巴西橡膠樹HbMAPK1基因的克隆及表達分析
2015-05-30 20:45:32吳紹華等
熱帶作物學報 2015年9期
吳紹華等

摘 要 植物促分裂素原活化蛋白激酶(Mitogen-activated Protein Kinase,MAPK)信號轉導途徑在植物應對低溫、鹽害、蟲害等逆境脅迫中起著重要的作用。采用RACE和RT-PCR技術從橡膠樹熱研7-33-97的膠乳cDNA中擴增到一條全長cDNA,共1 909 bp,包含1 119 bp開放閱讀框,編碼372個氨基酸,含有特征性的STKc_TEY_MAPK和S_TKc結構域,命名為HbMAPK1(GenBank:KP262500),其編碼蛋白分子量為42.57 ku,理論等電點為7.21。熒光定量PCR結果顯示,HbMAPK1在橡膠樹的根、樹皮、膠乳及葉片中均有表達,其中穩(wěn)定期葉片的基因表達量最高。4 ℃低溫處理條件下,HbMAPK1在93-114與熱墾501無性系葉片間的基因表達模式存在明顯差異,表現為93-114葉片中HbMAPK1的表達受4 ℃低溫的誘導顯著上調,熱墾501葉片中的表達下調。結合2個品系在生產上的抗寒表現,即93-114屬于抗寒性品系,抵御低溫能力較強,而熱墾501屬于不抗寒性品系,對低溫較敏感。因此,推測HbMAPK1可能與橡膠樹的抵御溫脅迫有關。
關鍵詞 巴西橡膠樹;HbMAPK1;基因克隆;表達分析
中圖分類號 S749.1 文獻標識碼 A
Molecular Cloning and Expressional Analysis of HbMAPK1 from Rubber Tree(Hevea brasiliensis Müll. Arg.)
WU Shaohua1, WANG Jianxiao1,2, CHEN Yueyi1, LI Mengfang2, TIAN Weimin1*
1 Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences, Minstry of Agriculture Key Laboratory of Rubber
Biology/State Key Laboratory Incubation Base for Cultivation & Physiology of Tropical Crops, Danzhou, Hainan 571737, China
2 College of Agriculture, Hainan University, Haikou, Hainan 570228, China
Abstract Mitogen-activated Protein Kinase(MAPK) signaling pathways play important role in response to the stress such as low temperature, salt and pest. In this study, the full length cDNA of HbMAPK1(GenBank:KP262500)was obtained from cDNA library in the latex of rubber tree clone Reyan7-33-97. It was 1 909 base pairs(bp) in length, containing a 1 119 bp open reading frame(ORF), encoding 372 amino acid residues with a predicted molecular mass of 42.57 ku and a pI of 7.21. The deduced amino acid sequence contained two specific domains of STKc_TEY_MAPK and S_TKc domain. Real-time PCR analysis showed that HbMAPK1 expressed in root, bark, latex and leaves. The transcript level of mature leaf was the highest. The expression patterns of HbMAPK1 was different in 4 ℃-treated leaf of rubber tree clone 93-114 and Reken 501. The HbMAPK1 transcript levels were upregulated in leaf by 4 ℃ treatment for 2 hours in rubber tree clone 93-114, but the result was the opposite in rubber tree clone Reken501. Based on the differential cold resistance of the two rubber tree clones in pructice,i. e., the cold resistance of 93-114 was stronger than Reken 501, HbMAPK1 is supposed to haer a role in enhancing cold resistance of rubber tree.
