烏珠穆沁肉羊自然放牧條件下肌內脂肪相關基因表達分析
2015-05-30 10:48:04曹欣宇梁圖雅齊志欣斯琴其木格格日勒圖
肉類研究 2015年7期
曹欣宇 梁圖雅 齊志欣 斯琴其木格 格日勒圖

摘 要:為研究自然放牧條件下不同月齡烏珠穆沁肉羊的股二頭肌、肱三頭肌、背最長肌3 個部位骨骼肌肌內脂肪相關基因的表達,采用實時熒光定量聚合酶鏈式反應(polymerase chain reaction,PCR)技術測定肉羊生長過程中過氧化物酶體增殖劑激活受體γ亞型(peroxisome proliferator activated receptors,PPARγ)和脂肪酸合成酶(fatty acid synthase,FAS)相關基因(PPARγ、FAS)表達量。結果表明:兩種基因表達量在各部位骨骼肌上的趨勢相近。PPARγ基因的表達量在1月齡和12月齡較高,9月齡和18月齡較低;FAS基因的表達量在3月齡和12月齡較高,
9月齡和18月齡較低。通過對基因表達量與肌內脂肪的相關性分析可以得出,PPARγ基因和FAS基因的表達量與肌內脂肪含量呈負相關。
關鍵詞:肉羊;肌內脂肪;PPARγ基因;FAS基因
Expression of Genes Related to Intramuscular Fat Content in Wuzhumuqin Sheep under Natural Grazing Conditions
CAO Xinyu, LIANG Tuya, QI Zhixin, Siqinqimuge, BORJIGIN Gerelt*
(College of Food Science and Engineering, Inner Mongolia Agricultural University, Hohhot 010018,China)
Abstract: To investigate the relationship between intramuscular fat contents and the related genes, biceps femoris (BF), triceps brachii (TB) and longissimus dorsi (LD) muscles were obtained from naturally grazing Wuzhumuqin sheep, and gene expression related to intramuscular fat contents were measured by real-time polymerase chain reaction (PCR). The results showed that the expression patterns of the peroxisome proliferator activated receptors (PPARγ) and fatty acid synthase (FAS) genes were similar in the three skeletal muscles. The PPARγ gene was highly expressed at 1 and 12 months of age but lowly expressed at 