兩個煙草赤星病抗源的遺傳分析
2015-04-18 08:43:15蔣彩虹程立銳楊愛國鄭吉云趙清海楊修峰尹華玲馮全福
中國煙草科學 2015年5期
馮 瑩,蔣彩虹,程立銳,楊愛國,鄭吉云,趙清海,楊修峰,尹華玲,馮全福*
兩個煙草赤星病抗源的遺傳分析
馮 瑩1,蔣彩虹1,程立銳1,楊愛國1,鄭吉云1,趙清海2,楊修峰2,尹華玲3,馮全福1*
(1.煙草行業煙草基因資源利用重點實驗室,中國農業科學院煙草研究所,青島 266101;2.山東濰坊煙草有限公司諸城分公司,山東 諸城 262200;3.云南省煙草公司楚雄州公司雙柏縣分公司,云南 雙柏 675100)
以高抗赤星病煙草品種凈葉黃(JYH)、Beinhart1000-1(Beinhart)和感病品種NC82為材料分別構建了2個雜交組合的P1、P2、F1、F2四世代群體,成熟期赤星病菌人工接種鑒定后,采用主基因+多基因混合遺傳模型對JYH和Beinhart兩個材料進行抗性分析,結果表明,兩者的赤星病抗性均受兩對加性-完全顯性主基因+加性-顯性多基因控制。組合1的加性效應以第1對主基因為主,且多基因的加性效應大于顯性效應;組合2的兩對主基因負向加性效應相等,且多基因的顯性效應大于加性效應;2個組合F2群體主基因遺傳率分別為64.72%和63.88%,表明赤星病的抗性遺傳以主基因效應為主,并且受環境影響較大。
煙草;赤星病;主基因+多基因;遺傳分析
煙草是我國重要的葉用經濟作物,整個生育期易受多種真菌類病害侵染[1]。由交鏈孢菌引起的赤星病[A lternarialongipes(Ellet Ev.)Tisdaleet Wadk]作為一種煙草成熟期真菌病害,在各煙區普遍發生,直接影響煙葉的產量和品質[2],造成嚴重經濟損失[3-4]。考慮到降低農藥殘留及保護環境,培育煙草赤星病抗病品種是最為經濟有效的方法[5]。國內外專家對此開展了一系列研究[6]。Chaplin等[7]認為雪茄煙品種Beinhart的赤星病抗性是由單基因控制的部分顯性遺傳。從河南地方品種長脖黃選育的煙草品種凈葉黃(JYH)高抗赤星病[8]。王素琴等[9]用JYH與12個感病品種雜交后得到的多數F2群體病情指數呈現1∶2∶1的質量遺傳,且抗病性由顯性基因控制。
蓋鈞鎰等[10]研究的主基因+多基因混合遺傳模型可以準確估算基因效應和方差等遺傳參數。該模型已在大豆豆腐和豆乳得率[11]、黃瓜抗黑星病[12]、菊花花器性狀[13]、玉米株高[14,15]、小麥[16]、青花菜花球莢葉性狀[17]、煙草部分烘烤及農藝性狀[18-24]等方面得到應用,但是在煙草抗赤星病多抗源遺傳分析上尚未可見。已報道的煙草赤星病抗性遺傳結果均采用傳統分析方法,本試驗利用2份抗病材料(JYH、Beinhart)和1份感病材料(NC82)分別構建的P1、P2、F1、F2四世代分離群體,運用主基因+多基因模型分析煙草抗赤星病的遺傳規律,為煙草抗赤星病育種創新及抗病等研究提供理論依據和實踐經驗。
