高粘附性戊糖片球菌的篩選、標記及其表面疏水與自凝聚性特征
2015-11-11 02:38:04楊振泉靳彩娟張咪王曉霖高璐顧瑞霞
食品與生物技術學報 2015年9期
關鍵詞:能力
楊振泉,靳彩娟,張咪,王曉霖,高璐,顧瑞霞
(1.揚州大學食品科學與工程學院,江蘇揚州225127;2.江蘇省乳品生物技術與安全控制重點實驗室,江蘇揚州225127)
高粘附性戊糖片球菌的篩選、標記及其表面疏水與自凝聚性特征
楊振泉1,2,靳彩娟1,張咪1,王曉霖1,高璐1,顧瑞霞1,2
(1.揚州大學食品科學與工程學院,江蘇揚州225127;2.江蘇省乳品生物技術與安全控制重點實驗室,江蘇揚州225127)
通過常規乳酸菌分離技術結合形態觀察、生理生化試驗,以及16S rDNA同源性分析等方法,從臭豆腐發酵鹵水中分離獲得7株戊糖片球菌分離株,并通過隨機擴增多態性DNA技術構建了不同菌株的特征指紋圖譜;以Caco-2細胞和固定化腸黏液蛋白質作為體外模型,研究了菌株的粘附能力,并探討了菌株粘附能力與基因型以及表面疏水性、自凝聚能力等表型特征的相關性。結果顯示,發酵鹵水中的戊糖片球菌存在高度分子多樣性,7個分離株中存在6種不同的指紋圖譜模式,其中A5型菌株(F28-8和Y27-4)對Caco-2細胞和腸黏液蛋白質的粘附性最強,并顯示了高度的疏水性(>90%)和較強的自凝聚能力(>25%)。相關性分析表明,戊糖片球菌表面疏水率和自凝聚率與Caco-2細胞粘附率測定結果呈顯著正相關(r=0.900和0.792,P<0.05),但是與腸黏液蛋白質粘附率測定結果相關性不顯著(r=0.426和0.700,P>0.05)。研究成果為建立高粘附性戊糖片球菌快速篩選方法及其體內定植和分布研究提供了依據。
戊糖片球菌,粘附性,表面疏水性,自凝聚能力,DNA指紋圖譜
戊糖片球菌(Pediococcus pentosaceus)廣泛分布于腌制和泡制蔬菜、干酪、香腸等傳統發酵食品中,對提高發酵食品的營養、風味及安全性具有重要作用[1-2]。該菌代謝糖類產生乳酸,并產生IIa類片球菌素,對病原菌及腐敗微生物產生強烈的抑制效應[3]。已有的研究表明,戊糖片球菌的一些菌株具有提高機體的天然免疫能力、促進健康以及抵抗病原菌的侵襲等益生特性[4-5],具有用于人類或動物的微生態制劑的開發潛力。菌株粘附能力是微生態制劑在腸道發揮生態效應和生理作用的重要基礎,篩選高粘附性菌株對微生態制劑開發具有重要意義,但目前國內外對戊糖片球菌粘附特性及其機制的研究尚不多見。利用人結腸癌細胞系Caco-2評價乳酸菌的粘附能力是目前廣泛使用的體外模型和方法[6],但是沒有考慮到腸道內覆蓋在腸上皮細胞外層的腸黏液的作用[7],固定化小腸黏液蛋白質模型能夠反映乳酸菌在體內與腸黏液的作用[8],結合這兩種模型可以較好地評價乳酸菌的粘附特性,但是這兩種模型需要復雜的細胞培養和實驗動物操作,價格昂貴,不利于大規模的菌株篩選。
細菌表面疏水性和自凝聚性與粘附特性密切相關。表面疏水性是決定細菌非特異性粘附到生物表面的重要動力,在乳酸菌向腸壁粘附和定植的過程中發揮重要的作用[9]。自凝聚特性有助于細菌形成生物膜并在宿主腸道定植,阻止病原菌的粘附和侵襲,并且與致病菌的清除存在一定關聯[10]。目前已有研究表明,在部分雙歧桿菌和乳酸桿菌菌種中,基于菌株的表面疏水性和自凝聚能力與其對腸上皮細胞粘附能力的相關性,可以應用簡單廉價的凝聚性和疏水性測定方法替代昂貴的細胞和動物評價模型對菌株的粘附能力進行預測和初步篩選[11],對從大量菌株中篩選高粘附性菌株,縮小試驗范圍和降低成本具有重要意義。但是,目前對于戊糖片球菌的粘附能力和表面疏水性以及自凝聚能力之間的相關性還需要進一步數據證實。本研究中從臭豆腐發酵鹵水中分離鑒定了7株戊糖片球菌,通過Caco-2細胞和固定化小腸黏液蛋白質模型,比較了不同菌株的粘附能力,并探討了粘附能力與菌株基因型、表面疏水性以及自凝聚特性的相關性,為高粘附性戊糖片球菌菌株高效定向篩選及微生態制劑開發提供了候選菌株和依據。
