擬絲孢鐮刀菌侵染對馬鈴薯塊莖生理生化特性的影響
2015-08-06 20:30:38趙慧袁強蔣先鋒張元磊史麗娟呂文河李鳳蘭
湖北農業科學 2015年10期
趙慧 袁強 蔣先鋒 張元磊 史麗娟 呂文河 李鳳蘭
摘要:馬鈴薯干腐病是馬鈴薯貯藏期主要病害之一,已經成為馬鈴薯產業發展的一個主要限制因子,確定病原菌的侵染特性對于早期預報和防治干腐病具有重要的意義。對馬鈴薯干腐病主要致病菌擬絲孢鐮刀菌(Fusarium trichothecioides)侵染馬鈴薯過程中薯塊的生理生化特性進行研究。結果表明,不同抗性的馬鈴薯被擬絲孢鐮刀菌侵染后,丙二醛(MDA)含量都出現出下降的趨勢,而超氧化物歧化酶(SOD)活性對薯塊抵抗病原菌的侵染起到了一定的作用,可溶性蛋白質含量和過氧化物酶(POD)活性和抗病性密切相關。
關鍵詞:馬鈴薯抗性;擬絲孢鐮刀菌(Fusarium trichothecioides);超氧化物歧化酶;過氧化物酶;丙二醛
中圖分類號:S435.32 文獻標識碼:A 文章編號:0439-8114(2015)10-2380-04
DOI:10.14088/j.cnki.issn0439-8114.2015.08.019
馬鈴薯是糧食、蔬菜、飼料和工業原料兼用的主要農作物,具有豐產性好、適應性強、經濟效益高和營養豐富的特點,為解決中國經濟欠發達地區的溫飽問題和糧食安全問題做出了重要貢獻,在促進農業結構調整、優化區域布局和抗災救災中發揮了重要作用[1]。近年來,中國馬鈴薯種植面積不斷擴大,種植面積和總產量均占世界的1/4,已成為中國的第四大糧食作物,中國已成為世界馬鈴薯第一生產大國[2]。在北方地區,馬鈴薯無論作為商品薯還是種薯,冬季儲藏方式都是以窖藏為主,在儲藏期間由于儲藏條件的不適宜,常會導致某些病害的加重,如晚疫病、環腐病、干腐病、黑心病及軟腐病等發生,嚴重的會出現爛窖,極大地影響了馬鈴薯的產量和品質,對馬鈴薯的生產和銷售都帶來很大的影響[3-5]。隨著種植面積不斷擴大,馬鈴薯的各種病害日趨嚴重,成為限制馬鈴薯產業發展的主要因素,其中馬鈴薯干腐病是馬鈴薯貯藏期主要病害之一,因馬鈴薯干腐病導致窖貯損失率可高達60%[6],該病已成為馬鈴薯產業進一步發展的瓶頸問題[7]。