9 and 18 months of age, whereas the FAS gene was highly expressed at 3 and 12 months of age and also lowly expressed at 9 and 18 months of age. The correlation analysis revealed that there was a negative correlation between intramuscular fat contents and the expression of the PPARγ and FAS genes in skeletal muscle.
Key words: sheep; intramuscular fat; PPARγ gene; FAS gene
中圖分類號:Q812 文獻標志碼:A 文章編號:1001-8123(2015)07-0025-05
doi: 10.7506/rlyj1001-8123-201507006
羊肉具有膽固醇含量低,營養價值高,味美多汁等品質特點[1-2]而受到廣大群眾的喜愛。影響羊肉品質的因素有很多,其中肌內脂肪含量是一個重要因素,肌肉不同部位脂肪含量的高低直接影響羊肉的風味、嫩度、多汁性等。肌內脂肪含量由年齡、環境、飼養以及肌內脂肪相關基因等多種因素來決定。參與基因調控的主要物質有過氧化物酶體增殖劑激活受體(peroxisome proliferator activated receptors,PPARs)[3-5]和脂肪酸合成酶(fatty acid synthase,FAS)。PPARs包括PPARα、PPARβ、PPARγ 3種亞型[4,6],能調控脂肪基因的表達并能使其脂肪細胞分化,調節脂類代謝相關基因和參與脂肪的吸收、運輸、形成和分解過程[7]。FAS是機體內合成脂肪途徑中的一個關鍵性的酶。FAS的表達可增加在體內甘油三酯的沉積而導致肥胖[8]。兩種基因的表達在不同品種及不同骨骼肌組織中脂肪沉積存在著差異性。張長慶等[9]研究指出PPARγ基因在不同部位脂肪組織和肌肉組織中均有表達且部位不同而表達量具有差異性。Lee等[10]
研究指出PPAR基因的相對表達量在韓國牛27月齡的表達與其在12月齡時的韓國牛的相對表達量相比有3倍的關系。Clarke等[11]研究指出用高碳水化合物的日糧能提高其肝臟中FAS基因的表達,而高蛋白的日糧飼喂會降低FAS基因的表達。QiaoYong等[12]用實時熒光定量PCR法檢測肌肉中FAS基因表達的發育性變化,發現FAS mRNA水平在新疆細毛羊和哈薩克羊肌肉中的發育模式各有不同,FAS基因mRNA的表達量與哈薩克羊肌內脂肪含量負相關。由于季節不同肉羊采食的牧草不同,肉羊在不同季節和生長階段中肌內脂肪含量大不相同。梁圖雅等[13]對烏珠穆沁肉羊不同月齡不同部位骨骼肌脂肪含量等進行了系統的研究,在此基礎上本研究對自然放牧條件下烏珠穆沁肉羊不同月齡、不同骨骼肌部位的肌內脂肪相關基因表達量進行分析,更加深入的探討自然放牧條件下季節變換對肉羊生長過程中肌內脂肪的影響。
1 材料與方法
1.1 材料與試劑
冬季出生并自然放牧條件的1、3、6、9、12、18月齡烏珠穆沁肉羊中各選3只。屠宰后采取股二頭肌、肱三頭肌、背最長肌的骨骼肌,小塊分割后立即用液氮冷凍并密封保存備用。