1 材料與方法
1.1材料
1.1.1主要試劑PCR試劑、溶菌酶、蛋白酶K,均購自上海生工生物工程有限公司;MRS培養基以及細菌生化鑒定試劑,廣州環凱生物試劑有限公司產品;細菌16SrDNA擴增引物8F:5’-AGAGTTTGATCCTGGCTCAG-3’和15R:5’-AAGGAG GTGATCCAGCCGCA-3’,以及DNA指紋圖譜擴增隨機引物M13:5'-GAGGGTGGCGGTTCT-3'和M14:5'-GAG GGTGGGGCCGTT-3',均由上海生工生物工程有限公司合成;其它化學試劑均為國產分析純。
1.1.2主要儀器設備ABI2720型熱循環儀,美國Life Technologies公司制造;凝膠成像系統,法國Vilber Lourmat公司制造;UV-7504C分光光度計,上海欣茂儀器有限公司制造;Elx-800型酶標儀,美國Bio-Tek公司制造;顯微鏡,日本Olympus公司制造;S-4800型場發射掃描電鏡,日本日立公司制造;Heracell 240i CO2培養箱,美國Thermo Scientific公司制造。
1.1.3細胞與實驗動物Caco-2細胞株,購自中國科學院上海生命科學研究院細胞資源中心;DMEM培養液、胎牛血清、質量分數0.25%胰蛋白酶、DHank′s緩沖液,均購自GIBCO公司;青霉素和鏈霉素,美國Sigma公司產品;6周齡ICR實驗小鼠,購自揚州大學比較醫學中心。
1.2方法
1.2.1戊糖片球菌的分離及生理生化鑒定臭豆腐發酵鹵水樣品分別采集自揚州本地不同臭豆腐生產作坊。發酵鹵水樣品充分混勻后用滅菌生理鹽水10倍稀釋。吸取250 μL不同稀釋度樣品液涂布含有質量分數0.75%CaCO3的MRS平板,放入厭氧罐,37℃恒溫培養48 h,選擇分離程度較好的平板挑取有溶鈣圈的單菌落,在MRS平板上反復劃線純化,按文獻[12]所述的方法對接觸酶陰性、產酸的革蘭氏陽性球菌進行耐鹽性、溫度、耐酸堿、動力學以及糖和氨基酸發酵試驗,細菌的細胞微觀形態采用掃描電鏡觀察。
1.2.2細菌基因組DNA提取基因組DNA的制備按文獻[13]所述的CTAB法進行,并作適當改良。吸取48 h培養物1.5 mL置于離心管中,10 000 r/ min離心5 min,收集細胞;細胞用TE緩沖液洗滌2次后重懸于567 μL TE緩沖液中,加入30 μL質量分數10%SDS和5 μL 20 mg/mL蛋白酶K,顛倒混勻,37℃溫育1 h;加入100 μL 5 mol/L的NaCl溶液,混勻后加入80 μL CTAB-NaCl溶液,65℃水浴10 min。用酚-氯仿-異戊醇(體積比25∶24∶1)抽提兩次,上清液加2倍體積的無水乙醇和0.1倍體積的3 mol/L NaAc,-20℃靜置1 h;10 000 r/min離心5 min收集沉淀,用體積分數70%乙醇洗滌兩次,沉淀經真空冷凍干燥2 h后用40 μL TE緩沖液溶解,加入1 μL 10 mg/L的RNA酶,37℃消化1 h。提取物用核酸測定儀測定總DNA濃度,-20℃保存備用。
