錢塘江河口浮游植物分布與環境因子的典范對應分析
2015-11-16 09:07:44馬永星張波濤臧家業張愛軍劉煒
海洋開發與管理 2015年6期
馬永星,張波濤,臧家業,張愛軍,劉煒
(1.青島大學化學科學與工程學院 青島 266071;2.國家海洋局第一海洋研究所海洋生態研究中心 青島 266061;3.青島市地震監測中心 青島 266000)
1 引言
世界上大多數河口都受到了不同程度的人類活動干擾,導致水體富營養化程度不斷加劇,引起河口生態環境的退化甚至崩潰[1-2]。在河口區的生態環境能量與物質的流動中,河口浮游植物是河口水生態系統的初級生產者和食物鏈的基礎環節,其種類和數量的變化對環境變化具有指示作用,環境因子的改變也會影響浮游植物的群落結構,進而影響河口區生態系統的結構以及功能[3-4]。
錢塘江河口位于杭州灣內,面向東海,受到錢塘江沖淡水、長江沖淡水和潮汐的共同影響,屬于典型的河口型海灣[5],且浮游植物以硅藻為主[6]。近年來由于人為的干擾與損害,杭州灣已經成為污染嚴重的區域[7-9]。然而,目前,大多研究關注在長江口和東海水域,而對于杭州灣的浮游植物相關的研究較少。事實上,除長江口作用外,東海近岸的赤潮高發區顯然也與杭州灣的貢獻息息相關。因此,對于杭州灣浮游植物分布的變化規律及與環境因子典范對應分析的研究,將有助于認識杭州灣生態環境的發展趨勢,為杭州灣乃至東海的生態環境的保護與修復提供有力的依據。
2 材料與方法
2.1 樣品的采集
2013年5 月、2013 年8 月、2013 年11 月和2014 年1 月在錢塘江河口分別進行了春、夏、秋、冬4個季節的海洋科學綜合調查,調查范圍覆蓋杭州灣大部分區域,調查站位如圖1所示。
調查中,用Niskin采水器采集水樣,同時用多參數水質剖面儀(JFE,AAQ122)獲得水體溫度和鹽度等水文參數。調查項目包括:總懸浮顆粒物(SPM)、營養鹽和葉綠素a等。浮游植物的采樣按《海洋調查規范》(GB 17378.7—2007)進行,水采樣品:采集表層水樣500 mL,裝入聚丙烯瓶中,加1%魯哥試劑固定、沉淀和濃縮;網采樣品:使用淺水Ⅲ型浮游生物網自海底至表層垂直拖網,樣品經5%福爾馬林溶液固定保存。
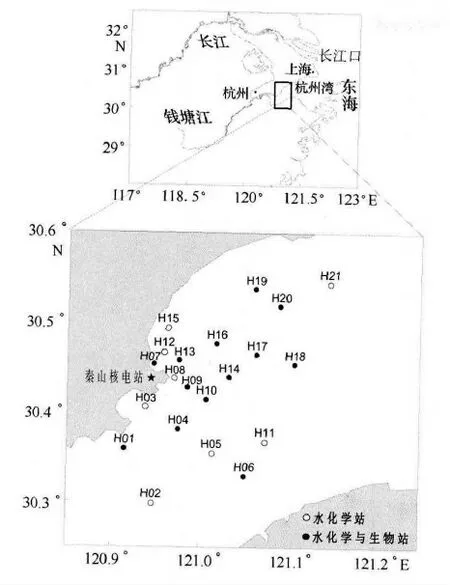
圖1 錢塘江河口調查站位圖
2.2 樣品的分析