前人對馬鈴薯干腐病的研究大部分集中在病原菌的分離鑒定和防治上[8],對于馬鈴薯干腐病和馬鈴薯二者互作機制尚不是十分清楚,對于鐮刀菌(Fusarium sp.)侵染馬鈴薯過程中的細胞、生理生化變化特點及遺傳特性等方面報道較少[9-11]。對馬鈴薯干腐病的防治目前主要采用化學農藥進行化學防治,主要目的是減輕馬鈴薯干腐病的危害程度,難以有效控制病害發生,使用不當還會導致食品安全、馬鈴薯藥害和環境污染問題,尚未找到防治干腐病的有效方法,究其原因是對于防治缺乏理論基礎,存在較嚴重的盲目性。解決馬鈴薯干腐病防治問題行之有效的方法是在充分掌握馬鈴薯與鐮刀菌互作機制的基礎上[12,13],了解馬鈴薯的感病實質,從而選擇切實有效并且可行的防治方法對馬鈴薯干腐病進行防治[14]。
馬鈴薯干腐病是由鐮刀菌引起的,病原菌為鐮刀菌屬(Fusarium)的多種真菌。適宜發病的溫濕度范圍較寬,5~30 ℃均能導致馬鈴薯發病。高溫、高濕有利于發病,在貯藏條件差、通風不良環境利于此病的發生[15],早熟品種比晚熟品種易發病[16]。黑龍江省馬鈴薯干腐病主要病原有6個種及變種,分別為接骨木鐮刀菌(Fusarium sambucinum)、燕麥鐮刀菌(Fusarium avenaceum)、擬絲孢鐮刀菌(Fusarium trichothecioides)、茄病鐮刀菌藍色變種(Fusarium solani var. coeruleum)、茄病鐮刀菌(Fusarium solani)、擬枝孢鐮刀菌(Fusarium sporatrioides)、黃色鐮刀菌(Fusarium culmorum)。其中致病力較強的為接骨木鐮刀菌、燕麥鐮刀菌和擬絲孢鐮刀菌[17,18]。本試驗以擬絲孢鐮刀菌為致病菌,對馬鈴薯與擬絲孢鐮刀菌的互作過程中塊莖主要防御物質——超氧化物歧化酶(SOD)、過氧化物酶(POD)、丙二醛(MDA)和可溶性蛋白質變化進行測定,探討擬絲孢鐮刀菌侵染過程中馬鈴薯的抵御作用,闡述馬鈴薯與鐮刀菌的互作機理,為科學防治馬鈴薯擬絲孢鐮刀菌病害、選育和培育具有廣譜抗病性的馬鈴薯品種提供理論依據。
1 材料與方法
1.1 材料
馬鈴薯選用黑龍江省馬鈴薯主栽品種克山1號(K1,抗性品種)、大西洋(Altantic,感病品種)的脫毒薯塊(原原種)為寄主材料。供試病原菌為東北農業大學生命科學學院實驗室保存的擬絲孢鐮刀菌,保存于PDA培養基上。
1.2 方法
用打孔器在擬絲孢鐮刀菌活化培養基上將菌體做成菌餅待用。在無菌操作臺中將馬鈴薯塊莖表面消毒,去皮后用刀將其切成5 mm的薄片,薄片直徑為3 cm。然后將菌餅放在馬鈴薯薄片中心位置,放入平皿中,以不接種的為對照。 25 ℃恒溫培養。6、12、24、48、72 h取樣,每次取樣1.0 g,3個重復。所取樣品迅速用錫紙包上,做好標記后放入液氮中速凍,保存于-80 ℃冰箱中。采用酶-蛋白聯合測定法[19]測定生理生化指標。