1.2.316S rDNA PCR擴增與測序鑒定以細菌基因組DNA為模板進行16S rDNA的PCR擴增。PCR反應體系(50 μL)包括:2 μL模板(50 ng/μL)、2 μL dNTPs(10 mmol/L)、引物8F(10 pmol/μL)和15R(10 pmol/μL)各2.0 μL,以及5 μL 10×Buffer、5 μL MgCl2(25 mmol/L)、0.3 μL Tag酶(5 U/μL),最后加ddH2O補足至50 μL。PCR擴增程序為:94℃預變性5 min;94℃變性40 s,50℃退火35 s,72℃延伸2.0 min,35次循環;72℃延伸10 min。取7.0 μL PCR產物用質量分數1.0%的瓊脂糖凝膠電泳檢測片段大小,其余PCR產物委托上海生物工程有限公司測序,所得序列在GenBank(http://www.ncbi. nlm.nih.gov)數據庫中進行在線比對,序列同源性大于99%設為相同種。
1.2.4菌株分子指紋圖譜鑒別菌株之間的基因型差異通過RAPD指紋圖譜鑒別。以細菌基因組DNA為模板,應用M13和M14隨機引物進行RAPD擴增。將DNA提取物用ddH2O稀釋至100 ng/μL,隨機引物濃度稀釋至10 pmol/μL;優化的PCR擴增反應體系為:模板2.0 μL,dNTPs 0.5 μL,隨機引物2.0 μL,10×Buffer 2.5 μL,MgCl22.5 μL,Tag酶0.3 μL,ddH2O 15.7 μL;RAPD反應循環參數為:95℃預變性2 min,94℃變性1 min;53℃退火1 min;72℃延伸1 min,30個循環;72℃補充延伸8 min。取10 μL產物在質量分數1.5%的瓊脂糖凝膠上進行電泳(電壓5 V/cm)1.5 h,EB染色20 min,凝膠成像系統拍照記錄,根據條帶清晰程度、彌散背景以及帶型強弱情況選擇穩定的指紋圖譜進行分析。
1.2.5指紋圖譜聚類分析根據在指紋圖譜中同一遷移率位點上的擴增條帶的“有”和“無”進行數字化處理,用“1”表示“有”,“0”表示“無”,建立矩陣,應用DPS7.05數據處理系統中的0-1系統聚類分析程序,選擇Nei氏遺傳距離和類平均法進行聚類分析。
1.2.6菌株表面疏水性測定參照文獻[8]應用微生物粘著碳氫化合物(Microbial Adhesion To Hydrocarbons,MATH)法測定菌株的表面疏水性。取36 h細菌培養物4 000 r/min離心10 min收集菌體,生理鹽水洗滌兩次,5 000 r/min離心10 min。以生理鹽水調整受試菌株菌液濃度,使菌懸液在600 nm波長下A值約為1.0(A600=1.0)。取2 mL菌懸液與2 mL二甲苯混合,渦旋120 s,室溫靜置30 min分層。取1 mL水相,以生理鹽水為空白對照組,測定A600,每株細菌平行做3管重復。細菌表面疏水率(CSH)計算:
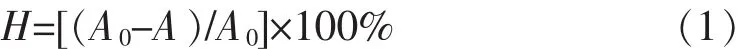
式(1)中:H即CSH,A0和A分別是與二甲苯混勻前后水相測定所得的A600值。
1.2.7菌株自凝聚能力測定菌株自凝聚能力測定參照文獻[14]進行。菌株懸液制備同1.2.6。取4 mL制備好的菌懸液(A600=1.0)置于10 mL帶刻度的螺口試管中,室溫靜置分層,分別在靜置1、2、3、4、5 h后吸取1 mL上層懸液測A600值,分別記作A1、A2、A3、A4、A5,每個菌株每個時間點平行做3管重復。細菌自凝聚率(AA)計算:
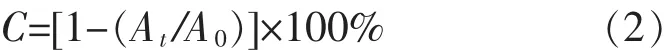
式(2)中:C即AA,A0和At分別是自凝聚前后上層懸液在600 nm下測量所得的A值。
1.2.8菌株對Caco-2細胞的粘附能力測定采有Caco-2細胞作為體外模型測試菌株的粘附能力。Caco-2細胞用添加體積分數10%胎牛血清和100 U/mL青鏈雙抗的DMEM培養液培養,細胞于體積分數5%CO2培養箱中37℃培養,2 d換液一次。細胞用質量分數0.25%胰酶消化并轉移到24孔細胞培養板中,于37℃、體積分數5%CO2培養箱中培養至單層細胞形成。戊糖片球菌用MRS活化兩代,離心收集菌體,PBS洗2次,用不含胎牛血清和抗生素的DMEM培養液重懸菌體。傾去Caco-2細胞培養板中的培養液,用無菌PBS沖洗2次,然后加入0.5 mL菌懸液,培養1 h后用無菌PBS洗滌4次,洗去未粘附的菌體,加入1 mL Triton 100(體積分數1%),使粘附的菌體同細胞分離,吹打混勻,梯度稀釋平板法測定粘附細菌數。用細菌數與陰性對照孔中的細胞數的比值(cfu/cell)表示菌株對Caco-2細胞的粘附能力。