Key words Hevea brasiliensis Müll. Arg.; HbMAPK1;Gene clone;Expression analysis
doi 10.3969/j.issn.1000-2561.2015.09.006
促分裂原活化蛋白激酶(Mitogen-activated Protein Kinase,MAPK)是一類傳遞胞外和胞內信號到細胞響應的絲氨酸/蘇氨酸蛋白激酶,是真核生物信號傳遞網絡中的重要途徑之一,在真核生物中普遍存在。MAPK級聯(lián)途徑作為多種生長信號跨膜傳遞的交匯點或共同通路,可將外源信號轉入胞內并引發(fā)細胞應答,調控細胞的基因表達,在植物的生長發(fā)育和抗逆響應中發(fā)揮著極為重要的作用[1-5]。
MAPK級聯(lián)途徑由促分裂原活化蛋白激酶(MAPK)、促分裂原活化蛋白激酶的激酶(MAPKK)、促分裂原活化蛋白激酶的激酶之激酶(MAPKKK)3個組分組成[6],由MAPKKK-MAPKK-MAPK依次磷酸化傳遞信號[7-8]。MAPK級聯(lián)途徑各激酶基因家族在不同植物物種中是保守的。植物MAPKKK磷酸化MAPKK的S/T-X5-S/T(X代表任意氨基酸)序列,MAPKK磷酸化MAPK的保守氨基酸基序TDY或TEY中的蘇氨酸(T)和酪氨酸(Y),使其激活。T(D/E)Y位于VII和VIII亞區(qū)之間的活化環(huán)。活化的MAPK除了可激活轉錄因子,也可通過蛋白質之間的相互作用引起細胞響應[6]。MAPK級聯(lián)系統(tǒng)通過參與蛋白質磷酸化作用傳遞和放大信號,調控下游基因的表達,最終引起植物的一系列生理反應[9]。
橡膠樹原產于高溫、高濕、靜風的巴西亞馬遜河流域,是典型的熱帶雨林樹種,抵御低溫的能力不強,當氣溫降至10 ℃以下時,橡膠樹的光合作用就停止了,氣溫降至5 ℃以下時就開始受害。中國的植膠區(qū)位于北緯18~24°熱帶北緣和南亞熱帶(主要是海南、云南和廣東),寒潮是危害中國橡膠樹最為嚴重的自然災害之一。因此,選育抗寒高產的橡膠樹無性系是中國橡膠樹育種的重要目標。目前,抗寒性種質的鑒定主要采用抗寒前哨梯度苗圃系比鑒定法,即在一般年份有寒流侵襲的地區(qū)或寒流通道上建立前哨梯度苗圃系比區(qū)。通過抗寒苗圃鑒定,也選育出一批抗輻射或抗平流低溫能力強或較強的無性系[10]。但抗寒前哨梯度苗圃系比鑒定法也存在一定的缺陷,如受寒流類型及方向的不確定性,種植區(qū)域的小環(huán)境氣候差異等會造成鑒定的效率低、周期長。而且,一旦遇到暖冬,將難以比較無性系間耐寒性的差異,給橡膠樹抗寒選育種工作造成時間和物力的浪費。因此,在抗寒相關基因的研究基礎上,開發(fā)抗寒性分子標記輔助抗寒前哨苗圃鑒定,將大幅度提高鑒定的效率,推進抗寒性品種的選育進程。本研究從橡膠樹膠乳中分離鑒定了一個低溫響應的新的MAPK基因——HbMAPK1,旨在為進一步探討橡膠樹抗逆機理及開發(fā)抗逆性分子標記輔助橡膠樹抗逆育種奠定基礎。
1 材料與方法
1.1 材料