1 材料與方法
1.1 試驗材料
2份抗病材料為國內烤煙品種JYH和美國雪茄煙品種Beinhart;1份感病材料為烤煙品種NC82。種子均由國家煙草中期庫提供。利用上述3份材料配制兩個雜交組合。組合1以JYH(P1)為母本,NC82(P2)為父本,構建F1、F2群體;組合2以Beinhart(P1)為母本,NC82(P2)為父本,構建F1、F2群體。兩個組合、4世代試驗材料(表1)播種于中國農業科學院煙草研究所青島試驗基地。

表1 兩組合各世代材料數Table 1 The Number of each generation of two combinations
1.2 試驗方法
1.2.1 成熟期赤星病人工接種鑒定 參照蔣彩虹的[25]方法完成菌液制備、接種、病情劃分。
1.2.2 遺傳分析 采用混合遺傳模型多世代聯合分離分析方法[26]進行數據遺傳分析,建立A-E 5類共24種遺傳模型。通過最大似然法和IECM(Iterated expectation and conditional maximization)估計各世代、各成分分布的參數,然后通過AIC(Akaike's information criterion)值最小原則選擇最佳模型,同時進行一系列適合性檢驗,包括均勻性檢驗,Smirnov(nW2)和Kolmogorov(Dn),選擇最優遺傳模型。從而估計各基因效應值、各組合的遺傳效應、誤差方差σ2,成分分布方差f群體表型方差并計算主基因和多基因的遺傳方差和遺傳率
2 結 果
2.1 兩組合4世代赤星病病級的次數分布
分別對組合1和組合2的四世代群體進行成熟期人工接種赤星病菌,調查各世代單株病級。兩組合4世代赤星病抗性病級次數見圖1和圖2。兩組合抗性分離表現及卡方檢驗見表2。
對兩個抗源與NC82進行赤星病抗性顯著性檢測,結果顯示JYH與Beinhart表現抗病,與感病親本NC82差異極顯著(P<0.01)。JYH的單株病級多集中在1級,表現為高抗赤星病;Beinhart的單株病級0、1、2級均有分布,同樣也表現為高抗。組合1的F1代的平均病級為2.60,傾向于感病;組合1的F2群體呈現單峰較明顯的偏態分布,赤星病的抗性遺傳表現出主基因特征,初步推測受主基因加多基因共同控制;但卡方檢測結果表明,雜交F2代抗病單株與感病單株的分離比符合兩對互補基因理論分離比(9∶7)(表2),因此,需要利用混合遺傳模型對其抗性遺傳進一步分析和驗證。組合2的F1代的平均抗病級別為2.20,傾向于抗病。F2群體單株病級在低值和高值呈現雙峰分布,赤星病的抗性表現出主基因遺傳特征。F2代抗病單株與感病單株的比例約為0.97∶1(表2),卡方檢測結果表明該分離比均不符合一對基因(3∶1)、兩對互補基因(9∶7)或兩對顯性主基因(15∶1)的理論值,因此組合2的抗性遺傳受主基因和多基因共同控制。