營養鹽利用營養鹽自動分析儀(SEAL 公司,QuAAtro型)進行分析,對于1~10μmol/L含量的樣品,分析不確定性為 小于5%~10%,大于10μmol/L的樣品含量分析不確定性為1%~5%[10];葉綠素a用1∶9丙酮提取后,采用熒光法分析。浮游植物采樣結束后在實驗室內進行鏡檢分析,室內分析鑒定按《海洋調查規范》中規定的方法進行。
3 結果和討論
3.1 溫、鹽及其他水化學因子的分布特征
調查海區夏季水溫為全年最高,冬季的水溫為全年最低,各季節水溫變化由高到低依次排列為夏季、春季、秋季、冬季。由圖2 可以看出,春季表層海水溫度在東南側近岸水域水溫比較高,并沿監測水域兩岸向中心方向呈降溫的趨勢;夏季表層海水溫度分布與春季相似;秋季表層海水溫度整體上呈現從核電附近到離岸逐漸降低的趨勢,低值區位于調查海域的北部;冬季上游方向海水溫度相對較低,而入海口以及上游處水溫相對較高。
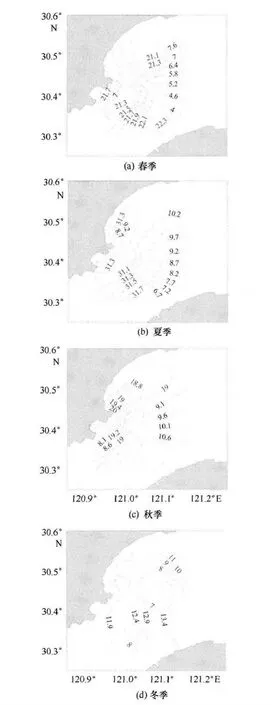
圖2 錢塘江河口溫度(℃——)和鹽度(------)的分布
整體上,表層水溫相對較高,底層水溫相對較低,且各個季節表層和底層水溫相差不大,同一站位垂直分布差異也不明顯;不過,表層和底層水溫平面分布有明顯的變化趨勢,均呈現與岸線平行的帶狀分布趨勢。水溫一定程度上受到核電溫排水的影響,各季節核電附近水溫相對較高。同時,該海域水溫還受到杭州灣海水涌入的影響,如冬季,入海口方向水溫相對較高。大多季節溫度的分布也受到了東南側陸地的影響,近岸溫度相對離岸較高。可見,調查海域各季節水溫的分布受溫排水、海水入侵以及地形的影響較大。
各季節表層鹽度的水平分布見圖2,從圖2中可以看出,春季水體表層鹽度分布趨勢為從入海口向上游方向逐漸降低;夏季表層水體的鹽度分布與春季類似,鹽度的高值出現在監測海域東北部入海口處;秋季呈均勻帶狀分布,海水鹽度自核電站沿岸海域到離岸海域逐漸上升;冬季表層水體鹽度分布呈現從上游向入海口方向逐漸增高的趨勢。
整體上表層鹽度相對較低,底層相對較高,各個季節表底層鹽度數值相差不大,垂直分布差異也不明顯。除了冬季,其他季節表層和底層鹽度平面分布相似,且普遍呈現從上游向入海口方向逐漸增高的趨勢。