實驗材料巴西橡膠樹(Hevea brasiliensis Müll. Arg.)無性系熱研7-33-97,種植于中國熱帶農業(yè)科學院實驗場。巴西橡膠樹無性系93-114、熱墾501為當年芽接苗。RNAprep Pure多糖多酚植物總RNA提取試劑盒,DNaseⅠ購自天根公司。RACE試劑盒SMARTerTM RACE cDNA Amplification Kit購自clontech公司。反轉錄試劑盒RevertAidTM First Strand cDNA Synthesis Kit購自Ferments公司。DNA凝膠回收試劑盒購自OMEGA公司。PyrobestTM DNA Polymerase、實時熒光定量PCR試劑SYBR Premix Ex TaqⅡ(Tli RNaseH Plus)購自大連寶生物公司。pEASY-Blunt Simple Cloning Kit購自北京全式金公司。其它生化試劑均為進口或國產分析純試劑。引物合成和DNA測序由Invitrogen公司完成。
1.2 方法
1.2.1 材料處理 無性系熱研7-33-97的根、皮、膠乳、葉片的RNA用于HbMAPK1的組織特異性相對表達分析;橡膠樹無性系93-114和熱墾501用于4 ℃低溫脅迫處理2、4、8、12、24、48 h,不處理作為對照樣品,每個處理設3個重復,取每個時間點葉片混合提取RNA,分析低溫脅迫處理對HbMAPK1基因表達的影響。
1.2.2 總RNA的提取與cDNA的合成 橡膠樹不同組織總RNA的提取參照RNAprep Pure多糖多酚植物總RNA提取試劑盒的操作說明進行。用DnaseⅠ柱上消化RNA樣品中殘留的微量DNA,cDNA第一鏈的合成根據Ferments公司試劑盒的操作步驟進行。
1.2.3 HbMAPK1的RACE和全長cDNA擴增 搜索本實驗室的轉錄組數據庫,獲得1個注釋為MAPK的Unigene,采用RACE技術進行3′和5′端序列的擴增。RACE 擴增參照clontech公司的試劑盒 SMARTerTM RACE cDNA Amplification Kit 進行,其中第1輪和第2輪擴增所用的基因特異性引物見表1。以獲得的RACE序列與已知的Unigene基因序列拼接的序列設計全長cDNA擴增引物(表 1),利用RT-PCR技術進行基因全長cDNA擴增。cDNA模板制備:1 μg膠乳RNA,20 μL反應體系,逆轉錄合成cDNA第一鏈,稀釋5倍后用作PCR擴增。全長cDNA擴增體系:Pyrobest DNA Polymerase(5 units/μL)0.25 μL,10×Pyrobest Buffer II 5 μL,dNTP Mixture(2.5 mmol/L each)4 μL,FL-CDNA-MAPK1-F(10 μmol/L)2.5 μL,FL-CDNA-MAPK1-R(10 μmol/L)2.5 μL,cDNA模版1 μL,滅菌水補足 50 μL。擴增程序為:95 ℃預變性3 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 2 min,共30個循環(huán);72 ℃延伸10 min。0.8%的瓊脂糖凝膠電泳分離PCR擴增產物,回收純化目的條帶,克隆到pEASY-Blunt Simple Cloning Vector載體上,送Invitrogen公司進行測序。利用BLAST檢索GenBank數據庫進行同源性分析。