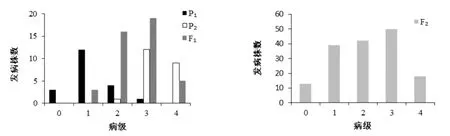
圖1 JYH×NC82組合的四世代抗性級值次數分布圖Fig. 1 Distribution of the number of plants w ith different resistance grades in 4 populations of combination YJH×NC82

表2 兩組合抗性分離表現及卡方檢驗Table 2 The χ2test of genetic resistance of two combinations
2.2 主基因+多基因遺傳分析
用“主基因+多基因”混合遺傳模型對2個組合四個世代的赤星病級數據進行遺傳分析,通過IECM算法獲得A、B、C、D、E5類共24種遺傳模型的極大似然函數值(Max-likelihood-value)和AIC值(akaike’s information criterion, AIC)。
2.2.1 JYH(組合1)抗赤星病遺傳模型 選擇AIC值較小的E-2、E-5、D-3、D-4模型為備選模型(表3)。適合性檢驗表明(表4),在20個檢驗統計量中,選擇AIC值最小且統計量達到顯著水平個數最少的E-5模型為JYH抗赤星病最優遺傳模型,即兩對加性-完全顯性主基因+加性-顯性多基因模型。
2.2.2 Beinhart(組合2)抗赤星病遺傳模型 選擇AIC值較小的E-5、E-1、D-0模型為備選模型(表3)。適合性檢驗表明(表5),在20個檢驗統計量中,選擇AIC值最小且統計量達到顯著水平個數最少的E-5模型為Beinhart抗赤星病最優遺傳模型,即兩對加性-完全顯性主基因+加性-顯性多基因模型。

表3 兩組合遺傳模型的AIC值Table 3 The AIC values of the different genetic models in two combinations
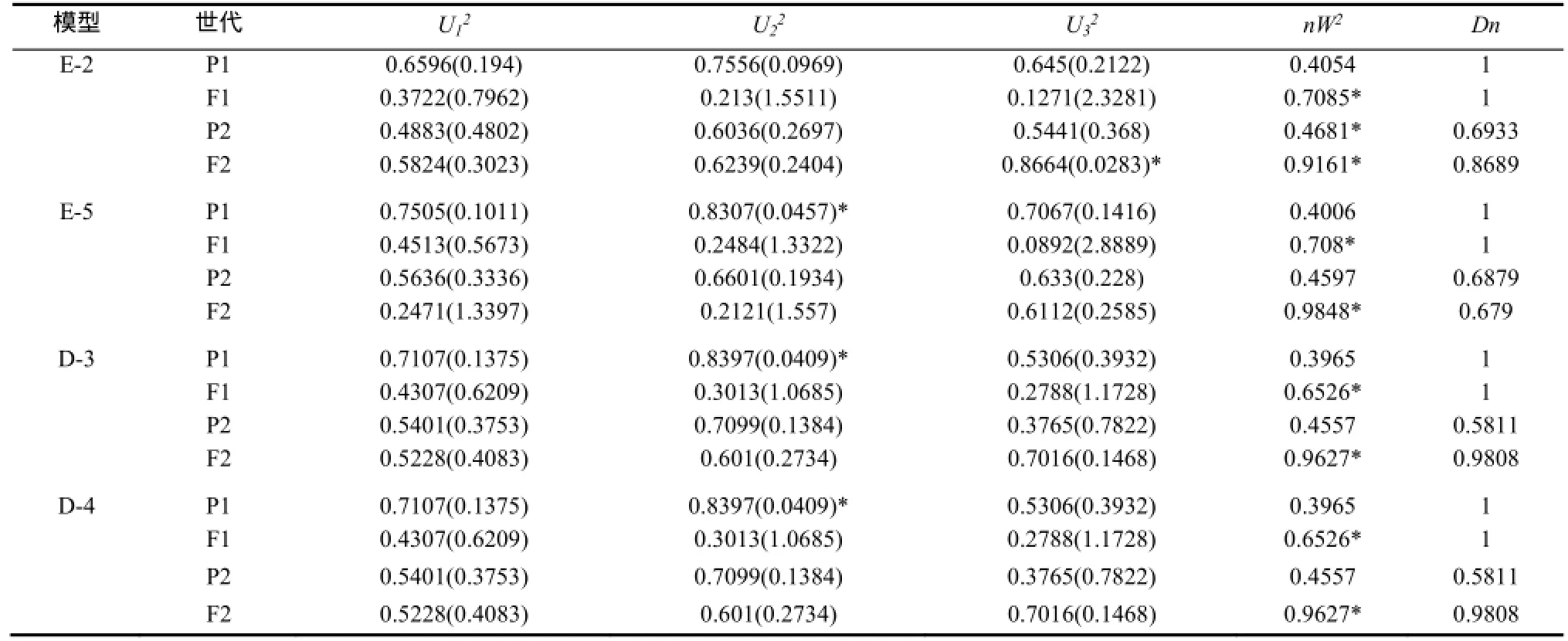
表4 YJH×NC82遺傳模型的適合性檢驗Table 4 Test for goodness of fit about YJH×NC82 genetic model
2.3 遺傳效應分析