Trizol RNA抽提試劑 美國TaKaRa試劑公司;BioRT Two Step反轉錄試劑盒、2×real time PCR Master Mix熒光定量PCR試劑、核酸燃料 北京百泰克生物有限公司;異丙醇、氯仿、無水乙醇、焦炭酸二乙酯 北京
全式金生物技術有限公司;5×TBE緩沖液、6×上樣緩沖液溴酚蘭、DNA Maker 天根生化科技有限公司。
1.2 儀器與設備
ND-1000型微量紫外分光光度計 美國NanoDrop公司;GDS-8000凝膠成像系統 美國UVP公司;高速冷凍離心機 湖南湘儀離心機儀器有限公司;電泳儀 北京百晶生物技術有限責任公司;CFX96TM實時熒光定量PCR儀 美國Bio-Rad公司。
1.3 方法
1.3.1 引物設計
目的基因PPARγ上下游引物序列參考Huang等[14]的方法,目的基因FAS上下游序列參考張英杰等[15]的方法。內參基因采用β-actin,其上下游引物序列參考吳瓊彪等[16]的方法。引物如表1所示。
表 1 PPARγ、FAS基因引物序列
1.3.2 RNA的提取
取一定肉樣在液氮中磨碎,約100 mg組織加入1mL裂解液。在25 ℃放置5 min,將其裂解細胞與溶劑混勻使核酸蛋白復合物完全分離。之后將樣品4℃、12 000×g離心10 min,取上清液。上清液中加入0.2 mL氯仿,振蕩15s,室溫放置3 min。4 ℃、12 000×g離心15 min。待出現分層,分別為黃色有機相底層,無色水相上層以及中間層。小心提取含有RNA的水相轉移到新管中,用冰異丙醇沉淀水相中的RNA,每使用1 mL Trizol加入0.5 mL異丙醇,低溫放置10 min。4 ℃、12 000×g離心10 min,將上清液移去。75%乙醇洗滌RNA沉淀,每使用1 mL Trizol至少加1 mL 75%乙醇。4 ℃、7 500×g離心5 min,棄上清液。室溫放置干燥RNA沉淀。再加25 μL無RNase水,輕搖RNA溶解。-80 ℃保存備用。
1.3.3 RNA濃度純度檢測
使用紫外分光光度計,用DEPC處理水校正零點,讀取RNA樣品OD260 nm/280 nm的比值及濃度。
1.4 數據分析
實時熒光定量PCR反應所得到的Ct值用運Excel軟件進行計算其各自的表達量并用SAS軟件進行顯著性和與肌內脂肪含量的相關性分析。
2 結果與分析
2.1 烏珠穆沁肉羊不同月齡、不同骨骼肌部位PPARγ和FAS基因表達量
2.1.1 不同月齡、不同骨骼肌部位PPARγ基因表達
小寫字母不同,表示相同部位、不同月齡間差異顯著(P<0.05);大寫字母不同,表示相同月齡、不同部位骨骼肌間差異顯著(P<0.05)。下同。
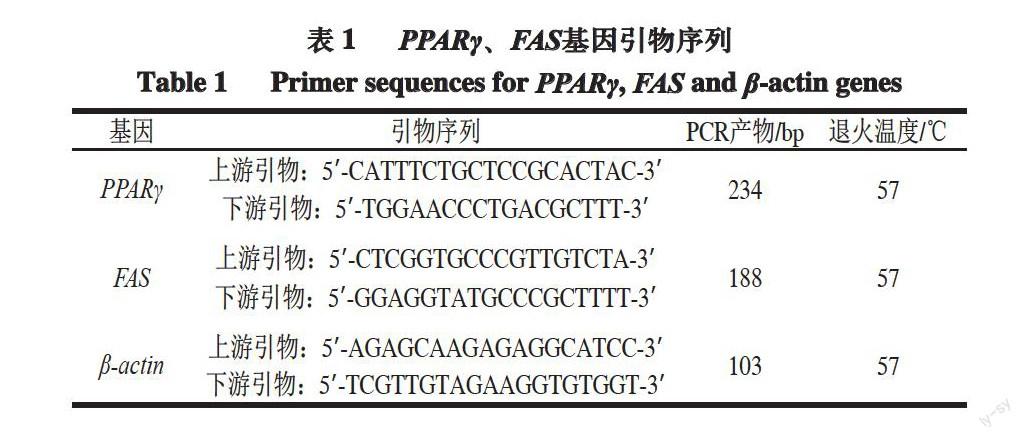
圖 1 肉羊不同月齡不同骨骼肌部位PPARγ基因表達量
Fig.1 Relative expression levels of PPARγ gene in different muscles at different months of age
由圖1可知,同一部位骨骼肌不同月齡之間的對比可以得出:對于背最長肌,肉羊PPARγ表達量1月齡和3月齡之間無顯著差異;6月齡相對于3月齡其表達量顯著提高(P<0.05);而9月齡的表達量最低,12月齡的表達量最高,均與1、3、6、9、18月齡間存在顯著差異
