13]Sun MF,Zhang LX,Ning CS,et al.Identification ofCryptosporidaisolate from ostrich in China and biological characters studies[J].Sci Agric Sin,2007,40(7):1528-1534.(in Chinese)孫銘飛,張龍現(xiàn),寧長申,等.鴕鳥源隱孢子蟲的種類鑒定及其生物學(xué)特性研究[J].中國農(nóng)業(yè)科學(xué),2007,40(7):1528-1534.
[14]Wang R,Qi M,Jingjing Z,et al.Prevalence ofCryptosporidiumbaileyiin ostriches(Struthiocamelus)in Zhengzhou,China[J].Vet Parasitol,2011,175(1/2):151-154.DOI:10.1016/j.vetpar.2010.10.005
[15]Feng Y,Yang W,Ryan U,et al.Development of a multilocus sequence tool for typingCryptosporidiummurisandCryptosporidiumandersoni[J].J Clin Microbiol,2011,49(1):34-41.DOI:10.1128/JCM.01329-10
[16]Wang R,Jian F,Zhang L,et al.Multilocus sequence subtyping and genetic structure ofCryptosporidiummurisandCryptosporidiumandersoni[J].PLoS One,2012,7(8):e43782.DOI:10.1371/journal.pone.0043782
[17]Tyzzer E.An extracelluar coccidium,Cryptosporidiummurisof the gastric glands of the common mouse[J].Med Res,1910,18:487-509.
[18]Iseki M,Maekawa T,Moriya K,et al.Infectivity ofCryptosporidiummuris(strain RN 66)in various laboratory animals[J].Parasitol Res,1989,75(3):218-222.
[19]Taylor MA,Marshall RN,Green JA,et al.The pathogenesis of experimental infections ofCryptosporidiummuris(strain RN 66)in outbred nude mice[J].Vet Parasitol,1999,86(1):41-48.
[20]Mstsue T,F(xiàn)ujino T,Kajima J,et al.Infectivity and oocyst excretion patterns ofCryptosporidiummurisin slightly infected mice[J].J Vet Med Sci,2001,63(3):319-320.
[21]Modry D,Hofmannova L,Antalova Z,et al.Variability in susceptibility of voles(Arvicolinae)to experimental infection withCryptosporidiummurisandCryptosporidiumandersoni[J].Parasitol Res,2012,111(1):471-473.DOI:10.1007/s00436-012-2821-1
[22]Masuno K,F(xiàn)ukuda Y,Kubo M,et al.Infectivity ofCryptosporidiumandersoniandCryptosporidiummuristo normal and immunosuppressive cynomolgus monkeys[J].J Vet Med Sci,2014,76(2):169-172.
[23]Kvac M,Kestranova M,Kvetonova D,et al.Cryptosporidium tyzzeriandCryptosporidiummurisoriginated from wild West-European house mice(Mus musculus domesticus)and East-European house mice(Musmusculusmusculus)are non-infectious for pigs[J].Exp Parasitol,2012,131(1):107-110.DOI:10.1016/j.exppara.2012.03.016
[24]Kvac M,Sak B,Kvetonova D,et al.Infectivity of gastric and intestinalCryptosporidiumspecies in immunocompetent Mongolian gerbils (Merionesunguiculatus)[J].Vet Parasitol,2009,163(1-2):33-38.DOI:10.1016/j.vetpar.2009.03.047
[25]Melicherova J,Ilgova J,Kvac M,et al.Life cycle ofCryptosporidiummurisin two rodents with different responses to parasitization[J].Parasitology,2014,141(2):287-303.DOI:10.1017/S0031182013001637
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