兩組合的赤星病抗性均受2對加性-完全顯性主基因+加性-顯性多基因控制。組合1的加性效應以第1對主基因為主,[h]/[d]<1,說明多基因的加性效應大于顯性效應。組合2兩對主基因的加性效應均為-0.8935,[h]/[d]>1,說明多基因的顯性效應大于加性效應。兩組合F2群體主基因遺傳率分別為64.72%和63.88%,說明該組合赤星病的抗性遺傳以主基因效應為主,并且受環境影響較大(表6)。
3 討 論
煙草屬于自花授粉作物,長期的人工選擇過程中產生嚴重的遺傳瓶頸,導致現有品種之間遺傳背景狹窄。在煙草抗赤星病育種過程中主要使用的抗源為JYH,長期使用單一抗源會導致其抗病性下降,存在抗性品種喪失抗性的風險,因此充分利用新赤星病抗源具有重要意義。在本試驗還對赤星病的另一個重要抗源Beinhart進行了赤星病抗性遺傳研究,了解兩個抗源的遺傳規律,可以為煙草赤星病抗性育種提供理論基礎。
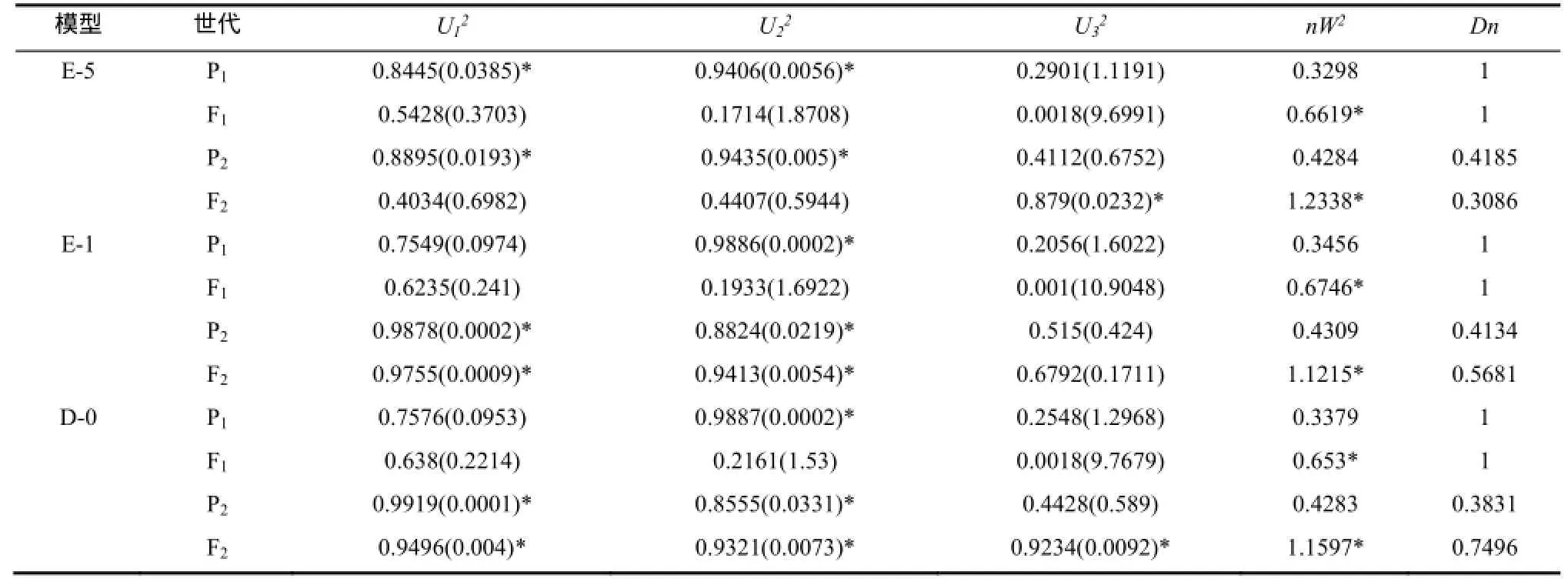
表5 Beinhart×NC82遺傳模型的適合性檢驗Table 5 Test for goodness of fit about Beinhart×NC82genetic model

表6 兩組合遺傳參數估計Table 6 The estimates of genetic parameters in tw o combinations
此前,國內外學者對這2種抗源抗病遺傳分析所得的結論不盡相同。Stavely等[27]對包括Beinhart在內的8個高抗赤星病材料做了遺傳分析,認為其抗病性都是由多基因控制的,表現為數量遺傳。Monga等[28]對10個印度黃花煙品系抗性遺傳做的雙列雜交分析,表明其抗病性由多個微效基因控制,控制抗病性的基因大部分為顯性。郭永峰等[29-30]通過研究表明抗源Beinhart的抗性位點表現為受多基因控制的水平抗性遺傳規律,而JYH抗性基因大部分是隱性的。蔣彩虹等[31]以JYH和NC82為親本,構建的F1、F2、BC1群體進行遺傳分析發現其赤星病抗性由顯性多基因控制。
本研究對上述2份高抗材料的遺傳分析顯示,兩者的抗病性均受兩對加性-完全顯性主基因+加性-顯性多基因控制,且以主基因效應為主,多基因的加顯效應有不同表現。且能夠充分利用每個單株信息,確定主基因個數及其效應,與傳統孟德爾遺傳分離比率方法相比(卡方分析),克服了孟德爾方法難以確定分組標準的難題,而且也可以直接估算主基因與多基因的遺傳效應值[12]。另外試驗利用非分離世代的表型可以估算環境誤差,在表6得出兩個組合的σ在數值上分別為0.4608、0.4608和0.5155、0.5155,根據前文1.2.2得出多基因方差及其遺傳率為0,所以多基因方差及效應值為0是由于環境對表型影響較大[23,32],導致利用非分離世代(P1、P2和F1)誤差方差較大,從而導致多基因的效應值被低估,這是試驗方法及數量性狀試驗誤差本身造成的[33]。由于F2群體無法對試驗結果重復驗證,針對出現的問題在以后的試驗中可以考慮利用DH或RIL等重復性較好的群體對赤星病抗性等多基因遺傳率較低的數量性狀進行分離分析,從而提高遺傳分析的精度。