1.2.4 生物信息學分析 利用DNAMAN軟件對HbMAPK1全長cDNA序列進行比對分析,利用NCBI網站ORF Finder軟件進行開放閱讀框(ORF)預測,同時翻譯成蛋白序列。從NCBI 數據庫下載不同植物的MAPK蛋白序列,首先用Clustal W進行序列多重比對,再利用MEGA 4.0 軟件,選擇neighbour -joining(NJ)模型,并進行1 000次bootstrap統(tǒng)計學檢驗,構建包括HbMAPK1蛋白序列在內的植物MAPK蛋白的系統(tǒng)進化樹。采用ProtParam軟件分析該蛋白的分子量與等電點;SOPMA SECONDARY STRUCTURE PREDICTION
METHOD在線軟件預測蛋白的二級結構;TargetP 1.1 Server在線軟件進行導肽的預測;PSORT在線軟件對該蛋白進行亞細胞定位預測;ProtScale在線軟件預測該氨基酸序列的疏水性/親水性;NetPhos 2.0 Server在線軟件分析該蛋白的磷酸化位點。
1.2.5 熒光實時定量PCR分析 采用Bio-Rad公司的CFX實時熒光定量PCR系統(tǒng),實驗操作按儀器使用說明書進行。取1 μg膠乳RNA,逆轉錄合成cDNA第一鏈,稀釋10倍后作為實時定量PCR分析模板。20 μL反應體系中,包含2 μL模板,10 μL 2×SYBR Premix和10 μmol/L的每條引物0.6 μL(引物終濃度為0.3 μmol/L)。PCR反應程序為:95 ℃變性30 s;95 ℃ 10 s,60 ℃ 20 s,72 ℃ 20 s,共40個循環(huán)。40個循環(huán)后進行溶解曲線分析。利用CFX manager 3.0軟件自動進行基線和Cq值分析,以HbActin作為內參基因,算法為2-△△Cq法。
1.3 數據處理
采用T-test方法對處理與對照的基因表達水平進行差異顯著性分析(p<0.05為達到顯著性差異)。
2 結果與分析
2.1 HbMAPK1全長cDNA的克隆
通過RACE方法及測序從巴西橡膠樹膠乳中分離得到499 bp的5′端序列和334 bp的3′端序列。經拼接和RT-PCR擴增測序獲得HbMAPK1全長cDNA 1 909 bp,其中包括1 119 bp的開放閱讀框,5′非翻譯區(qū)長481 bp,3′非翻譯區(qū)長309 bp,該基因編碼372個氨基酸(圖1)。
2.2 HbMAPK1生物信息學分析
按照1.2.4所描述的方法對HbMAPK1進行生物信息學分析。結果顯示:HbMAPK1編碼一個由372個氨基酸殘基組成的蛋白,分子式為C1913H3004N520O540S20,帶負電荷的氨基酸殘基數(Asp+Glu)為42,帶正電荷的氨基酸殘基數(Arg+Lys)為42。該蛋白分子量為42.57 ku,理論等電點為7.21,不穩(wěn)定系數為37.20,屬于穩(wěn)定類蛋白。該氨基酸序列的二級結構由42.47%的α-螺旋、17.20%的延伸鏈、6.45%的β-轉角和33.87%的無規(guī)則卷曲組成。α-螺旋和無規(guī)則卷曲是其主要的結構元件,而β-轉角和延伸鏈則散布于整個蛋白質中。該蛋白可能不存在導肽酶切位點,不具有導肽,預測可信度為2級。該蛋白定位于細胞質的可能性為45%,定位于微體(過氧化物酶體)的可能性為30%,定位于葉綠體基質及葉綠體類囊體膜的可能性均為20%。結合跨膜區(qū)分析,結果顯示該蛋白不具有跨膜區(qū)。因此,推測該蛋白可能是細胞質蛋白。