總體而言,海水鹽度春季和夏季相對較低,主要是上流徑流量在這兩個時期相對較高,淡水稀釋作用更明顯。調查海域鹽度的分布與變化主要取決于江河入海徑流量的多少以及海水上涌的強度,因此,調查水域鹽度會明顯地受到潮汐作用的影響。春季和夏季上游流域雨量較大,鹽度相對較低;入秋以后,降雨量減小,徑流減小,水體鹽度增加,使得鹽度的分布更多地受到潮汐與海水涌入的影響。
調查海區水化學因子的統計見表1。總體來看,調查海域海水理化參數溫度、鹽度、懸浮物等調查結果由于受到河流和海洋的雙重影響,具有顯著的河口特性,波動性大,懸浮物濃度和濁度處于一個較高的水平,水體透明度因此較低。分析營養鹽測量結果,該水域富營養化嚴重,尤其是無機氮和磷酸鹽的濃度較高,全年均值超過二類海水水質標準,屬于典型的富營養化河口。
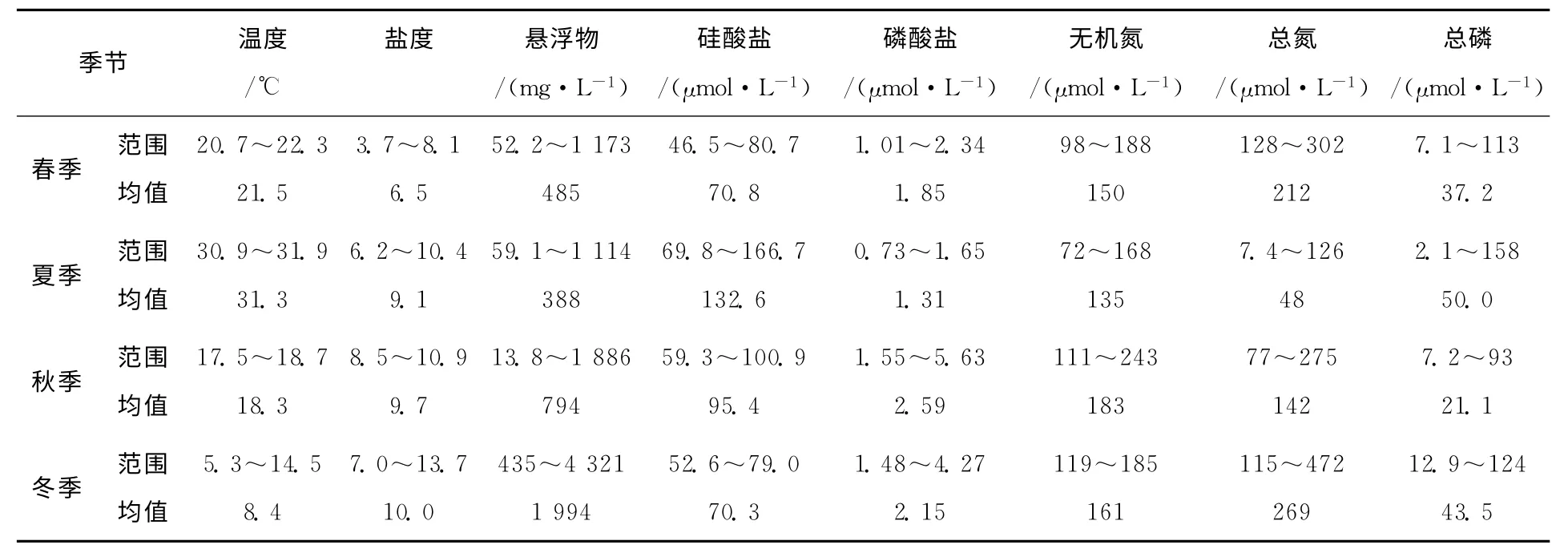
表1 錢塘江河口水化學因子統計
3.2 葉綠素a的分布特征
調查海區春季葉綠素a的含量范圍在0.08~2.03μg/L之間,平均值為0.48μg/L;夏季葉綠素a的含量范圍在0.01~2.99μg/L 之間,平均值為0.83μg/L;秋季葉綠素a 的含量范圍在0.15~2.29μg/L之間,平均值為0.77μg/L;冬季葉綠素a的含量范圍在0.08~1.23μg/L 之間,平均值為0.51μg/L;全年調查海域葉綠素a的含量變化范圍在0.01~2.99μg/L 之間,平均值為0.76μg/L。總體上全年調查期間調查海域的葉綠素a波動較大,夏季海水中葉綠素a含量較高,春季相對較低。
圖3 給出了錢塘江河口葉綠素a的分布情況,從圖3可以看出,春季表層海水葉綠素a在西側近岸海域較低,東側離岸海域較高;夏季表層海水葉綠素a分布整體上與春季類似,呈現近岸低、離岸高的趨勢;秋季表層海水葉綠素a整體上分布與春季和夏季相反,高值區更多地集中在西側近岸海域;冬季海水葉綠素a分布趨勢整體上相對較低,高值區位于核電南側鄰近海域。整體上表層葉綠素a相對較高,底層相對較低,各個季節表底層葉綠素a相差不大,垂直分布差異不明顯。調查海區夏季葉綠素a為全年最高,冬季和春季的葉綠素a相對較低;調查海域各季節葉綠素a含量相對其他海區較小,主要原因可能是該水域具有暖溫帶強潮河口區生態特征,盡管水體含豐富營養鹽和充足溶解氧,由于同時存在著諸如懸沙含量高、透明度小等不利于生物繁殖生長的環境因子,使得該海域生產力偏低。與鄰近的東海相比[11],該水域葉綠素a的含量普遍較低,大約為前者的1/4,與較為混濁的長江水體葉綠素a的含量接近[12]。