1.2.9菌株對小腸黏液的粘附能力測定參照文獻[15]方法制備小腸黏液,將6周齡ICR小鼠固定24 h后解剖,取小腸置于冰面,用無菌PBS(pH 7.2,0.01 mol/L)沖洗小腸3次后縱向剪開,刮取小腸內表面黏液,加入2倍體積的PBS緩沖液,混勻,然后12 000 r/min、4℃離心10 min,收集上清液,用考馬斯亮藍法測定黏液蛋白質質量濃度,分裝后-20℃保存備用。取1.0 mL不同濃度的戊糖片球菌懸液于離心管中,10 000 r/min離心10 min,棄上清液,加入1.0 mL質量分數0.1%結晶紫溶液染色50 min后離心收集菌體,PBS洗滌5次后加入1.0 mL檸檬酸鹽緩沖液重懸,室溫放置60 min后測定A540,建立A540和菌懸液細胞數的標準曲線。再參考Vesterlund等建立的方法[16]測定菌株對小腸黏液的粘附能力。取96孔板,每孔加入100 μL鼠小腸黏膜提取液(蛋白質2.0 mg/mL),4℃過夜固定。每孔加入100 μL菌懸液,4℃孵育2 h傾去懸液并用PBS緩沖液洗滌4次,除去未粘附的細菌。將96孔板置于60℃烘箱30 min。每孔加入100 μL質量分數0.1%結晶紫溶液染色50 min,250 μL PBS洗滌5次,加入100 μL檸檬酸鹽緩沖液(pH 4.3,0.02 mol/L),室溫放置60 min后測定A540,并根據建立的標準曲線計算多孔板中粘附的細菌數量,以粘附細菌數和加入細菌數的百分數表示菌株的粘附能力。
1.2.10數據分析菌株表面疏水率、自凝聚率以及粘附指數試驗結果取3次試驗的平均值,以平均值±標準差表示,菌株間差異及數據間的相關性采用Sigmaplot 10.0軟件中的t檢驗和線性回歸模式進行統計分析,以P<0.05為有統計學意義。
2 結果與分析
2.1戊糖片球菌分離鑒定結果
通過改良MRS平板分離和厭氧培養在采集的不同來源的臭豆腐發酵鹵水樣品中均分離得到大量革蘭氏陽性、接觸酶陰性的產酸菌株,其中菌株F28-8、Y45-30、F3、Y27-4、H13、H6和H10的細胞形態呈二聯或四聯球狀的分離株,如圖1(a)所示,掃描電鏡結果顯示分離株的細胞形態以四聯球為主,并且成片聚集在一起,如圖1(b)所示。這7個菌株具有相同的生理生化特征,實驗結果如表1所示,根據伯杰細菌手冊[17]及常見細菌系統鑒定手冊[12]描述,這7株球菌菌株均具有典型的戊糖片球菌的生理生化特征。進一步提取7個球菌分離株的基因組DNA并進行16S rDNA擴增,擴增產物和預期大小一致(1 500 bp左右)。將擴增產物測序結果在基因庫GenBank(http://www.ncbi.nlm.nih.gov)中進行Blasten比對,結果如表2所示,序列同源性相似性最高的均為戊糖片球菌(最大同源性>99%),只是不同的菌株在GenBank中相似度最高的菌株有所不同。因此,這7個分離株均屬于戊糖片球菌種,但可能屬不同的基因型。
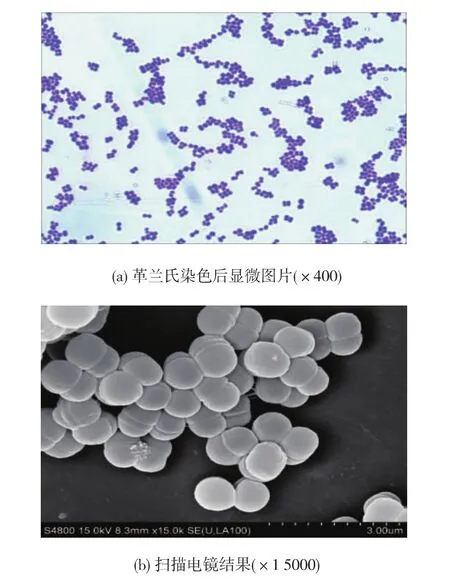
圖1 戊糖片球菌分離株的細胞形態Fig.1 Cell morphology of Pediococcus pentosaceus strains
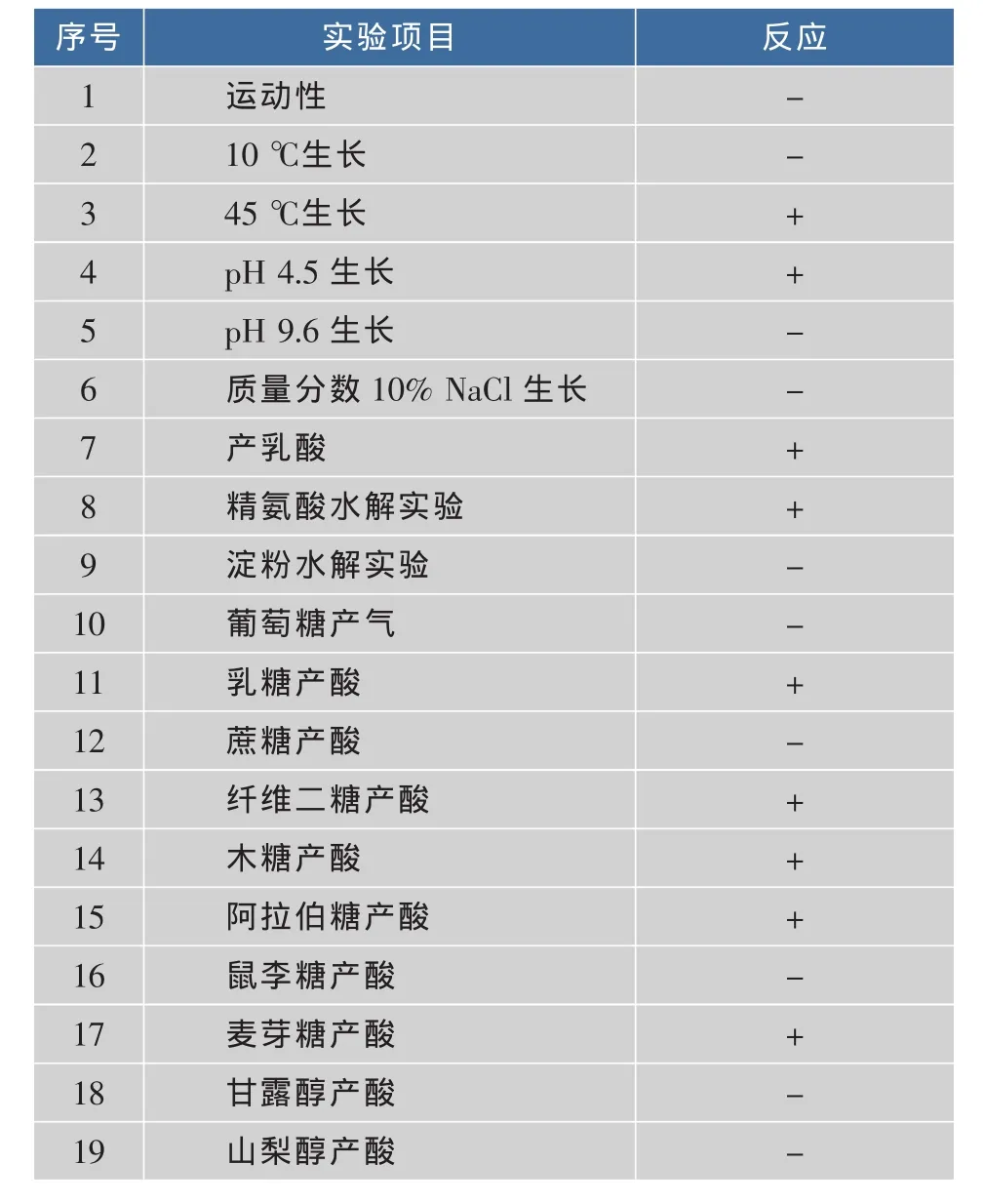