2 結果與分析
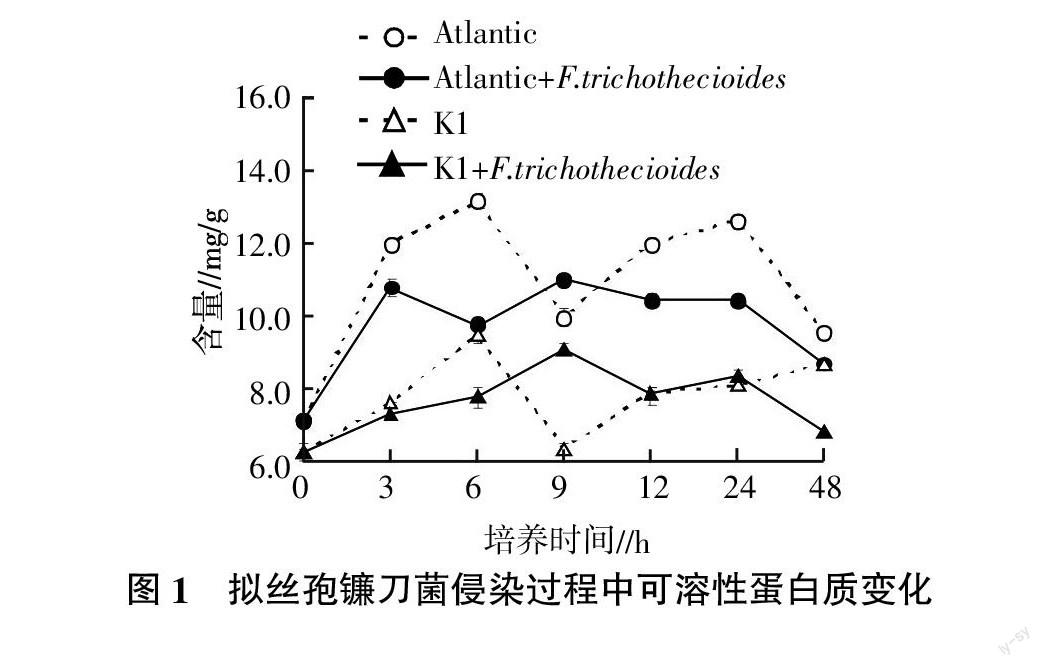
2.1 擬絲孢鐮刀菌侵染過程中可溶性蛋白質含量變化
采用擬絲孢鐮刀菌對大西洋和克山1號的薯塊進行侵染,測定不同時間可溶性蛋白質含量,結果見圖1。由圖1可見,擬絲孢鐮刀菌侵染大西洋薯塊后,薯塊可溶性蛋白質含量僅在9 h時略高于對照,其余時間均明顯低于對照,表現出含量隨侵染時間延續呈降低趨勢。受擬絲孢鐮刀菌侵染的克山1號薯塊可溶性蛋白質含量除了在6 h時明顯低于對照外,其他時期含量與對照比較接近。可見,不同抗性馬鈴薯品種被擬絲孢鐮刀菌侵染后薯塊內可溶性蛋白質含量變化趨勢存在差異,可溶性蛋白質含量可能與馬鈴薯品種對擬絲孢鐮刀菌的抗性呈正相關。
2.2 擬絲孢鐮刀菌侵染過程中SOD活性變化
采用擬絲孢鐮刀菌侵染大西洋和克山1號薯塊,測定不同時間的SOD活性,結果見圖2。擬絲孢鐮刀菌侵染馬鈴薯后,大西洋和克山1號薯塊內的SOD活性在3 h時都表現為降低,然后出現了快速升高,其中克山1號在6 h達到了最高,大西洋在9 h時達到了最高值;在9~48 h抗病品種克山1號薯塊中的SOD活性上升,而感病品種大西洋薯塊中的SOD活性處于下降的趨勢,表明在擬絲孢鐮刀菌侵染馬鈴薯薯塊后,不斷上升的SOD活性對于薯塊抵抗病原菌的侵染起到了一定的作用。
2.3 擬絲孢鐮刀菌侵染過程中POD活性變化
采用擬絲孢鐮刀菌侵染大西洋和克山1號薯塊,測定不同時間的POD活性,結果見圖3。擬絲孢鐮刀菌侵染后大西洋和克山1號薯塊內POD活性在0~6 h呈下降趨勢,大西洋內POD酶活性變化較大。在6 h后,2個品種的POD活性都開始呈現出上升趨勢,在0~12 h抗性品種克山1號POD活性始終高于感病品種大西洋,表明這個階段POD酶活性與抗病性強密切相關。
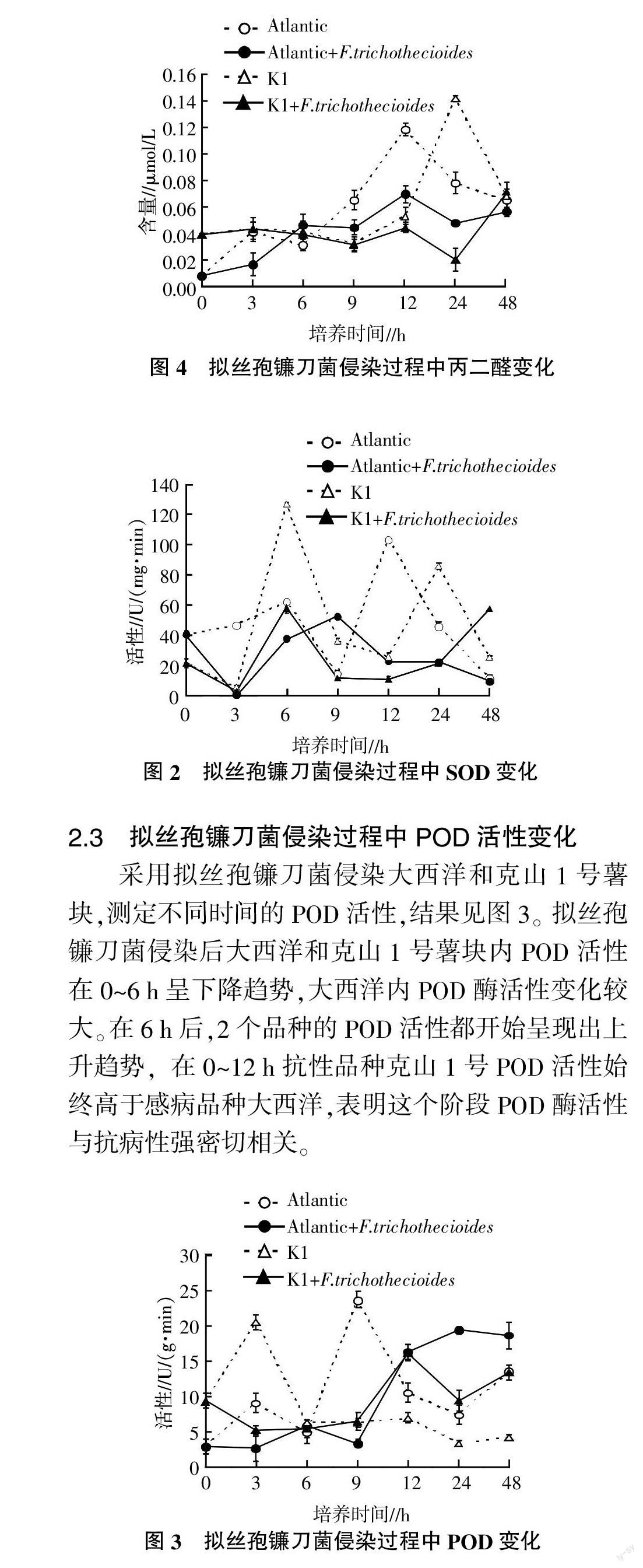
2.4 擬絲孢鐮刀菌侵染過程中丙二醛含量變化
采用擬絲孢鐮刀菌侵染大西洋和克山1號薯塊,測定不同時間的丙二醛含量,結果見圖4。擬絲孢鐮刀菌侵染后,侵染組相對對照組丙二醛含量都出現下降趨勢,其中克山1號中的丙二醛含量的下降幅度較小,只是在24 h時出現了較大幅度下降,而感病品種大西洋薯塊中的丙二醛含量下降的幅度較大,在12 h時已出現了較大幅度下降。丙二醛含量下降的原因可能是在病原菌侵染過程中薯塊中產生了大量的抗氧化酶。