(P<0.05);18月齡表達量與1月齡和6月齡表達量無顯著差異,但比3月齡顯著較高(P<0.05)。
對于股二頭肌,肉羊1月齡PPARγ的表達量最高,與3、6、9、12、18月齡間存在顯著差異(P<0.05);3月齡和6月齡PPARγ的表達量沒有顯著差異,且均顯著高于9月齡PPARγ的表達量;12月齡PPARγ的表達量顯著高于3、6、9、18月齡的表達量(P<0.05);18月齡PPARγ的表達量與9月齡沒有顯著差異,均顯著低于1、3、6、12月齡的表達量(P<0.05)。
對于肱三頭肌,1月齡PPARγ的表達量高于6月齡,但無顯著性差異,1月齡PPARγ的表達量顯著高于3月齡
(P<0.05);3、6、9月齡PPARγ的表達量沒有顯著差異;12月齡PPARγ的表達量顯著高于1、3、6、9、18月齡(P<0.05);18月齡PPARγ的表達量其顯著低于1、3、6、9、12月齡(P<0.05)。
同一個月齡段不同骨骼肌部位之間的對比可以得出:1月齡股二頭肌的PPARγ基因表達量最高、肱三頭肌次之、背最長肌最低,且3個部位間均存在顯著差異
(P<0.05),背最長肌的表達量顯著低于其他兩個部位(P<0.05);3、6、9月齡背最長肌、股二頭肌間PPARγ的表達量沒有顯著性差異,肱三頭肌的表達量顯著高于背最長肌和股二頭肌兩個部位(P<0.05);12月齡肱三頭肌PPARγ的表達量顯著高于其他兩個部位(P<0.05),背最長肌的表達量顯著高于股二頭肌
(P<0.05)。18月齡背最長肌和肱三頭肌表達量沒有顯著差異,均顯著高于股二頭肌表達量(P<0.05)。
2.1.2 不同月齡、不同骨骼肌部位FAS基因表達量
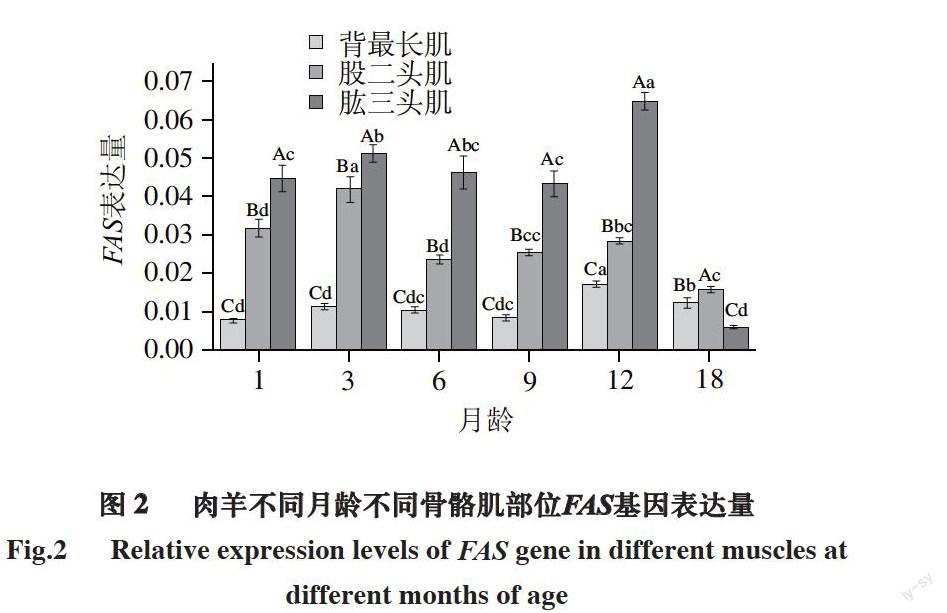
圖 2 肉羊不同月齡不同骨骼肌部位FAS基因表達量
Fig.2 Relative expression levels of FAS gene in different muscles at different months of age
由圖2可知,同一部位骨骼肌不同月齡之間的對比可以得出:對于肉羊背最長肌,3月齡、6月齡和12月齡之間FAS基因表達量無顯著差異,但1月齡、9月齡、12月齡之間差異顯著(P<0.05)。1月齡FAS的表達量極顯著低于3月齡和6月齡((P<0.05),3月齡表達量顯著高于9月齡,6月齡與9月齡之間無顯著差異。1月齡與9月齡表達量無顯著差異,9月齡表達量顯著低于12月齡(P<0.05),18月齡相比于12月齡,表達量顯著低
(P<0.05)。
對于股二頭肌,1月齡FAS的表達量顯著低于3月齡(P<0.05),3月齡的表達量最高,與1、6、9、12、18月齡間均具有顯著差異(P<0.05);6月齡FAS的表達量稍低于9月齡FAS的表達量,而無顯著差異;9月齡FAS的表達量低于12月齡FAS的表達量,但無顯著差異;12月齡FAS的表達量顯著高于18月齡FAS的表達量(P<0.05)。