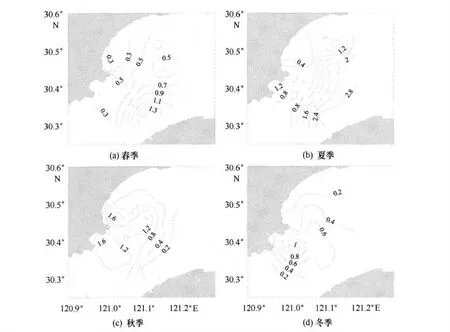
圖3 錢塘江河口葉綠素a(μg/L)的分布
3.3 浮游植物生物量及其影響因素
圖4給出了錢塘江河口浮游植物細胞總量分布情況,由圖4 可以看出,春季航次共采集鑒定浮游植物35種,其中硅藻31種,占出現總種類數的94.1%,優勢種(以優勢度Y≥0.02計)主要有中肋骨條藻、圓篩藻類(主要包括瓊氏圓篩藻、虹彩圓篩藻等)和菱形藻類(主要是洛氏菱形藻);表層浮游植物總細胞數量在70×105~2 475×105cell/m3,最大值出現在H06 號站,最小值出現在H04 號站,平均值為444×105cell/m3。夏季航次共采集鑒定浮游植物40 種,其中硅藻35種,硅藻類的貢獻占出現總種類數的87.5%;表層浮游植物總細胞數量在96×105~1 221×105cell/m3,最大值為H19 號站,最小值出現在H14號站,平均值為420×105cell/m3,細胞總量在杭州灣灣頂錢塘江口較低,其中中肋骨條藻在表層水采樣品中占絕對優勢,其次是直鏈藻類和圓篩藻類。秋季航次共鑒定浮游植物37 種,其中硅藻33種,硅藻類的貢獻最大,占出現總種類數的89.2%;表層浮游植物總細胞數量在108×105~1 834×105cell/m3,平均值為405×105cell/m3,最大值出現在13號站,最小值出現在10號站,調查海域中部細胞總數相對較高,中肋骨條藻在表層水采樣品中占絕對優勢,其次是圓篩藻類。冬季航次共鑒定浮游植物31種,其中硅藻32種,硅藻類占出現總種類數的96.8%;表層浮游植物總細胞數量在33×105~72×105cell/m3,平均值為54×105cell/m3,整個調查海域表層浮游植物總量處在一個較低的水平,總體上調查海域北側相對高于調查海域南側,水采樣品中優勢種主要有圓篩藻類、菱形藻和舟形藻(表2)。
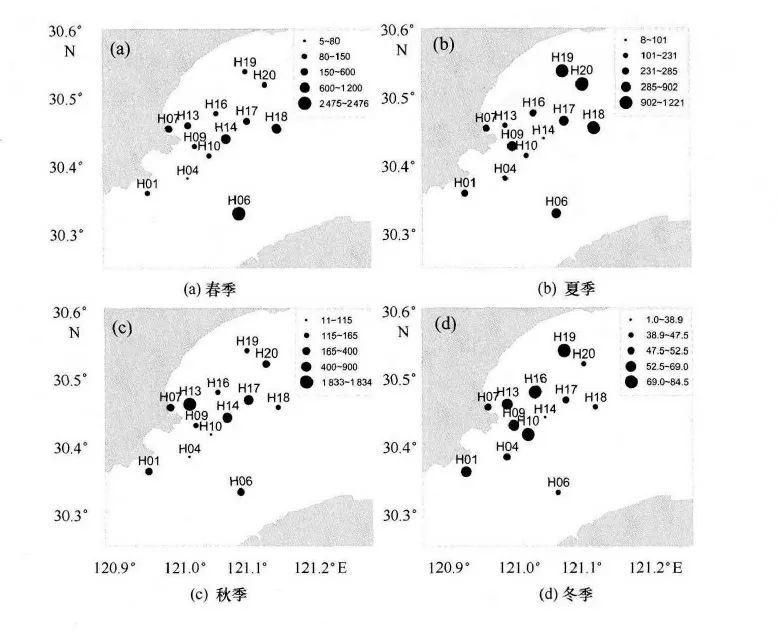
圖4 錢塘江河口浮游植物細胞總量(×105cell/m3)的分布
一般而言,高渾濁的水域浮游植物群落結構以硅藻為主,非硅藻浮游植物較少。與鄰近的東海相比[11,13],該水域浮游植物種類較為單一,且生物量普遍不高。調查海域浮游植物的物種多樣性指數在河口左岸水域和河口的中部水域較高,而在河口右岸和河口下游數值較低。淡水種和半咸水種是該海域優勢的浮游植物,淡咸水混合的結果造成了上述分布格局的出現。