本研究采用主基因+多基因混合遺傳模型,不需要分子實驗室條件,僅依據表型數據分析即可確定赤星病抗性遺傳模型。而現在發展相對成熟的QTL分子標記技術需要適當的分離群體和準確的試驗數據,這些試驗數據也可用來進行遺傳分析,兩種方法所得結果可以相互印證。因而,分離分析法可以單獨使用,也可以作為QTL定位的前期研究基礎,在開展主效QTL定位之前,利用分離分析法優先分析其遺傳規律,對發掘主效QTL具有重要參考價值。本課題組分別對JYH和Beinhart這兩個赤星病抗源進行了QTL定位。結果表明,對于抗源Beinhart,高婷婷等[34]發掘到2個與赤星病抗性相關的主效QTL,分別位于7號和15號連鎖群上,抗性位點來源于抗源Beinhart,這與前面所得結論是相互印證的。蔣彩虹[31]以赤星病抗源JYH為材料獲得1個與抗性基因緊密連鎖的QTL標記,位于23號連鎖群。本研究表明,雖然赤星病抗源JYH的抗性受2對主基因+多基因控制,但是以第一對主效基因的加性效應為主,因此與QTL定位結果基本相同。在本研究中確定的另外一個效應值相對較小的主效基因在QTL定位中沒有定位到的原因可能與我們在進行抗源JYH定位中采用的是BSA法有關。BSA法是一種行之有效的發掘主效QTL的方法,但是其缺點是會漏掉效應值較小的位點。所以,在利用BSA法進行赤星病抗性QTL定位的過程中,可能將來源于抗源JYH的另一個主效QTL遺漏。因此,根據遺傳分析結果,用抗源JYH和感病親本配制的分離分析群體,利用SSR標記分析每個后代基因型,進行全基因組QTL發掘將是接下來的一個研究重點。除此之外,根據課題組已有的研究基礎,選擇JYH為進一步挖掘和鑒定赤星病抗性基因的材料。在充分了解兩個抗源抗病機理和抗性遺傳方式的基礎上,采取常規雜交育種和分子標記相結合,完成抗性基因的轉移,加快品種繁育,更快獲得優質高抗,適合生產需要的抗病品種也是今后的一個研究重點。
4 結 論
本研究采用主基因+多基因混合遺傳模型對JYH和Beinhart兩個材料進行分析,結果表明,兩者的赤星病抗性均受兩對加性-完全顯性主基因+加性-顯性多基因控制,且以主基因效應為主,受環境影響較大。試驗依據表型即可判斷遺傳模型,可為后續的基因定位提供參考。
[1] 張嚴柱. 中國煙草行業發展戰略選擇問題研究[D].大連:東北財經大學,2012.