疏水性/親水性分析結果顯示,多肽鏈第53位的Glu具有最低的分值(-2.189),親水性最強;第224位的Phe具有最高的分值(2.167),疏水性最強。但從整體來看,親水性氨基酸均勻分布在整個肽鏈中,且存在一個明顯的親水區(qū)域。因此,該蛋白整條多肽鏈表現為親水性。
另外,MAPK作為MAPK級聯(lián)途徑MAPKKK →MAPKK→MAPK中的末位信號轉導,磷酸化位點預測和分析對于了解該蛋白在細胞信號傳遞過程中具有重要的作用。結果顯示,該蛋白共有18個磷酸化位點,其中包括5個絲氨酸位點、8個蘇氨酸位點、5個酪氨酸位點。
2.3 氨基酸序列比對及進化樹構建
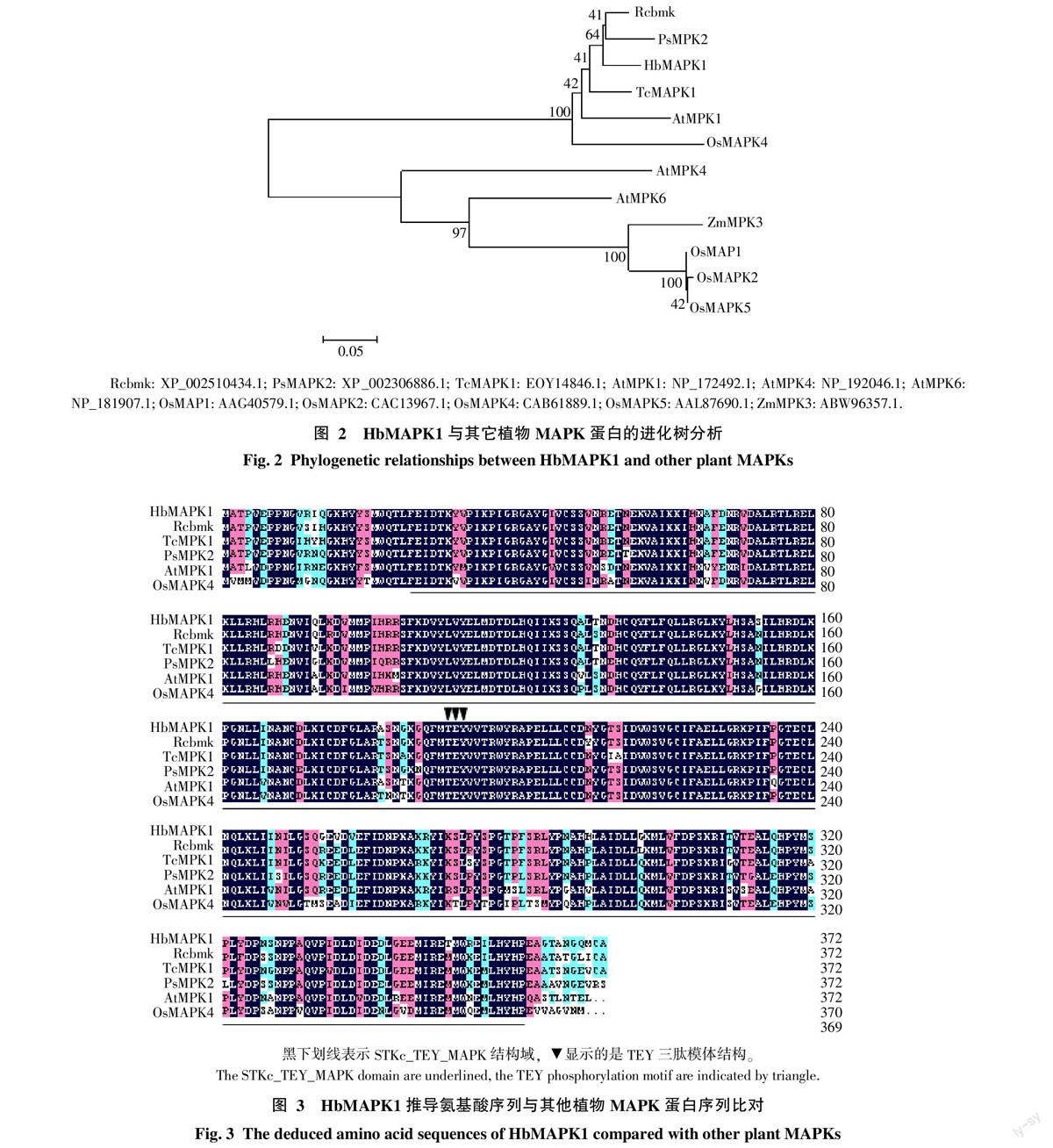
根據HbMAPK1序列推導的氨基酸序列與其他植物同源基因的氨基酸序列構建系統(tǒng)進化樹,聚類結果顯示,橡膠樹HbMAPK1與蓖麻(Ricinus communis)(XP_002510434.1)的親緣關系最近(圖2)。
HbMAPK1推導的氨基酸序列與其他植物的MAPK基因所編碼的氨基酸序列進行比對,結果顯示,該蛋白序列與蓖麻Rcbmk(XP_002510434.1)、可可(Theobroma cacao)TcMAPK1(EOY14846.1)、毛果楊(Populus trichocarpa)PsMAPK2(XP_002306886.1)、
擬南芥(Arabidopsis thaliana)AtMPK1(NP_172492.1)及水稻(Oryza sativa)OsMAPK4(CAB61889.1)蛋白相似性分別為93.82%、91.13%、90.86%、86.02%和83.06%(圖3),均具有STKc-TEY-MAPK結構域和TEY三肽模體機構。
2.4 HbMAPK1基因表達與分析
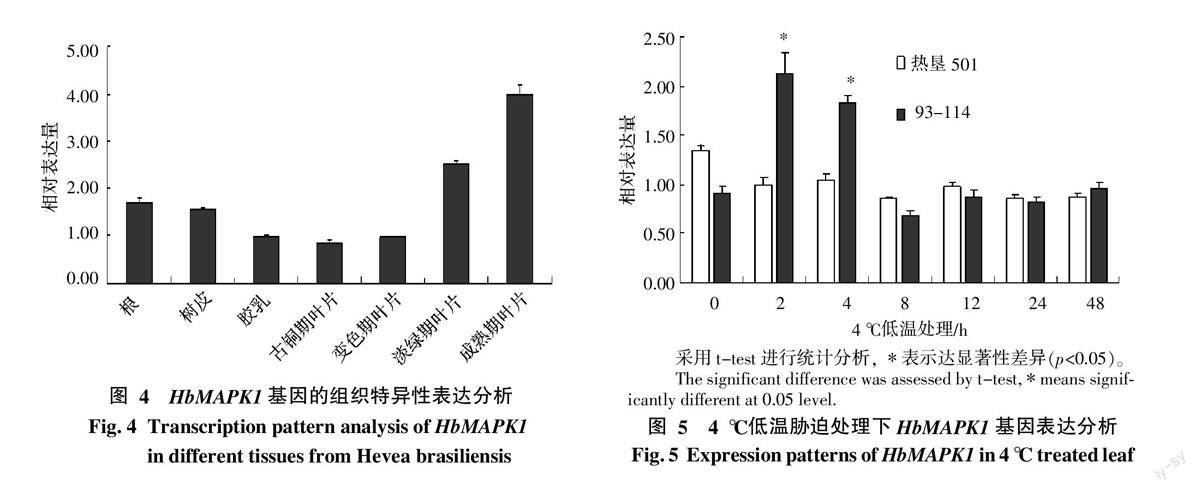
HbMAPK1基因的組織特異性相對表達分析結果顯示,HbMAPK1在橡膠樹的根、樹皮、膠乳及葉片中均有表達,其中膠乳中的表達量最低,成熟期葉片中表達最高,不同時期葉片HbMAPK1的表達量也有所不同,從古銅期至成熟期表達量逐漸升高(圖4)。4 ℃ 低溫處理條件下,HbMAPK1在93-114與熱墾501無性系間的基因表達模式存在明顯差異。常溫條件下,熱墾501葉片中HbMAPK1本底表達量比93-114中的表達量高。但是,4 ℃低溫處理2 h,橡膠樹抗寒品系93-114的樹葉中HbMAPK1的表達顯著上調,達到2倍以上,并能持續(xù)至4、8 h后基因表達開始回落至本底水平,并穩(wěn)定至48 h。而熱墾501在4 ℃低溫處理2 h后,HbMAPK1的基因表達開始下調,8 h達到最低,而且在2~48 h范圍內的表達均處于下調狀態(tài)(圖5)。由此可見,橡膠樹無性系93-114葉片中HbMAPK1的表達受到4 ℃低溫的誘導顯著上調,無性系熱墾501葉片中的表達下調。根據2個品系在生產上的抗寒表現,即無性系93-114屬于抗寒性品系,抵御低溫能力較強,而熱墾501屬于不抗寒性品系,對低溫較敏感,因此推測HbMAPK1可能在橡膠樹抵御低溫脅迫中起到一定的作用。