表2 錢塘江河口浮游植物細胞總量和優勢種(優勢度)
典范對應分析(CCA)顯示,溫度、鹽度和懸浮顆粒物是影響優勢種中肋骨條藻生物量的重要水文因素,而DSi、N/P 與Si/N 比是影響其生物量的化學要素。懸浮顆粒物與優勢種生物量成負相關關系,這表明濁度對生物量的制約機制,亦表明該水域濁度是控制生物量的重要因素;盡管該水域營養鹽濃度普遍較高,但該水域并未有發生赤潮的記錄,顯然高的濁度減少了赤潮發生的幾率。溫度均與優勢種呈正相關關系,這表明高的溫度有利于優勢種的增殖。不過,鹽度與優勢種多呈負相關關系,這很大程度上是由于該水域物種多屬于淡水種和半咸水種,低的鹽度有利于其增殖過程。總氮與總磷多與優勢種呈負相關關系,這很大程度上是由于該水域水體懸浮物含量較高,顆粒氮磷又是該海域的主要營養鹽形態,其增加了水體的濁度,影響了優勢種的增殖。溶解硅多與優勢種呈正相關的關系,Si/N在春季和冬季是優勢種存在的限制性因子,這主要是由于這兩個季節溶解硅濃度較低的緣故,導致Si/N 比低于其他季節,成為限制生物生長的因素。值得說明的是,在冬季,圓篩藻超過中肋骨條藻,成為該海域的優勢種,這是因為在冬季懸浮顆粒物的含量最高,相比于中肋骨條藻而言,CCA 顯示圓篩藻更適宜生活在高渾濁的水體,加之冬季溫度較低,這也限制了中肋骨條藻的增殖。因此,該區域控制生物量與組成的主要因素是濁度/懸浮顆粒物含量,其次是溫度,高的營養鹽并未對該水域生物量格局造成大的影響。然而,與1989—1990年數據相比[6],該海域葉綠素a含量以及浮游植物細胞數明顯降低,硅藻種類數也有減少了30%~50%,另外綠藻、藍藻和甲藻的出現頻率也明顯增加。造成這方面變化的原因很可能與營養鹽結構變化有關,從圖5可以看出,Si/N 比或N/P 比往往是優勢種生物量的限制性因子,如春季的Si/N 比與夏季的N/P均對優勢種的數量產生限制。
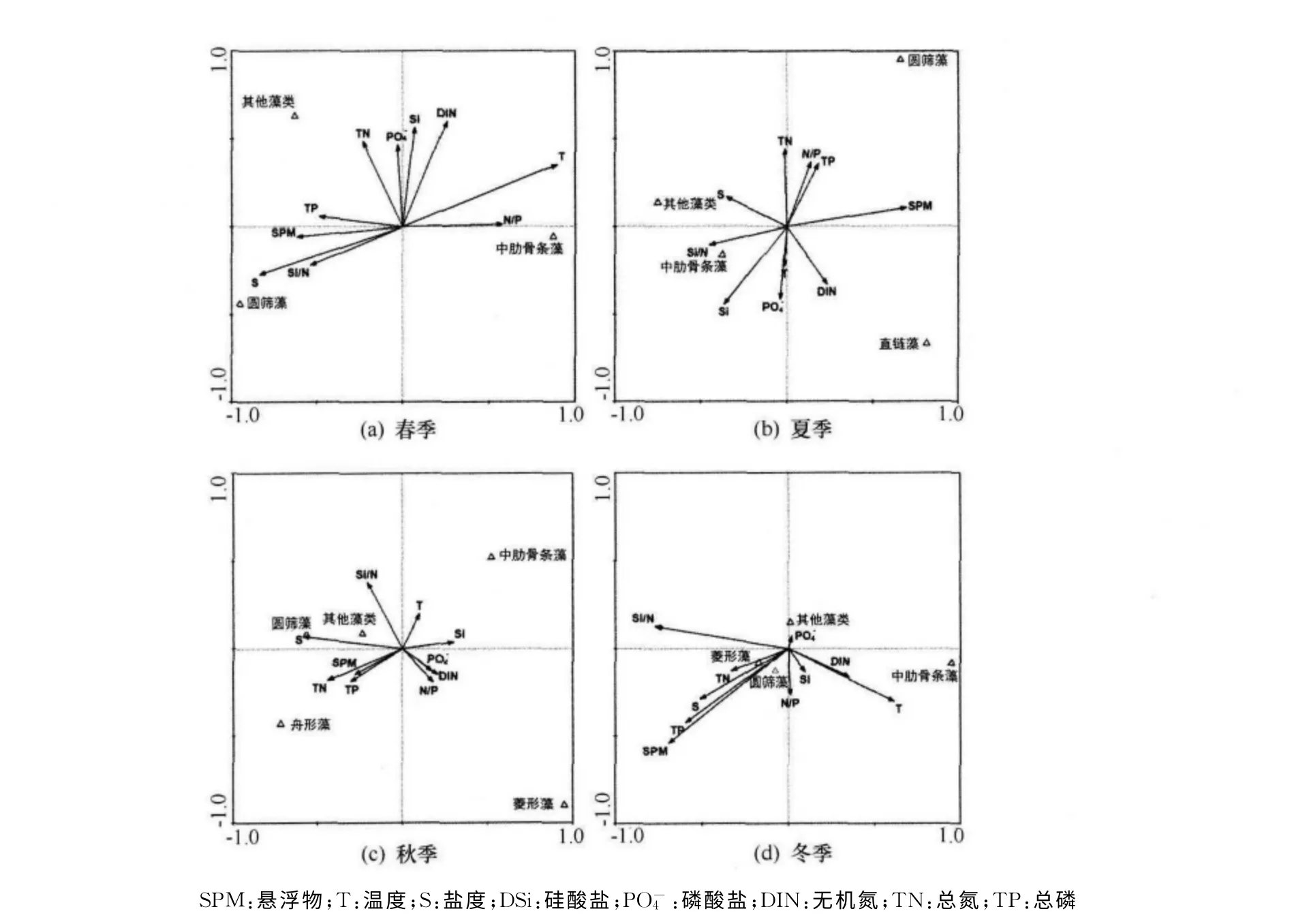
圖5 錢塘江河口環境因子與優勢種生物量的關系
由此可見,營養鹽豐富的水域,生產力并不一定高。由于同時存在著多種不利于浮游植物繁殖的環境因子的綜合影響,營養鹽不能被充分的利用而顯得過剩,反而會使得鄰近水體存在潛在的富營養化導致赤潮的趨勢,而影響物質的再循環。從目前數據來看,杭州灣潮流強,水體渾濁度高,生物量相對較低,因此其“河口過濾器”效應是有限的。但是,由于杭州灣近海的氮磷負荷超過了硅的輸入量,使得灣內營養鹽結構發生了較大的變化,很可能對鄰近的東海產生較大的影響,增加誘發赤潮的風險,應加強監測。
