融合蛋白TAP-SSL5對激活的血小板與人淋巴細胞結合的影響*
2015-04-17 07:48:41貝俊杰胡厚源
中國病理生理雜志 2015年1期
彭 松, 貝俊杰, 胡厚源, 陳 強
(第三軍醫大學西南醫院心血管內科,重慶 400038)
?
融合蛋白TAP-SSL5對激活的血小板與人淋巴細胞結合的影響*
彭 松, 貝俊杰, 胡厚源△, 陳 強
(第三軍醫大學西南醫院心血管內科,重慶 400038)
目的: 研究抗炎、抗凝雙效融合蛋白蜱抗凝血肽(TAP)-金黃色葡萄球菌超抗原樣蛋白5(SSL5)對激活的血小板與人淋巴細胞結合作用的影響。方法: 采用免疫磁珠分選法篩選人外周血總淋巴細胞;CCK-8法檢測TAP-SSL5對細胞活力的影響;流式細胞術檢測Jurkat細胞 (人外周血白血病T細胞株) 表面CD162 (PSGL-1)的表達及TAP-SSL5對小鼠抗人CD162單抗(KPL-1)與Jurkat細胞結合的抑制作用。以20 μmol/L ADP激活人血小板,流式細胞術檢測血小板與Jurkat細胞或人淋巴細胞的結合情況,并研究TAP-SSL5的干預作用。 結果: 30 mg/L及以下濃度的TAP-SSL5對Jurkat細胞的活力無明顯影響。流式細胞術檢測顯示,10 mg/L 的TAP-SSL5能顯著抑制KPL-1與Jurkat細胞的結合;20 μmol/L ADP激活的血小板與Jurkat細胞或淋巴細胞的結合率分別為(11.86±4.49)% 和 (8.32±1.00)%;細胞經10 mg/L TAP-SSL5預先處理后,結合率分別降至 (6.73±2.71)% 和 (5.51±0.70)% ,差異有統計學意義。結論: TAP-SSL5可與淋巴細胞表面的PSGL-1結合,從而抑制激活的血小板與人淋巴細胞的結合,這可能是抗炎、抗凝雙效融合蛋白TAP-SSL5發揮其抗炎作用的機制之一。
蜱抗凝血肽; 金黃色葡萄球菌超抗原樣蛋白5; 融合蛋白; 血小板; 淋巴細胞; P-選擇素糖蛋白配體1
動脈粥樣硬化(atherosclerosis,AS)是一種慢性炎癥性疾病,免疫調節在其發生發展過程中起到重要作用。淋巴細胞和血小板作為免疫及炎癥反應中的重要效應細胞,參與了動脈粥樣硬化病程的各個環節。血小板與淋巴細胞之間通過直接接觸和分泌可溶性介質的方式發生著相互作用而影響彼此功能,從而調控動脈粥樣硬化等炎癥性疾病的病理生理過程[1]。P-選擇素糖蛋白配體1(P-selectin glycoprotein ligand 1, PSGL-1)廣泛表達于白細胞表面,它可與激活的內皮或血小板表面的P-/E-選擇素結合,導致白細胞在內皮或血小板表面的黏附,而促進白細胞在病變血管處的募集和白細胞-血小板聚集體的形成。在前期研究中,我們已將能夠同PSGL-1結合的金黃色葡萄球菌超抗原樣蛋白5 (staphylococcal superantigen like protein-5, SSL5)[2]與凝血因子Xa (FXa)的強效抑制劑蜱抗凝血肽(tick anticoagulant peptide, TAP)聯合,成功構建了具有抗炎抗凝雙效功能的融合蛋白TAP-SSL5。并且發現TAP-SSL5可抑制粒細胞在P-選擇素表面的黏附,還可抑制激活血小板與粒細胞的結合[3-4]。血小板與淋巴細胞的結合可以在動脈血流條件下促進淋巴細胞黏附到內皮下基質[5],并影響淋巴細胞的增殖、分化等功能[6]。因此,本文進一步研究了人血小板同淋巴細胞的結合效應及融合蛋白TAP-SSL5對該結合的影響,旨在進一步探討融合蛋白TAP-SSL5在抑制炎癥方面的作用機制。