3 討論與結論
橡膠樹HbCBF1[11]、HbCBF2[12]、HbMT2a[13]、HbSoloit[14]等基因受低溫誘導表達。安澤偉等[15]采用cDNA-AFLP技術對11個橡膠樹抗寒無性系和12個不抗寒無性系低溫處理前后基因表達譜進行差異分析,從中篩選12條在2組材料中表現一致的差異片段,其中TDF7-8、TDF5-15、TDF16-10片段分別與植物抗逆相關的ABC轉運蛋白、3-脫氧-D-阿拉伯庚酮糖-7-磷酸合成酶(DAHPS)、WRKY基因具有較高的相似性。Silva等[16]在橡膠樹低溫誘導的文庫中發(fā)現ATP合酶CF0 C亞基(CF0)、NAD(P)H醌氧化還原酶 H亞基(NADH)、光系統(tǒng)II10 ku多肽(PsbR)及吲哚-3-乙酸誘導蛋白ARG2-1和ARG2-2的表達受低溫誘導,并通過qRT-PCR的驗證。植物中MAPK廣泛參與多種生物與非生物脅迫,如病害、低溫、干旱、鹽害和紫外線等引起的信號傳導過程[17-18]。研究結果表明,煙草(NtNPK1)、擬南芥(AtMAP3、AtMPK4、AtMPK6)、水稻[OsMAP1、OsMAPK2、OsMAPK4、OsMAPK5、OsMAPK8(OsWJUMK1)]、玉米(ZmMPK3)等MAPK級聯(lián)組分與低溫脅迫相關[19-20]。
目前,橡膠樹低溫脅迫相關MAPK基因的鑒定及機理的研究還未見報道。本研究鑒定了橡膠樹MAPK級聯(lián)途徑組分基因HbMAPK1。其他植物MAPK1的研究結果表明,擬南芥AtMAPK1激酶可被機械損傷、ABA、JA、H2O2等脅迫信號所激活[21]。玉米根系中克隆的ZmOSMAPK1的轉基因擬南芥對干旱和鹽脅迫、轉基因煙草對低溫均具有較高的耐性[22]。由于植物對寒害、干旱和鹽脅迫有通用的路徑,本研究重點關注橡膠樹抗寒性,因此,采用熒光定量PCR探討了HbMAPK1在低溫處理下橡膠樹葉片中HbMAPK1的基因表達情況。結果表明,抗寒無性系93-114中HbMAPK1的轉錄水平對4 ℃低溫處理響應較快,在2 h即達到最高,這與水稻OsMAP1和擬南芥中AtMPK4、AtMPK6對低溫的響應相似[23-26]。水稻7 d幼苗的根組織中OsMAP1的表達在12 ℃低溫處理2 h達到最高,嫩枝中OsMAP1的表達在處理6 h達到最高[23]。擬南芥中AtMPK4和AtMPK6的響應更快,其中AtMPK4激酶活性在低溫處理1 h后達到最高,AtMPK6在處理10 min內活性達到最高[24-26],更進一步說明擬南芥可能是以MEKK1-MKK2-MPK4/MPK6途徑在響應低溫脅迫的過程中起作用的[25, 27]。此外,本研究還探討了HbMAPK1在低溫較敏感的無性系熱墾501的表達情況,結果發(fā)現HbMAPK1在低溫敏感無性系熱墾501葉片中的表達模式與抗寒無性系93-114存在明顯不同,表現為受低溫誘導表達下調。