4 結論
(1)調查海域春、夏、秋、冬季葉綠素a含量分別為0.48μg/L、0.83μg/L、0.77μg/L 和0.51 μg/L。與鄰近的海域相比,該水域葉綠素a的含量普遍較低,主要是由于河口區水體懸沙含量高、透明度小等環境因子不利于浮游植物繁殖和生長所致。
(2)調查海域浮游植物的出現總量以春季最高,夏季和秋季次之,到冬季則顯著降低。浮游植物的種類主要為硅藻,占浮游植物總量的87.5%~96.8%,優勢種為中肋骨條藻和圓篩藻,浮游植物的物種多樣性指數在河口左岸水域和河口的中部較高。
(3)調查海域環境因子與優勢種生物量的CCA 結果表明,濁度是控制該海域浮游植物種類的限制因子,并在群落結構演替中發揮關鍵作用。與歷史數據相比,該海域葉綠素a含量以及浮游植物細胞數明顯降低,硅藻種類數也有減少了30%~50%,另外綠藻、藍藻和甲藻的出現頻率也明顯增加,營養鹽結構變化可能是浮游植物數量和種類變化的重要原因。
[1]TURNER R E,RABALAIS N N.Coastal eutrophication near the Mississippi river delta[J].Nature,1994,368(6472):619-621.
[2]CHAI C,YU Z,SONG X,et al.The status and characteristics of eutrophication in the Yangtze River(Changjiang)Estuary and the adjacent East China Sea,China[J].Hydrobiologia,2006,563(1):313-328.
[3]CUSHING D H.Plankton production and year-class strength in fish populations:an update of the match/mismatch hypothesis[J].Advances in Marine Biology,1990,26:249-293.