材 料 和 方 法
1 主要材料
RPMI-1640和胎牛血清購自HyClone;Ficoll-paque液購自Pharmacia;CCK-8試劑盒購自Dojindo;免疫磁珠細胞分選(MACS)試劑盒為Miltenyi Biotec產品;BSA、二磷酸腺苷(ADP)為Sigma產品; PE標記的小鼠抗人CD62P單克隆抗體、PE標記的小鼠抗人CD162(PSGL-1)單克隆抗體(KPL-1)和PE標記的小鼠IgG1(mIgG1-PE)為BD產品。融合蛋白 TAP-SSL5及重組SSL5蛋白的表達與純化采用曲小龍等[3]介紹的方法實施,融合蛋白TAP-SSL5純度≥95%,重組SSL5蛋白純度≥95% 。
2 方法
2.1 人外周血白血病T細胞株Jurkat細胞的培養 Jurkat細胞由第三軍醫大學全軍免疫學研究所贈送,采用含10%胎牛血清的RPMI-1640完全培養基,于37 ℃、5% CO2恒溫箱中培養。
2.2 Ficoll-paque密度梯度離心法分離人外周血單個核細胞 取健康成人抗凝血白膜(取自西南醫院輸血科),用PBS等比例稀釋,按2∶1的比例將白膜輕輕鋪于Ficoll-paque分離液之上,400 ×g離心30 min,吸取單個核細胞層至無菌管中,PBS洗滌細胞3遍,去除紅細胞、血小板,重懸于MACS緩沖液(含PBS、 0.5% BSA、1 mmol/L EDTA,pH 7.2)中。
2.3 免疫磁珠分選陽性總淋巴細胞 用90 μL MACS緩沖液重懸3×107個單個核細胞,加入30 μL FcR阻斷劑,加入30 μL 生物素化的抗淋巴細胞混合抗體,在4 ℃孵育10 min;加入60 μL抗生物素磁性微珠,4 ℃孵育15 min;用MACS緩沖液洗滌細胞1次,并重懸至2×1011/L;將細胞加入位于磁場中的分選柱上,洗脫未結合的細胞;將分選柱移出磁場,洗脫并收集混合抗體標記的細胞。臺盼蘭染色鑒定細胞活力,以改良Tyrode’s 緩沖液(150 mmol/L NaCl,12 mmol/L NaHCO3,2.5 mmol/L KCl,2 mmol/L CaCl2,2 mmol/L MgCl2,1 g/L D-glucose,1 g/L BSA,pH 7.4)調整細胞濃度。
2.4 洗滌血小板懸液的準備 健康成人枸櫞酸鈉抗凝的濃縮血小板(取自西南醫院輸血科),1 000 r/min離心15 min,得到富含血小板血漿(platelet-rich plasma, PRP)。 2 700 r/min,離心10 min以沉淀血小板,以CGS緩沖液(12.9 mmol/L trisodium citrate、120 mmol/L NaCl,30 mmol/L D-glucose,pH 6.5)洗滌沉淀2次,以SysmexKX-21N血細胞自動分析儀計數血小板,用改良Tyrode’s 緩沖液調整濃度至2.5×1011/L。2.5 細胞活力的檢測 取對數生長期的Jurkat細胞(1×108/L)接種于96孔板,100 μL/well, 每組設3個復孔,空白組只含培養基,不加細胞。置于37 ℃、5% CO2飽和濕度孵箱培養24 h后,干預組分別加入含TAP-SSL5或SSL5的培養基(終濃度為3、10和30 mg/L),陰性對照組以PBS代替重組蛋白,繼續培養24 h。然后加入CCK-8溶液,10 μL/well,繼續培養3 h。酶標儀(Bio-Rad)測定450 nm波長處的吸光度A450值,并計算細胞存活率。細胞存活率(%)=(實驗組A450-空白組A450)/(陰性對照組A450-空白組A450)×100%。實驗共重復3次。