[2] 易龍,肖崇剛,馬冠華,等. 防治煙草赤星病有益內生細菌的篩選及抑菌作用[J]. 微生物學報,2004,44(1):19-22.
[3] Lucas G B. Disease of Tobacco[M]. 3rd ed. Biological Consulting Association, Raleigh. Harold. E. Parker and Sons, Printers. Fuquay-Varina, New Caledonia, 1975∶267-296.
[4] Yakinova E T, Yordanova Z P, Slavov S, et al. Alternaria alternata AT toxin induces programmed cell death in tobacco[J]. Phytopathol, 2009(157)∶ 592-601.
[5] 佟道儒. 煙草育種學[M]. 北京:中國農業出版社,1997:432-438.
[6] 張萬良,翟爭光,謝揚軍,等. 煙草赤星病研究進展[J]. 江西農業學報,2011,23(1):118-120.
[7] Chaplin, J F, T W. Graham. Brown spot resistance in Nicotiana tabacum[J]. Tob. Sci., 1963(7)∶ 59-62.
[8] 中國農業科學院煙草研究所. 中國煙草品種志[M].北京:農業出版社,1987.
[9] 王素琴,李楊立,劉鳳蘭,等. “凈葉黃”抗赤星病遺傳規律的測定[J]. 煙草科技,1995(1):30-32.
[10] 蓋鈞鎰. 植物數量性狀遺傳體系的分離分析方法研究[J]. 遺傳,2005,27(1):130-136.
[11] 王春娥,蓋鈞鎰,傅三雄,等. 大豆豆腐和豆乳得率的遺傳分析與QTL定位[J]. 中國農業科學,2008,41(5):1274-1282.
[12] 李全輝,沈鏑,李錫香,等. 黃瓜抗黑星病不同基因源的遺傳分析[J]. 植物遺傳資源學報,2011,12(2):291-296.
[13] 張飛,陳發棣,房偉民,等. 菊花花器性狀雜種優勢與混合遺傳分析[J]. 中國農業科學,2010,43(14):2953-2961.
[14] 王鐵固,馬娟,張懷勝,等. 玉米株高主基因+多基因遺傳模型分析[J]. 玉米科學,2012,20(4):45-49.
[15] 馬娟. 玉米主要株型性狀的主基因+多基因遺傳模型分析[D]. 新鄉:河南科技學院,2012.
[16] 侯璐,宋曉賀,路亞明,等. 小簇麥易位系V9128-3抗條銹病基因的遺傳分析和SSR分子標記[J]. 植物病理學報,2009,39(1):67-75.
[17] 劉二艷,劉玉梅,方智遠,等. 青花菜花球‘莢葉’性狀主基因+多基因遺傳分析[J]. 園藝學報,2009,36(11):1611-1618.
[18] 王日新,任民,張興偉,等. 普通煙草栽培種內株高性狀主基因加多基因遺傳分析[J]. 中國煙草科學,2009,30(2):15-20.
[19] 倪超,徐秀紅,張興偉,等. 烤煙品種易烤性相關性狀的主基因+多基因遺傳分析[J]. 中國煙草科學,2011,32(1):1-4,11.
[20] 張興偉,王志德,牟建民,等. 烤煙葉綠素含量遺傳分析[J]. 中國煙草學報,2011,17(3):48-52.
[21] 張興偉,王志德,孫玉合,等. 烤煙葉數、葉面積的遺傳分析[J]. 植物遺傳資源學報,2012,13(3):467-472.
[22] 張興偉,王志德,任民,等. 烤煙幾個重要植物學性狀的遺傳分析[J]. 中國煙草科學,2012,33(5):1-8.
[23] 牟建英,錢玉梅,張興偉,等. 煙草白粉病抗性基因的遺傳分析[J]. 植物遺傳資源學報,2013,14(4):668-672.