因此,參照擬南芥、水稻及玉米MAPK途徑能夠對低溫逆境作出響應,并根據橡膠樹HbMAPK1在低溫下的表達模式,推測MAPK途徑可能參與橡膠樹的低溫響應。在后續(xù)的研究中,將著重分析不同逆境條件下HbMAPK1激酶的活性變化,鑒定與HbMAPK1互作的蛋白,為建立橡膠樹逆境響應的MAPK信號途徑及開發(fā)抗寒性分子標記輔助橡膠樹無性系的鑒定奠定基礎。
參考文獻
[1] Colcombet J, and Hirt H. Arabidopsis MAPKs: a complex signalling network involved in multiple biological processes[J]. Biochem J, 2008, 413(2): 217-226.
[2] Fiil B K, Petersen K, Petersen M, et al. Gene regulation by MAP kinase cascades[J]. Curr Opin Plant Biol, 2009, 12(5):615-621.
[3] Pitzschke A, Schikora A, Hirt H. MAPK cascade signalling networks in plant defence[J]. Curr Opin Plant Biol,2009,12(4): 421-426.
[4] Sinha A K, Jaggi M, Raghuram B, et al. Mitogen-activated protein kinase signaling in plants under abiotic stress[J]. Plant Signal Behav, 2011, 6(2): 196-203.
[5] Rodriguez M C, Petersen M, Mundy J. Mitogen-activated protein kinase signaling in plants[J]. Annu Rev Plant Biol,2010, 61(4): 621-649.
[6] MAPK Group. Mitogen-activated protein kinase cascades in plants: a new nomenclature[J]. Trends Plant Sci, 2002, 7(7):301-308.
[7] Chang L, Karin M. Mammalian MAP kinase signalling cascades[J]. Nature, 2001, 410(6 824): 37-40.
[8] Gustin M C, Albertyn J, Alexander M, et al. MAP kinase pathways in the yeast Saccharomyces cerevisiae[J]. Microbiol Mol Biol Rev, 1998, 62(4): 1 264-1 300.