[4]LEPIST?L,HOLOPAINEN A L,VUORISTO H.Type-specific and indicator taxa of phytoplankton as a quality criterion for assessing the ecological status of Finnish boreal lakes[J].Limnologica-Ecology and Management of Inland Waters,2004,34(3):236-248.
[5]GAO Shengquan,YU Guohui,WANG Yuhen.Distributional features and fluxes of dissolved nitrogen,phosphorus and silicon in the Hangzhou Bay[J].Marine Chemistry,1993,43:65-81.
[6]國家海洋局第二海洋研究所.秦山核電站鄰近水域零點生態調查報告[R].1990.
[7]張健,施青松,鄔翱宇,等.杭州灣豐水期主要污染因子的分布變化及成因[J].東海海洋,2002,20(4):35-41.
[8]秦銘俐,蔡燕紅,王曉波,等.杭州灣水體富營養化評價及分析[J].海洋環境科學,2009,28(A01):53-56.
[9]高生泉,陳建芳,金海燕,等.杭州灣及鄰近水域營養鹽的時空分布與富營養化特征[J].海洋學研究,2011,29(3):36-47.
[10]LIU S M,ZHANG J,CHEN H T,et al.Nutrients in the Changjiang and its tributaries[J].Biogeochemistry,2003,62(1):1-18.
[11]王作華.黃東海葉綠素a和溶解有機碳分布特征及固碳強度初探[D].青島:中國海洋大學,2008:1-89.
[12]RAN X B,YU Z G,CHEN H T,et al.Silicon transport of Changjiang River:could the three gorges reservoir be a filter?[J].Environ Earth Sci.,2013,70(5):1-13.
[13]夏平,陸斗定,朱德弟,等.浙江近岸海域赤潮發生的趨勢與特點[J].海洋學研究,2007,25(2):47-56.