表1 戊糖片球菌分離株的生理生化鑒定結果Table 1 Resultsofphysiologicalandbiochemical characteristicsofPediococcuspentosaceus strains
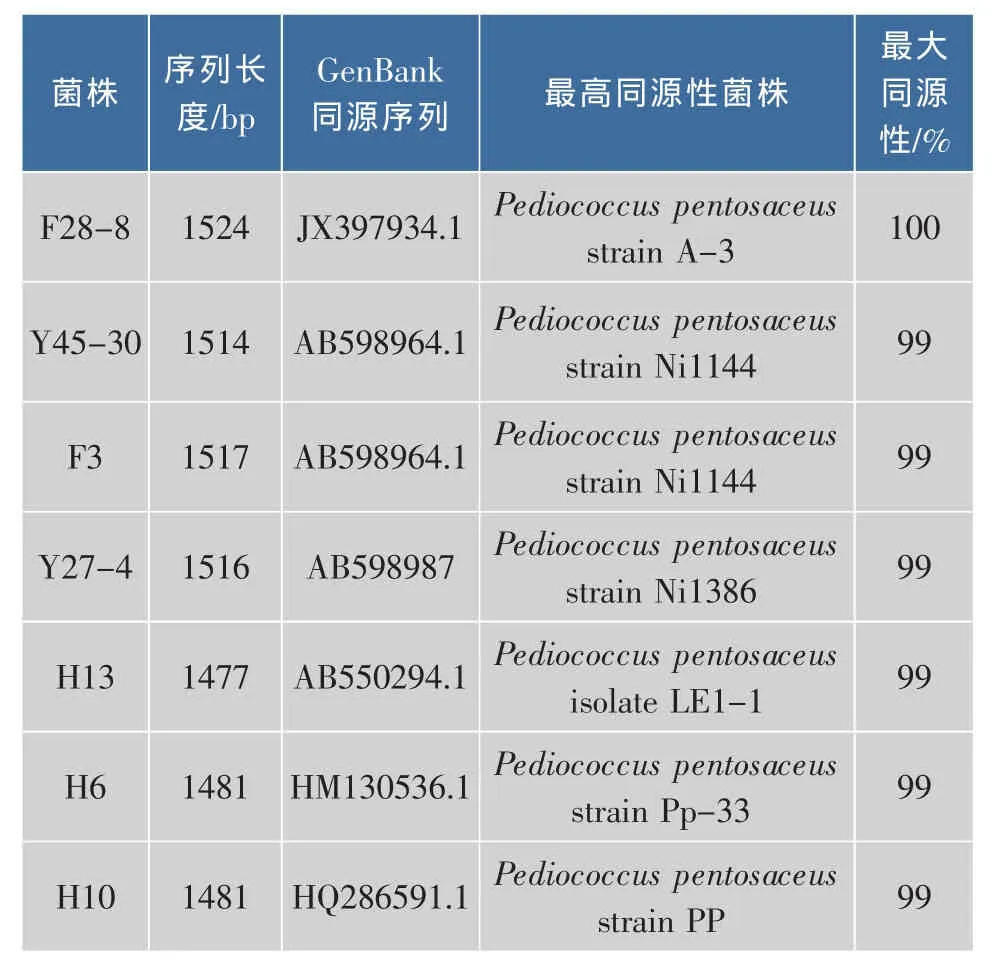
表2 戊糖片球菌分離株16S rDNA測序鑒定結果Table 2 Sequencing results of 16S rDNA identification of Pediococcus pentosaceus strains
2.2戊糖片球菌的菌株分子多樣性
應用RAPD分子分型方法對7株戊糖片球菌分離株的基因型進行分析,兩種不同隨機引物M13和M14的擴增譜帶模式如圖2所示。M13擴增得到2個不同的指紋圖譜(A型和B型),M14擴增得到5個不同的指紋圖譜,分別為1型、2型、3型、4型和5型。將兩種譜帶模式進行組合分型,獲得了6種基因型,表明發酵鹵水中的戊糖片球菌存在高度的分子多樣性,其中H6為A1型,H10為B2型,H13為B3型,F3為A4型,Y45-30為A6型,菌株F28-8和Y27-4譜型相同均為A5型。將菌株的特征指紋圖譜中的擴增條帶按“有”和“無”進行數字化處理,應用Nei氏遺傳距離和類平均法進行聚類分析,結果顯示在Nei氏遺傳距離為0.30處,可以分成4簇,其中第1簇包含菌株H6、F28-8、Y27-4和Y45-30,顯示了這4個菌株具有較為相似的基因型,而F3、H10和H13各自形成一簇,表明菌株間基因型具有較大的差異性。
2.3不同戊糖片球菌菌株細胞表面特性及粘附能力測定結果
7株不同戊糖片球菌細胞表面特性及粘附能力測定結果如圖3所示。MATH法測定表面疏水率結果顯示7株戊糖片球菌的表面疏水性存在高度的菌株差異性,如圖3(a)所示,不同菌株的表面疏水率大小介于12.7%~97.1%,其中3個基因型高度相似的菌株F28-8、Y27-4和Y45-30顯示了高度疏水性,疏水率分別為96.5%,97.1%和93.6%。而菌株F3和H13的疏水率為12.7%和24.2%,顯著低于其它菌株(P﹤0.01),為弱疏水性菌株。應用分光光度法對7株戊糖片球菌在渦旋后靜置1~5 h的自凝聚性進行測定,結果如圖3(b)所示。結果顯示,菌株懸液渦旋振蕩以后隨著靜置時間的延長自凝聚率逐漸增大,菌株間的自凝聚率具有顯著差異,其中菌株F28-8、Y27-7在2 h后的各個時間點的凝聚率均顯著高于其它菌株(P<0.01),顯示了較高的自凝聚能力。而菌株F3和H13顯示了較弱的自凝聚能力。應用Caco-2細胞作為模型研究了7株戊糖片球菌對腸上皮細胞的粘附性,結果如圖3(c)所示。結果表明,不同菌株的Caco-2細胞粘附率在12.8~57.4 cfu/cell之間,其中高疏水性菌株F28-8、Y27-4和Y45-30具有較高的粘附性,細胞粘附率分別為57.4、46.5 cfu/cell和51.7 cfu/cell,而弱疏水性菌株F3和H13細胞粘附率分別為12.8、10.6 cfu/ cell,顯示了相對較低的細胞粘附性。通過提取小鼠的腸黏液蛋白質建立體外模型測定了7株戊糖片球菌對腸黏液的粘附率,結果如圖3(d)所示。不同菌株對腸黏液蛋白質顯示了不同的粘附率,其中菌株F28-8和Y45-30顯示了較高的粘附性,分別為35.2%和25.8%,顯著高于其它菌株(P<0.01),菌株H13、H6和H10的粘附率分別為9.7%、5.7%和4.3%,顯示了相對較低的粘附性。
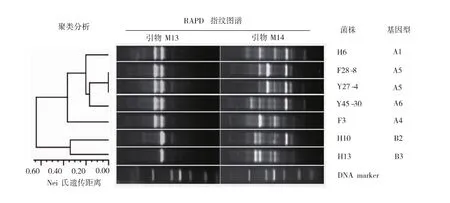