對于肱三頭肌,1月齡FAS的表達量顯著低于3月齡FAS的表達量(P<0.05),3月齡FAS的表達量相對高于6月齡FAS的表達量,但無顯著差異,6月齡FAS的表達量高于9月齡FAS的表達量,但無顯著差異,9月齡FAS的表達量顯著低于12月齡FAS的表達量(P<0.05),18月齡FAS的表達量顯著低于1、3、6、9和12月齡FAS的表達量(P<0.05)。
同一個月齡段不同骨骼肌部位之間的對比可以得出:1、3、6、9、12月齡時肉羊的肱三頭肌FAS的表達量最高,股二頭肌FAS的表達量次之,背最長肌FAS的表達量最低,而且互相之間均存在顯著差異(P<0.05);18月齡背最長肌FAS的表達量最高、股二頭肌FAS的表達量次之、肱三頭肌FAS的表達量最低,且均存在顯著差異(P<0.05)。
2.2 烏珠穆沁肉羊肌內脂肪含量與PPARγ、FAS基因表達量及相關性分析
2.2.1 肌內脂肪含量與PPARγ、FAS基因表達量
表 2 肉羊不同月齡不同骨骼肌部位肌內脂肪含量與PPARγ、FAS基因表達量
由表2可知,PPARγ基因在9月齡和18月齡時表達相對較低,在12月齡時表達水平達到最高水平。FAS基因3月齡和12月齡表達量較高,18月齡表達量較低。2個基因的表達量與肌內脂肪含量的趨勢正好相反,即與肌內脂肪含量呈負相關。
2.2.2 肌內脂肪含量與與PPARγ、FAS基因表達量相關性分析
由表3可知,在背最長肌部位上,PPARγ基因表達量與肌內脂肪含量之間呈負相關;在股二頭肌上,其PPARγ基因表達量與肌內脂肪含量之間呈負相關;在肱三頭肌上,其PPARγ基因表達量與肌內脂肪含量之間呈負相關。在背最長肌部位上,其FAS基因表達量與肌內脂肪含量之間呈負相關;在股二頭肌部位上,其FAS基因表達量與肌內脂肪含量之間呈負相關;肱三頭肌部位上,其FAS基因表達量與肌內脂肪含量之間的相關系數R為-0.980,在極顯著水平上呈負相關。
表 3 肌內脂肪含量與PPARγ、FAS基因表達量相關性分析
3 討 論
3.1 自然放牧條件下肉羊不同月齡,不同部位骨骼肌PPARγ基因與肌內脂肪的相關性
肉羊在自然放牧的條件下PPARγ基因表達量隨月齡的變化在背最長肌、股二頭肌和肱三頭肌3個部位上的均有相同趨勢。在9月齡和18月齡時表達量相對較低,而12月齡時達到最高水平。肉羊1月齡時,PPARγ基因表達量相對較高,其主要原因有可能是肉羊剛出生PPARγ基因要參與脂肪細胞的形成。3月齡時的肉羊通過運動、骨骼肌生長等原因,與1月齡相比較其PPARγ基因表達量變化較小。當肉羊6月齡時正值夏季,草原上牧草生長茂盛,PPARγ基因表達增加會使肌內脂肪增加。9月齡時牧草旺盛,肉羊肌內脂肪達到較高水平,因牧草旺盛機體本身具有較高的脂肪含量,而自身PPARγ基因表達量會降低并且不會過多表達使肌內脂肪沉積。12月齡的肉羊正處于冬季,因冬天牧草枯黃稀少,自身需要分化前脂肪細胞為脂肪細胞,因而PPARγ基因表達水平較高,使肌內脂肪含量保持在較低水平。18月齡時的肉羊又處于夏季,牧草旺盛,PPARγ基因表達量較低,原因可能是機體需要蓄積脂肪。因此,自然放牧的肉羊的PPARγ基因表達量以及肌內脂肪含量可能與牧草之間有很大的相關性。