[9] 朱 斌, 梁 穎. 植物MAPK C族基因的研究進展[J]. 生物技術通報, 2012, 11(1): 27-31.
[10] 黃華孫. 中國橡膠樹育種五十年[M]. 北京:中國農業(yè)出版社,2005: 82-86.
[11] 程 漢, 安澤偉, 黃華孫. 巴西橡膠樹CBF1基因的克隆和序列分析[J]. 熱帶作物學報, 2005, 26(1): 50-55.
[12] 蔡海濱, 胡彥師, 黃華孫, 等. 巴西橡膠樹CBF2基因的克隆和表達分析[J]. 熱帶農業(yè)科技, 2008, 32(3): 1-5, 12.
[13] 李 言, 楊署光, 陳月異,等. 滲透脅迫與植物激素等對巴西橡膠樹HbMT2a基因表達的影響[J]. 熱帶作物學報, 2011, 32(5): 828-832.
[14] 杜 磊, 張德春, 鄧 治,等. 巴西橡膠樹一個AP2/EREBP轉錄因子的克隆及表達分析[J]. 植物生理學報, 2013, 63(11): 1 238-1 244.
[15] 安澤偉, 陳根輝,程 漢,等. 橡膠樹冷應答轉錄組cDNA-AFLP分析[J]. 林業(yè)科學, 2010, 56(3): 62-67.
[16] Silva C C, Mantello C C, Campos T, et al. Leaf-, panel- and latex-expressed sequenced tags from the rubber tree under cold-stressed and suboptimal growing conditions: the development of gene-targeted functional markers for stress response[J]. Mol Breed, 2014, 34(3): 1 035-1 053.
[17] Danquah A, de Zelicourt A, Colcombet J, et al. The role of ABA and MAPK signaling pathways in plant abiotic stress responses[J]. Biotechnol Adv, 2014, 32(1): 40-52.
[18] Meng X, Zhang S. MAPK cascades in plant disease resistance signaling[J]. Annu Rev Phytopathol, 2013, 51(2): 245-266.
[19] Samajova O, Plihal O, Al-Yousif M, et al. Improvement of stress tolerance in plants by genetic manipulation of mitogen-activated protein kinases[J]. Biotechnol Adv, 2013, 31(1):118-128.
[20] Mizoguchi T, Irie K, Hirayama T, et al. A gene encoding a mitogen-activated protein kinase kinase kinase is induced simultaneously with genes for a mitogen-activated protein kinase and an S6 ribosomal protein kinase by touch,cold,and water stress in Arabidopsis thaliana[J]. Proc Natl Acad Sci U S A, 1996, 93(2): 765-769.
[21] Ortiz-Masia D, Perez-Amador M A, Carbonell J, et al. iverse stress signals activate the C1 subgroup MAP kinases of Arabidopsis[J]. FEBS Lett, 2007, 81(9): 1 834-1 840.
[22] 吳 濤. 玉米根系促分裂源活化蛋白激酶的表達特性、轉基因植株分析及cDNA文庫的構建[D]. 泰安: 山東農業(yè)大學, 2007.
[23] Wen J Q, Oono K, Imai R. Two novel mitogen-activated protein signaling components, OsMEK1 and OsMAP1, are involved in a moderate low-temperature signaling pathway in rice[J]. Plant Physiol, 2002, 129(4): 1 880-1 891.
[24] Ichimura K, Mizoguchi T, Yoshida R, et al. Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6[J]. Plant J, 2000, 24(5): 655-665.
[25] Teige M, Scheikl E, Eulgem T, et al. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis[J]. Mol Cell, 2004, 15(1): 141-152.
[26] Smekalova V, Doskocilova A, Komis G, et al. Crosstalk between secondary messengers,hormones and MAPK modules during abiotic stress signalling in plants[J]. Biotechnol Adv,2014, 32(1): 2-11.
[27] Pitzschke A, Djamei A, Bitton F, et al. A major role of the MEKK1-MKK1/2-MPK4 pathway in ROS signalling[J]. Mol Plant, 2009, 2(1): 120-137.