圖2 不同戊糖片球菌分離株的RAPD指紋圖譜及聚類分析Fig.2 RAPD fingerprinting and clustering analysis of different Pediococcus pentosaceus strains
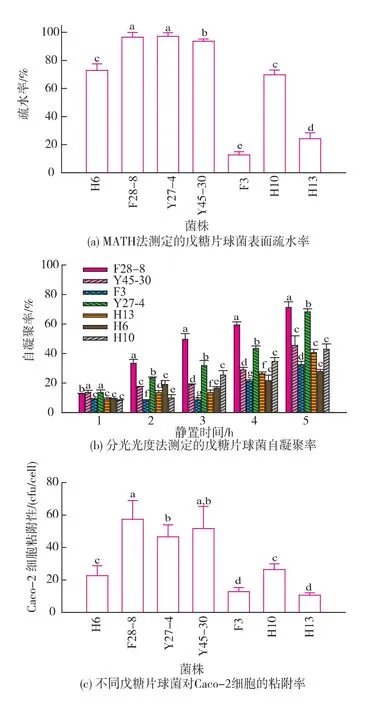
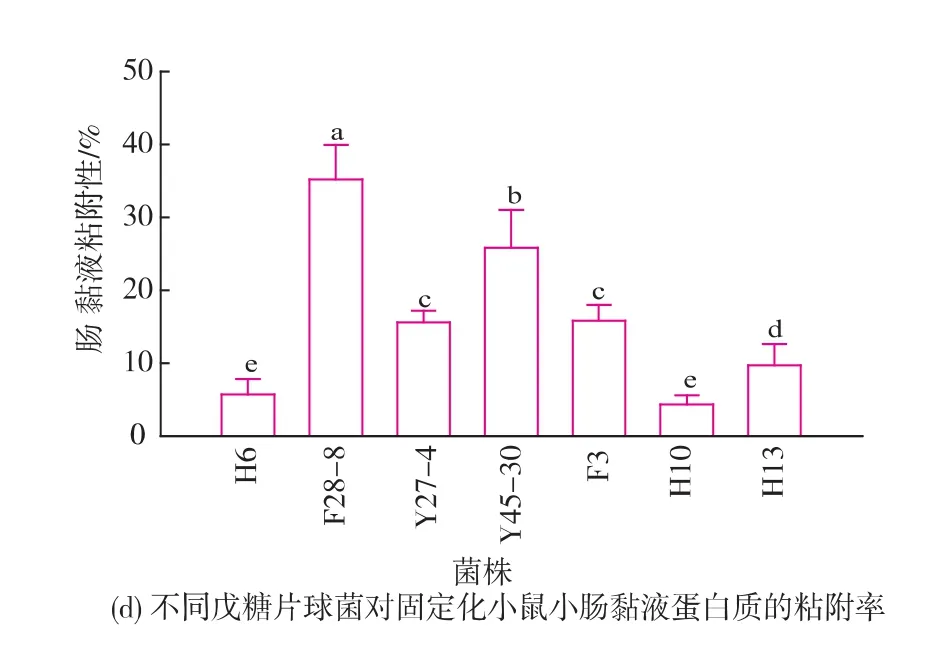
圖3 不同戊糖片球菌表面疏水性、自凝聚能力以及對Caco-2細胞和腸黏膜蛋白質的粘附能力比較Fig.3 Comparison of surface hydrophobicity,autoaggregation ability,and adhesion ability to Caco-2 cell andintestinalmucusofdifferentPediococcus pentosaceus strains
2.4相關性分析結果
將不同戊糖片球菌菌株的Caco-2細胞粘附率、黏液蛋白質粘附率、表面疏水率和自凝聚率(2 h)數據兩兩組合進行統計分析,繪制相關性散點圖(如圖4(a)—(f)所示),并用線性模型模擬它們之間的關系。結果表明戊糖片球菌不同菌株對Caco-2細胞粘附性和腸黏液蛋白質的粘附率呈顯著正相關(r=0.763,P=0.046,圖4(f)),可能Caco-2細胞和腸黏液蛋白質中存在共同的戊糖片球菌粘附素受體。菌株的表面疏水率和自凝聚率(2 h)與Caco-2細胞粘附率呈顯著正相關(r=0.900和0.792,P<0.05,圖4(a)(d)),但是與腸黏液蛋白質粘附率相關性不顯著(r=0.426和0.700,P>0.05,圖4(b)(e)),可能原因是戊糖片球菌在與Caco-2細胞粘附中表面疏水性和自凝聚能力等非特異粘附發揮了主要作用,而在腸黏液蛋白質作用體系中“粘附素-受體”特異性粘附起主要作用,表面疏水性和自凝聚能力影響較小。疏水性與自凝聚的相關性分析結果顯示,兩者呈正相關趨勢(r=0.727),但不顯著(P= 0.064,圖4(c)),疏水性是細菌發生自凝聚的主要驅動力,但是受多種因素的影響,如細菌表面組分和結構等,但是總體看來疏水性較高的菌株如F28-8和Y27-4自凝聚率也相對較高。
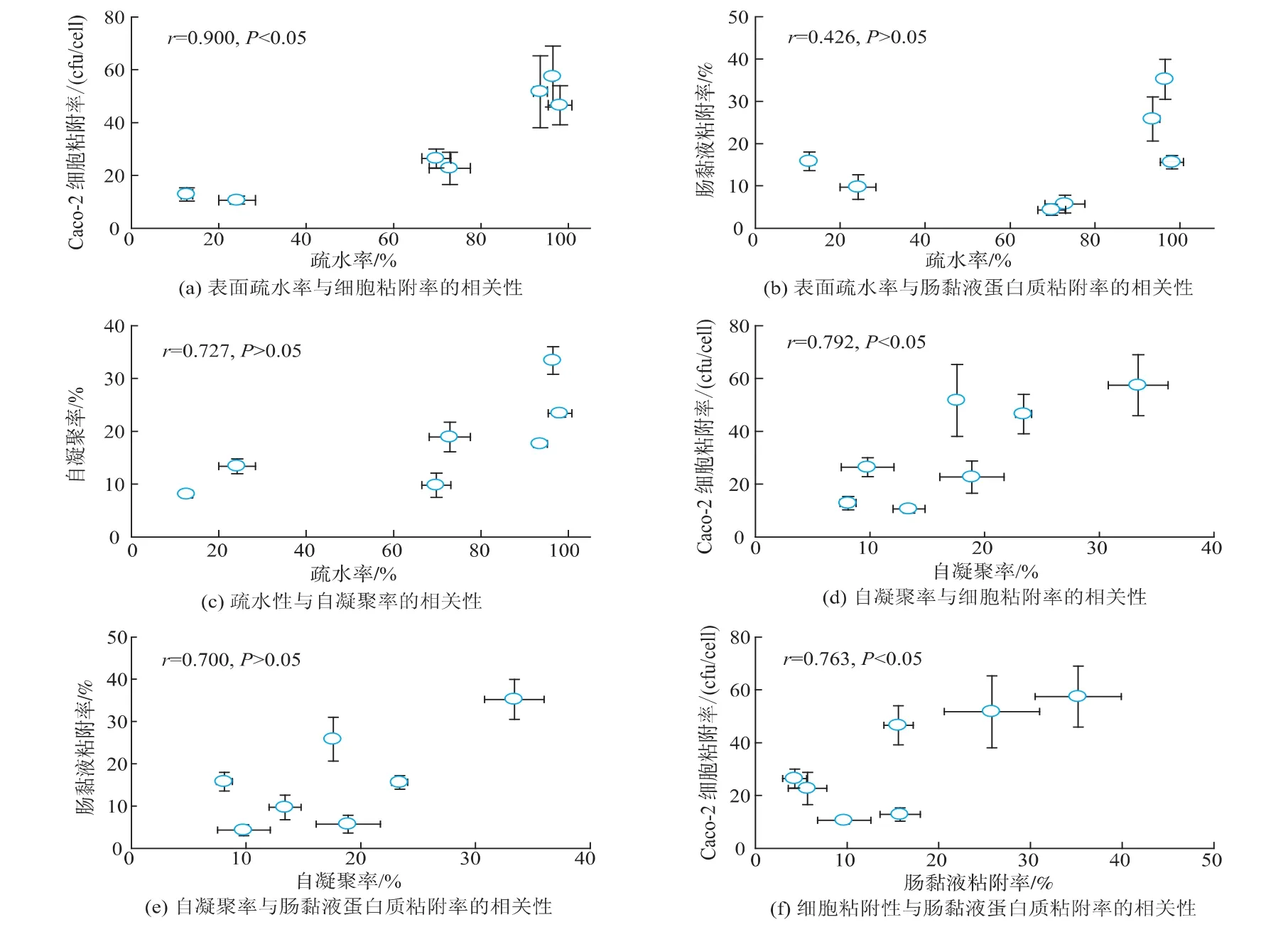
圖4 戊糖片球菌細胞表面特性與粘附能力的相關性分析Fig.4 Correlation analysis between cell surface characteristics and adhesion abilities of Pediococcus pentosaceus strains
3 討論
由于抗生素在人類疾病治療和動物飼養中的大量使用,導致環境中抗生素的積累和耐藥性菌株的不斷出現,對環境和食品安全帶來了威脅。微生態制劑因其能夠改善胃腸道功能、提高動物免疫力,在疾病預防和動物飼養中得到了大量推廣和應用。戊糖片球菌由于其獨特的抗菌性能和免疫調節功能而具有良好應用前景[4-5,18-19]。目前普遍認為粘附能力是益生菌發揮生理作用的前提,也是決定這種益生菌制劑實際效果的重要因素[20]。本課題研究中從臭豆腐發酵鹵水中分離鑒定了7株戊糖片球菌,DNA指紋圖譜顯示具有高度的種內分子多樣性。應用Caco-2細胞系和固定化腸黏液蛋白質兩種體外模型評價了7株戊糖片球菌分離株的粘附性,結果顯示特征指紋圖譜為A5型的兩個菌株(F28-8和Y27-4)顯示了最高的Caco-2細胞和腸黏液蛋白質粘附能力。