在肉羊生長各個階段,肌內脂肪含量與PPARγ基因表達量之間的相關性分析可知,背最長肌、股二頭肌和肱三頭肌部位上均存在負相關。這可能與PPARγ基因自身表達調控和本身脂肪豐富有一定的作用。有研究報道蒙古羊90~160日齡期間PPARγ基因與肌內脂肪含量相關系數為-0.884 9(P=0.1084)[17]。Huang等[14]研究發現在哈薩克羊中PPARγ基因在2~90日齡間的表達量與肌內脂肪含量呈負相關,其相關系數為-0.835(P<0.01)。郝稱莉等[18]研究發現背最長肌PPARγ基因的表達量在4~12 月齡期間與肌內脂肪含量呈負相關,相關系數為-0.441(P=0.076),腰大肌PPARγ基因的表達量在2~12月齡期間與肌內脂肪含量呈負相關,相關系數為-0.120,股二頭肌PPARγ基因的表達量在5~6月齡期間與肌內脂肪含量呈負相關,相關系數為-0.301。本實驗中PPARγ基因與肌內脂肪含量相關性支持以上研究結果。
3.2 自然放牧條件下肉羊不同月齡不同部位骨骼肌FAS基因與肌內脂肪的相關性
自然放牧飼養條件下肉羊FAS基因隨月齡變化的表達量為1月齡時背最長肌、股二頭肌和肱三頭肌部位上FAS基因表達量都在較低水平上。3月齡時肉羊FAS基因表達量升高,6月齡時肉羊FAS基因表達量在股二頭肌部位上顯著降低。
肉羊FAS基因表達量在1月齡時較低,原因可能是剛出生時主要以PPARγ等基因來調控脂肪細胞的形成。3月齡和12月齡的肉羊FAS基因表達量與1、6、9、18月齡時相比均偏高,而3月齡和12月齡的肉羊肌內脂肪含量均偏低,此原因可能與季節環境因素有關。又因3月齡和12月齡時冬季牧草缺少,肉羊機體需自身將葡萄糖等能量轉變為脂肪,此過程中的最后一步合成脂肪酸時會需要FAS,因此機體本身的FAS基因表達量會增加,所以3月齡和12月齡時FAS基因表達量較高。9月齡和18月齡時牧草旺盛,機體自身的脂肪含量較多,所以肉羊FAS基因表達量較低。而6月齡的表達量與3月齡的表達量相比較低,可能與采食到的牧草量的增加有關。因此FAS基因表達量與肌內脂肪含量的相關關系為負相關。姚焰礎[19]認為10~40 kg蘇尼特羔羊肌內FAS基因持續表達,其表達量隨體質量增加而下降,30 kg和40 kg時的表達量顯著低于10 kg和20 kg(P<0.05),與本實驗結果相同。
因此,自然放牧條件下肉羊的脂肪沉積和相關基因(PPARγ和FAS基因)的表達跟肉羊的生長(骨骼肌發達)、牧草以及飼養環境條件有著密切的關聯性。
4 結 論
自然放牧條件下,肉羊與肌內脂肪相關的PPARγ和FAS基因表達量會隨著月齡的增加、季節的變化而變化。肉羊PPARγ基因隨著月齡增加的表達量為1月齡(剛出生)和12月齡(牧草枯竭)時表達較高,9月齡和18月齡(牧草旺盛)時表達較低。PPARγ和FAS基因相對表達量與肌內脂肪含量呈負相關。
參考文獻:
[1] 梁富武. 烏珠穆沁羊的選育攻關[J]. 中國牧業通訊, 2010, 17: 38-39.
[2] 楊富民. 國內羊肉品質研究進展[J]. 甘肅科技, 2003, 19(2): 33-34.
[3] ENSER M, HALLETT K G, HEWETT B, et al. Fatty acid content and composition of UK beef and lamb muscle in relation to production system and implications for human nutrition[J]. Meat Science, 1998, 49(3): 329-341.