3 小結與討論
活性氧對植物的防御反應十分重要,它能夠誘導局部植物細胞程序性死亡,啟動細胞壁蛋白的氧化交聯,使病原限制在自殺性的寄主細胞內,從而減緩病原的侵染[20]。植物處于逆境下,也會產生自由基反應。胡景江等[21]對元寶楓進行模擬脅迫處理,發現在干旱脅迫條件下元寶楓的SOD活性先下降后逐漸回升至接近正常水平。耐干旱、霜凍、高鹽堿、有毒物質等適應性強的植物內SOD活性明顯較高,SOD與植物耐逆境能力有關,通過對植物SOD的分析有助于了解植物抗逆性,可用于培養抗逆能力強、有經濟效益的新品種[22]。過氧化物酶是植物體內普遍存在的、活性較高的一種酶,它與植物代謝及抗逆性都有密切關系[23,24]。它的作用具有雙重性,一是過氧化物酶可在逆境或衰老初期表達,表現為保護效應;二是過氧化物酶也可在逆境或衰老后期表達,參與活性氧的生成,是植物體衰老的產物,甚至可作為衰老指標[25]。丙二醛是最常用的膜脂過氧化指標[26]。植物在逆境下遭受傷害或衰老,與活性氧積累誘發的膜脂過氧化作用密切相關,膜脂過氧化的產物有二烯軛合物、脂類過氧化物、丙二醛、乙烯等。其中丙二醛是膜脂過氧化最重要的產物之一,可以通過測定丙二醛了解膜脂過氧化的程度以間接了解膜系統受損程度以及植物的抗逆性。
本研究中不同抗性馬鈴薯被擬絲孢鐮刀菌侵染后薯塊內可溶性蛋白質含量變化趨勢存在差異,在感病品種中可溶性蛋白質表現出下降趨勢,可溶性蛋白質含量可能與馬鈴薯品種對擬絲孢鐮刀菌抗性呈正相關。9~48 h抗病品種克山1號薯塊中的SOD活性上升,感病品種大西洋薯塊中的SOD活性處于下降的趨勢,說明SOD對薯塊抵抗病原菌的侵染起到了一定的作用。不同抗性馬鈴薯品種受擬絲孢鐮刀菌侵染時薯塊內POD活性變化差別較大,POD活性與馬鈴薯抗擬絲孢鐮刀菌侵染有密切相關性。相對于未被侵染薯塊,染病的馬鈴薯MDA含量出現了大幅度的下降。馬鈴薯在抵抗擬絲孢鐮刀菌所引起的干腐病過程中防御酶起到了至關重要的作用,被擬絲孢鐮刀菌侵染后抗性品種的各指標測定結果均不同程度地高于未被病原菌浸染時期,表明病原菌侵染后誘導馬鈴薯體內與抗干腐病有關的防御酶活性升高。本研究從防御酶活性變化方面闡明馬鈴薯抗病情況,為馬鈴薯抗病選種提供了理論依據。
參考文獻:
[1] 謝建華.我國馬鈴薯生產現狀及發展對策[J].中國農技推廣,2007(5):4-7.