應用Caco-2細胞系和固定化腸黏液蛋白質模型評價菌株的黏附能力涉及到細胞培養和動物解剖試驗,操作復雜且耗費昂貴。應用分光光度法測定細菌自凝聚能力和微生物粘著碳氫化合物(Microbial Adhesion To Hydrocarbons,MATH)法測定疏水性,對細菌粘附性進行預測和初步篩選更為簡單、廉價。本文中探討了戊糖片球菌表面理化特性與其腸道細胞和黏液粘附的相關性,研究結果顯示,戊糖片球菌的表面疏水性和自聚集能力存在顯著菌株差異性,其中指紋圖譜最相近的3個菌株F28-8(A5型)、Y27-4(A5型)和Y45-30(A6型)顯示了高度疏水性(>90%)。本研究中戊糖片球菌的表面疏水率和自凝聚率(2 h)與Caco-2細胞粘附率呈顯著正相關(P<0.05),與前期報道中對Bifidobacterium longum菌株的研究結果[21]一致,表明菌株的表面疏水性和自凝聚性在Caco-2細胞粘附過程中發揮了重要作用。盡管有研究表明乳酸菌的一些菌株在體外對Caco-2細胞粘附性和腸黏液粘附能力并不一致[22],但本研究中戊糖片球菌對Caco-2細胞和固定化腸黏液的粘附率卻呈顯著正相關(P<0.05),這可能是戊糖片球菌對這兩種基質的粘附存在共同的作用機制和影響因素。但是表面疏水率和自凝聚率與腸黏液蛋白質粘附率相關性不顯著(P>0.05),可能原因是戊糖片球菌在固定化腸黏液蛋白質作用體系中“粘附素-受體”特異性粘附起了主要作用[23],表面疏水性和自凝聚能力引起的非特異粘附影響較小。
4 結語
本研究結果為建立體外篩選高粘附性戊糖片球菌菌株提供了實驗依據,并為進一步體內研究戊糖片球菌功能特性提供了候選菌株。也為開發戊糖片球菌生態制劑提供了候選菌株,同時為深入研究戊糖片球菌在動物體內的定植和動態變化提供了分子追蹤方法。對從大量菌株中篩選高粘附性菌株,縮小試驗范圍、節約測試成本具有重要意義[9,11]。
[1]譚汝成,歐陽加敏,盧曉莉,等.接種植物乳桿菌和戊糖片球菌發酵對魚鲊品質的影響[J].食品科學,2007,28(12):268-271.
TAN Rucheng,OUYANG Jiamin,LU Xiaoli,et al.Fermentation conditions of Yuzha by inoculated Lactobacillus plantarum and Pediococcus pertosaceus[J].Food Science,2007,28(12):268-271.(in Chinese)
[2]李駿,張春暉,謝華.發酵劑戊糖片球菌I9應用于發酵香腸的效果分析[J].肉類工業,2006(9):36-37.
LI Jun,ZHANG Chunhui,XIE Hua.Application of Pediococcus pentosaceus I 9 in the fermentation sausage[J].Meat Industry,2006(9):36-37.(in Chinese)
[3]張紅星,劉麗,謝遠紅,等.產細菌素的戊糖片球菌的篩選及其細菌素的理化性質研究[J].現代食品科技,2011,27(2):135-138.
ZHANG Hongxing,LIU Li,XIE Yuanhong,et al.Screening of bacteriocin-producing Pediococcus pentosaceus strain and study on physicochemicalcharacteristicsofbacteriocin[J].Modern Food Science and Technology,2011,27(2):135-138.(in Chinese)
[4]Jonganurakkun B,Wang Q,Xu S H,et al.Pediococcus pentosaceus NB-17 for probiotic use[J].Journal of Bioscience and Bioengineering,2008,106(1):69-73.