[24] 高加明,王志德,張興偉,等. 香料煙青枯病抗性基因的遺傳分析[J]. 中國煙草科學,2010,31(1):1-4.
[25] 蔣彩虹. 煙草赤星病抗性分子標記篩選[D]. 北京:中國農業科學院,2007.
[26] 蓋鈞鎰等. 植物數量性狀遺傳體系[M]. 北京:科學出版社,2003:22-23.
[27] Stavely J R. Inheritance of brown spot resistance in Nicotiana tabacum[J]. Proc. Am. Phytopathol., 1975(2)∶228-230.
[28] Dobhal V K, Monga D. Genetic analysis of field resistance to brown spot caused by A lternaria alternata( Fries) Keissler in Nicotiana tabacum Linn[J]. Tob. Res. 1991, 17(1)∶ 11-15.
[29] 郭永峰,付憲奎,哈君利. 抗赤星病煙草及其研究利用[J]. 中國煙草科學,1998,19(1):32-35.
[30] 郭永峰,朱賢朝,石金開,等. 煙草對赤星病田間抗性的遺傳研究[J]. 中國煙草科學,1998,19(3):1-6.
[31] 蔣彩虹,羅成剛,任民,等. 一個與凈葉黃抗赤星病基因緊密連鎖的SSR標記[J]. 中國煙草科學,2012,33(1):19-22.
[32] 沈鏑,方智遠,李錫香,等. 黃瓜果肉色的遺傳分析[J]. 植物遺傳資源學報,2011,12(2):216-222.
[33] 章元明,蓋鈞鎰. 利用DH和IRIL群體鑒定QTL體系并估計其遺傳效應[J]. 遺傳學報,2000,27(7):634-640.
[34] 高亭亭,蔣彩虹,羅成剛,等. Beinhart1000-1抗赤星病基因的QTL定位[J]. 中國煙草學報,2014,20(2):104-107.
Genetic Analysis of Resistance to Brown Spot Disease in Tobacco Cultivars Jingyehuang and Beinhart1000-1
FENG Ying1, JIANG Caihong1, CHENG Lirui1, YANG Aiguo1, ZHENG Jiyun1, ZHAO Qinghai2,YANG Xiufeng2, YIN Hualing3, FENG Quanfu1*
(1. Key Laboratory for Tobacco Gene Resources, Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 2. Zhucheng Branch of Weifang Tobacco Co., Ltd., Zhucheng, Shandong 262200, China; 3. Shuangbai Branch of Chuxiong Tobacco Company, Shuangbai, Yunnan 675100, China)
Tobacco cultivar/line Jingyehuang, named JYH, and Beinhart1000-1, named Beinhart, have shown effective resistance to brown spot disease. In this study, populations of P1, P2, F1, and F2were generated as experimental materials from two crosses between the two cultivars and susceptible cultivar NC82. The joint segregation analysis method of mixed major gene plus polygene genetic model was used to investigate the inheritance of resistance to brown spot disease in tobacco. The results showed that the inheritances of both JYH and Beinhart fitted to a m ix genetic model of two major genes w ith additive-com plete dom inance effects p lus poly-genes w ith additive-dom inance effects (E-5 model). For high resistance material JYH, the additive effect of the first major gene was larger than the second one. For high resistance material Beinhart, the additive effect of two major genes was equal. Heritability of the major genes was 64.72% in JYH×NC82, and 63.88% in Beinhart×NC82 in F2, indicating that resistances to brown spot disease in tobacco cultivars JYH and Beinhart were mainly controlled by major genes.
tobacco; brown spot disease; major gene plus poly-gene; genetic analysis
S572.03
1007-5119(2015)05-0001-07
10.13496/j.issn.1007-5119.2015.05.001
公益性行業(農業)科研專項項目(201203091);中國煙草總公司科技重大專項項目[110201301009(JY-09)]
馮 瑩(1988-),女,在讀碩士研究生,研究方向:作物遺傳育種。E-mail:fyacang@sina.com。*通信作者,E-mail:quanff@163.com
2015-04-29
2015-09-16
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
光學精密工程(2016年6期)2016-11-07 09:07:19
當代化工研究(2016年9期)2016-03-20 16:22:08