[2] 孫東升,劉合光.我國馬鈴薯產業發展現狀及前景展望[J].農業展望,2009(3):25-28.
[3] 宋伯符.充滿朝陽的中國馬鈴薯產業[A].馬鈴薯產業與高新技術[C].哈爾濱:哈爾濱工程大學出版社,2002.
[4] 屈冬玉,謝開云,金黎平,等.中國馬鈴薯產業發展與食物安全[J].中國農業科學,2005,38(2):258-262.
[5] 田 芳.北方地區馬鈴薯窖藏病害的發生及防治[J].科技致富向導,2012(27):378-379.
[6] CARNEGIE S F,RUTHVEN A D,LINDSAY D A,et al.Effects of fungicides applied to seed potato at harvest or after grading on fungal storage diseases and plant development[J]. Ann Appl Biol,1990,116:61-72.
[7] 趙生山,牛樂華.馬鈴薯貯藏期病害調查及藥劑防治研究[J].農業科技與信息,2008(11):44-46.
[8] 彭學文,朱杰華.河北省馬鈴薯真菌病害種類及分布[J].中國馬鈴薯,2008,22(1):31-33.
[9] 劉程惠,胡文忠.不同貯藏溫度下鮮切馬鈴薯的生理生化變化[J].食品與機械,2008,24(2):38-42.
[10] 李國龍,吳海霞,溫 麗.作物抗旱生理與分子作用機制研究進展[J].中國農學通報,2010,40(23):185-191.
[11] CHEN Q, SUN H S, YANG J L, et al. Inheritance and molecular mapping of new genes conferring late blight and CPB resistance in Mexican wild potato species Solanum pinnatisectum[J]. Agricultural Science & Technology,2009,10(5):257-261.
[12] ZHANG R J, CHEN Y J, MENG M L, et al.Regulation of nitrogen on potato under NaCl stress[J]. Agricultural Science & Technology,2010,11(11):65-67.
[13] BI Y, LI Y C, GE Y H. Induced resistance in postharvest fruits and vegetables by chemicalsand its mechanism[J]. Stewart Postharvest Review, 2007,12(6):1-7.
[14] 吳 斌,王建富,孫瑞林,等.泰興市馬鈴薯病蟲害發生特點與防治技術[J].現代農業科技,2011(14):179-180.
[15] THEV D J,廖曉蘭.溫度對接種不同鐮刀菌的馬鈴薯干腐病發展的影響[J].國外農學——雜糧作物,1991(6):30-33.
[16] 侯忠艷,馬鈴薯干腐病的發生與防治[J].現代農業科技,2012(10):173,179.
[17] 閔凡祥,王曉丹,胡林雙,等.黑龍江省馬鈴薯干腐病菌種類鑒定及致病性[J].植物保護,2010,36(4):112-115.
[18] 李鳳蘭,蔣先鋒,史麗娟,等.兩株馬鈴薯干腐病病原菌的分離和鑒定[J].作物雜志,2013(4):125-127.
[19] 汪俏梅,郭得平,KYIKYI W.1-MCP對青花菜貯藏壽命、品質和抗氧化酶活性的影響[J].園藝學報,2004,31(2):205-209.
[20] BRISSON L F,TENHAKEN R,LAMB C J.Function of oxidative cross-linking of cell wall structural proteins in plant disease resistance[J].Plant Cell,1994,6(12):1703-1712.
[21] 胡景江,劉樹滔,潘劍茹.SOD特性改善技術研究進展[J].海峽藥學,2009,21(2):11-14.
[22] 李金華,王 豐,廖亦龍.水稻種子活力的生理生化及遺傳研究[J].分子植物育種,2009,7(4):772-777.
[23] 王晶英,趙雨森,王 臻,等.干旱脅迫對銀中楊生理生化特性的影響[J].水土保持學報,2006,20(1):197-200.
[24] 繳麗莉,倪志云,路丙社,等.低溫脅迫對青榨槭幼樹抗寒指標的影響[J].河北農業大學學報,2006,29(4):44-47.
[25] 張蜀寧,張振超,張紅亮,等.低溫脅迫對不同倍性不結球白菜生長及生理生化特征的影響[J].西北植物學報,2008,28(1):109-112.
[26] 趙世杰,許長成,鄒 琦,等.植物組織中丙二醛測定方法的改進[J].植物生理學通訊,1994,30(3):207-210.