[5]Xing C F,Hu H H,Huang J B,et al.Diet supplementation of Pediococcus pentosaceus in cobia(Rachycentron canadum)enhances growth rate,respiratory burst and resistance against photobacteriosis[J].Fish&Shellfish Immunology,2013,35:1122-1128.
[6]Dunne C,O’Mahony L,Murphy L,et al.In vitro selection criteria for probiotic bacteria of human origin:correlation with in vivo findings[J].American Journal of Clinical Nutrition,2001,73:386S-392S.
[7]Crociani J,Grill J P,Huppert M,et al.Adhesion of different Bifidobacteria strains to human enterocyte-like Caco-2 cells and comparison with in vivo study[J].Letters in Applied Microbiology,1995,21:146-148.
[8]Li X J,Yue L Y,Guan X F,et al.The adhesion of putative probiotic lactobacilli to cultured epithelial cells and porcine intestinal mucus[J].Journal of Applied Microbiology,2008,104:1082-1091.
[9]Del R B,Busetto A,Vignola G,et a1.Autoaggregation and adhesion ability in a Bifidobacteriumsuis strain[J].Letters in Applied Microbiology,1998,27:307-310.
[10]Collado M C,Meriluoto J,Salminen S.Measurement of aggregation properties between probiotics and pathogens:In vitro evaluation of different methods[J].Journal of Microbiological Methods,2007,71:71-74.
[11]Collado M C,Meriluoto J,Salminen S.Adhesion and aggregation properties of probiotic and pathogen strains[J].European Food Research and Technology,2008,226:1065-1073.
[12]東秀珠,蔡妙英.常見細菌系統鑒定手冊[M].北京:科學出版社,2001.
[13]奧斯伯.精編分子生物學指南[M].第5版.北京:科學出版社,2008.
[14]Rahman M M,Kim W S,Kumura H,et al.Autoaggregation and surface hydrophobicity of bifidobacteria[J].World Journal of Microbiology and Biotechnology,2008,24:1593-1598.
[15]Balcazar J L,Vendrell D.In vitro competitive adhesion and production of antagonistic compounds by lactic acid bacteria against fish pathogens[J].Veterinary Microbiology,2007,122:373-380.
[16]Vesterlund S,Ouwehand A C,Paltta J,et al.Measurement of bacterial adhesion in vitro evaluation of different methods[J]. Journal of Microbiological Methods,2005,60:225-233.
[17]希坎南.伯杰氏細菌鑒定手冊[M].第9版.北京:科學出版社,1989.
[18]Huang J B,Wu Y C,Chi S C.Dietary supplementation of Pediococcus pentosaceus enhances innate immunity,physiological health and resistance to Vibrio anguillarum in orange-spotted grouper(Epinephelus coioides)[J].Fish&Shellfish Immunology,2014,39:196-205.
[19]Shukla R,Goyal A.Novel dextran from Pediococcus pentosaceus CRAG3 isolated from fermented cucumber with anti-cancer properties[J].International Journal of Biological Macromolecules,2013,62:352-357.
[20]Henriksson A,Conway P L.Adhesion of Lactobacillus fermentum 104-S to porcine stomach mucus[J].Current Microbiology,1996,33:31-34.
[21]Del R B,Sgorbati B,Miglioli M,et al.Adhesion,autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum[J]. Letters in Applied Microbiology,2000,31:438-442.
[22]Van Tassell M L,Miller M J.Lactobacillus adhesion to mucus[J].Nutrients,2011(3):613-636.
[23]Rojas M,Ascencio F,Conway P L,et al.Purification and characterization of a surface protein from Lactobacillus fermentum 104R that binds to porcine small intestinal mucus and gastric mucin[J].Applied and Environmental Microbiology,2002,68:2330-2336.
Screening and Molecular Marking of Highly Adhesive Pediococcus pentosaceus and the Characteristics of Their Surface Hydrophobicities and Autoaggregation Abilities
YANG Zhenquan1,2,JIN Caijuan1,ZHANG Mi1,WANG Xiaolin1,GAO Lu1,GU Ruixia1,2
(1.College of Science and Engineering,Yangzhou University,Yangzhou225127,China;2.Jiangsu Key Laboratory of Dairy Biotechnology and Safety Control,Yangzhou 225127,China)
Seven strains of Pediococcus pentosaceus were isolated and identified from the fermented stinky tofu brine using methods of conventional bacteria isolation combined with physiological and biochemical test and 16S rDNA sequencing identification.The specific fingerprint for each strain was developed by the random amplified polymorphic DNA(RAPD)technique.The adhesive ability of the strain and the correlation of phenotypic characteristics of strain adhesion and genotype,surface hydrophobicity,and aggregation ability were studied using Caco-2 cell and intestinal mucus proteinas in vitro model.Results showed that six different fingerprint patterns were found in the seven Pediococcus pentosaceus strains,among them,the A5 subtype of strains(F28-8 and Y27-4)showed the strongest adhesion to the Caco-2 cells and intestinal mucus.Furthermore,these two strains also exhibited highly hydrophobic(the hydrophobicity rate>90%)and higher aggregation ability(the aggregation rate at 2 h>5%).Correlation analysis results showed that the surface hydrophobicity and aggregation ability of Pediococcus pentosaceus were positively related to the adhesive ability to the Caco-2 cell significantly(r=0.900 and 0.792,P<0.05),but they were not significantly correlated to the adhesive ability to the intestinal mucus(r=0.426 and 0.700,P>0.05).The results of this study provide the basis for establishing techniques of rapid screening highly adhesive Pediococcus pentosaceus strains and their colonization and distribution in vivo.
Pediococcuspentosaceus,adhesion,surface hydrophobicity,autoaggregation ability,DNA fingerprint
TS 254
A
1673—1689(2015)09—0926—09
2014-09-30
國家自然科學基金項目(31371806);江蘇省重點實驗室開放課題;江蘇省青藍工程資助項目。
楊振泉(1975—),男,江蘇南通人,農學博士,副教授,主要從事食品微生物資源開發與利用研究。E-mail:yangzq@yzu.edu.cn
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中學生數理化·七年級數學人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(2018年6期)2018-08-16 07:23:10
新高考(英語進階)(2018年1期)2018-04-18 14:00:11
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