[4] LEMBERGER T, DESVERGNE B, WAHLII W. Peroxisome proliferator-activated receptors: a nuclear receptor signaling pathway in lipid physiology[J]. Annual Review of Cell and Developmental Biology, 1996, 12(1): 335-363.
[5] SCHOONJAN K, STAELS B, AUWERX J. Role of the peroxisome proliferator-activated receptor (PPAR) in mediating the effects of fibrates and fatty acids on gene expression[J]. Journal of Lipid Research, 1996, 37(5): 907-925.
[6] BRAISSANT O, FOUFELLE F, SCOTTO C, et al. Differential expression of peroxisome proliferator-activated receptors (PPARs): tissue distribution of PPARα, PPARβ and PPARγ in theadultrat[J]. Endocrinology, 1996, 137(1): 354-366.
[7] DEVINE J H, EUBANK D W, CLOUTHIER D E, et al. A dipose expression of the phosphoenolpyruvate carboxykinase promoter requires peroxisome proliferator-activated receptor γ and 9-cis-retinoic acid receptor binding to an adipocyte-specific enhancer in vivo[J]. Journal of Biological Chemistry, 1999, 274(19): 13604-13612.
[8] YAN Xinchun, WANG Yizhen, XU Zirong. Regulation of fatty acid synthase (FAS)gene expression in animals[J]. Acta Zoonutrimenta Sinica, 2002, 14(2): 1-4.
[9] 張長慶, 劉婷, 趙生國. PPARγ基因在荷斯坦公牛不同組織部位表達差異性研究[J]. 中國農學通報, 2014, 30(23): 13-18.
[10] LEE S H, PARK E W, CHO Y M, et al. Identification of differentially expressed genes related to intramuscular fat development in the early and late fattening stages of hanwoo steers[J]. Journal of Biochemistry and Molecular Biology, 2007, 40(5): 757-764.
[11] CLARKE S D, AMSTRONG M K, JUMP D B. Nutritional control of rat liver fatty acid synthase and S14 mRNA abundance[J]. The Journal of Nutrition, 1990, 120(2): 218-224.
[12] QIAO Yong, HUANG Zhiguo, XIE Zhuang, et al. Developmental changes of the FAS and HSL mRNA expression and their effects on the content of intramuscular fat in Kazak and Xinjiang sheep[J]. Journal of Genetics and Genomics, 2007, 34(10): 909-917.
[13] 梁圖雅, 額日克古麗, 格日勒圖. 烏珠穆沁羊生長過程中食肉營養成分變化分析[J]. 肉類研究, 2013, 27(7): 30-34.
[14] HUANG Z G, XIONG L, LIU Z S, et al. The developmental changes and effect on IMF content of H-FABP and PPARγ mRNA expression in sheep muscle[J]. Acta Genetica Sinica, 2006, 33(6): 507-514.
[15] 張英杰, 劉月琴, 劉景云. 日糧不同蛋白水平對綿羊脂肪和肌肉中FAS基因表達的影響[J]. 畜牧獸醫學報, 2010, 41(7): 829-834.
[16] 吳瓊彪, 格日勒圖. 不同月齡烏珠穆沁羊骨骼肌中Myostatin表達量變化規律分析[D]. 呼和浩特: 內蒙古農業大學, 2013.
[17] 國家加工食品質量監督檢驗中心(廣州), 廣州市產品質量監督檢驗所等. GB/T 9695.18—2008 肉與肉制品 總灰分測定[S]. 北京: 中國標準出版社, 2008.
[18] 郝稱莉. 湖羊肌肉組織H-FABP和PPARγ基因表達水平與肌內脂肪含量的相關研究[J]. 中國農業科學, 2008, 41(11): 3776-3783.
[19] 姚焰礎. 蘇尼特羔羊肌內脂肪細胞分化相關因子和肌內脂肪代謝關鍵酶基因表達發育規律的研究[D]. 呼和浩特: 內蒙古農業大學, 2008.