2.6 Jurkat細胞表面PSGL-1的檢測及TAP-SSL5對KPL-1結合作用的影響 取適量Jurkat細胞,加入終濃度為10 mg/L的融合蛋白TAP-SSL5或SSL5,冰上孵育20 min,0.01 mol/L PBS洗滌細胞2次,再加入KPL-1-PE,冰上孵育30 min,同時以mIgG1-PE為同型抗體陰性對照,PBS洗滌細胞2次,200 μL PBS重懸細胞,流式細胞儀檢測TAP-SSL5對KPL-1在Jurkat細胞表面結合的影響。實驗共重復3次。
2.7 血小板與Jurkat細胞或淋巴細胞的結合 實驗分未激活血小板-淋巴細胞結合組(baseline組)、ADP激活血小板-淋巴細胞結合組(control組)、ADP激活血小板-經TAP-SSL5預處理的淋巴細胞結合組(1 mg /L 及10 mg/L TAP-SSL5組)、ADP激活血小板-經SSL5預處理的淋巴細胞結合組(SSL5組)。
取適量的Jurkat細胞或人淋巴細胞,調整細胞濃度至5×109/L,干預組分別加入終濃度為1 mg/L、10 mg/L 的TAP-SSL5或10 mg/L 的SSL5,冰上孵育20 min,以改良Tyrode’s緩沖液洗滌細胞2次,并調整細胞濃度為5×109/L。
取適量洗滌血小板懸液(2.5×1011/L),加入終濃度為20 μmol/L 的ADP以激活血小板,然后將其與Jurkat細胞或淋巴細胞(5×109/L)按等體積比例混合,再加入抗CD62P-PE mAb以標記血小板,并以mIgG1-PE作為同型對照,室溫下于暗處孵育20 min,1%多聚甲醛固定。200 ×g離心10 min,棄上清,用1%多聚甲醛重懸細胞后行流式細胞儀檢測。檢測時,先確認Jurkat細胞或淋巴細胞群,然后在FL2通道檢測細胞表面的血小板結合情況,進一步分析出結合有血小板的細胞占所檢測細胞總數的百分比。實驗共重復3~5次。
3 統計學處理
數據用均數±標準差(mean±SD)表示。采用GraphPad Prism 5.0軟件進行統計分析,比較重組蛋白對細胞活力的影響使用單因素方差分析,比較干預因素對血小板與細胞結合的影響使用Student’s配對t檢驗。以P<0.05為差異有統計學意義。
結 果
1 融合蛋白TAP-SSL5對Jurkat細胞活力的影響
TAP-SSL5或SSL5作用于Jurkat細胞24 h后,采用CCK-8法檢測細胞活力的結果表明,30 mg/L或以下的 TAP-SSL5對Jurkat細胞存活率無明顯影響,10 mg/L的 SSL5對Jurkat細胞存活率無明顯抑制,見圖1。
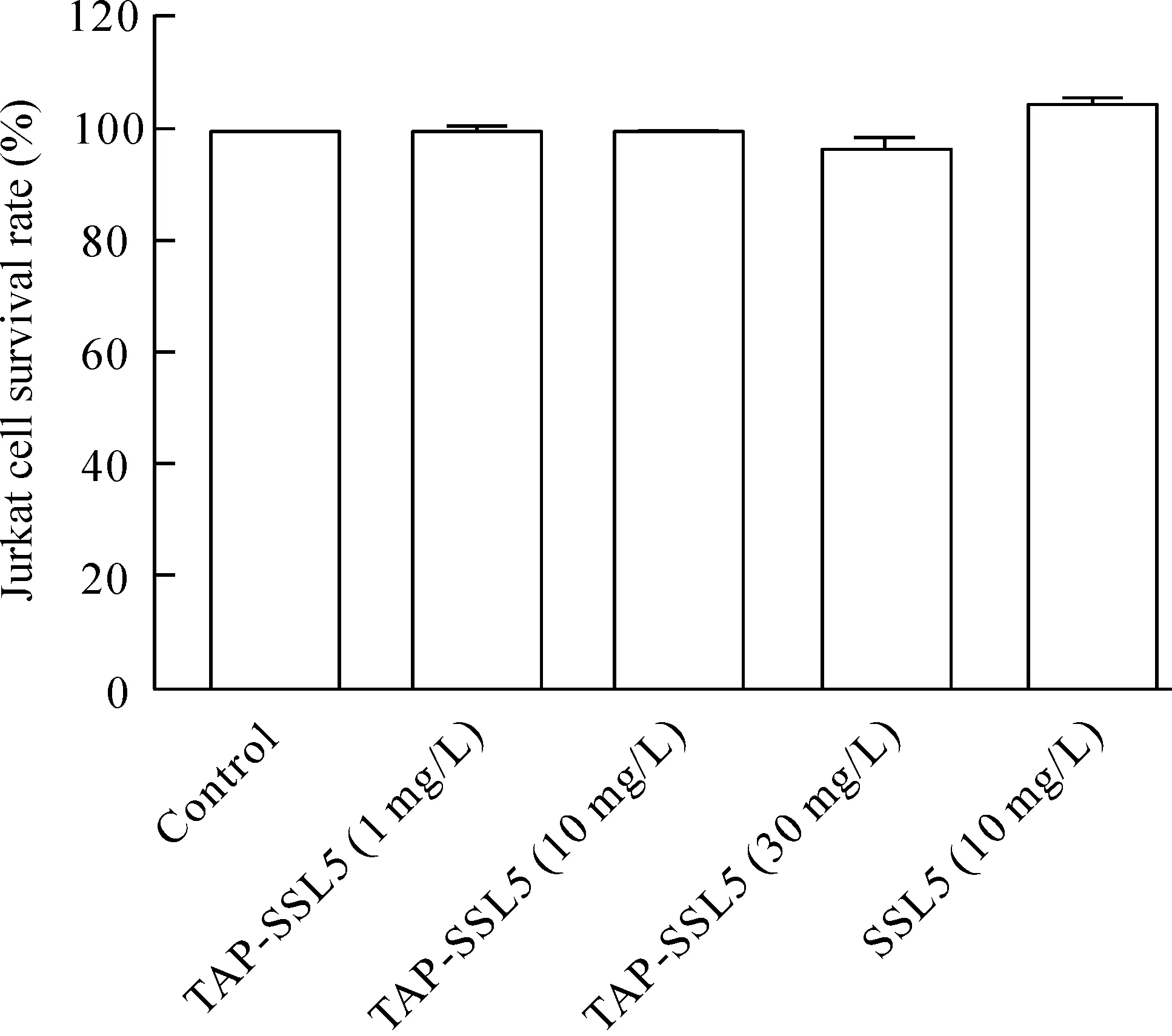
Figure 1.The effects of TAP-SSL5 and SSL5 on the viability of Jurkat cells.Mean±SD. n=3.
2 TAP-SSL5抑制KPL-1與Jurkat細胞的結合
流式細胞術檢測顯示,Jurkat細胞表面表達大量的PSGL-1(CD162),KPL-1(抗CD162 mAb)的結合陽性率為92.8%;10 mg/L 的TAP-SSL5 或SSL5能顯著抑制KPL-1與Jurkat細胞的結合,見圖2。
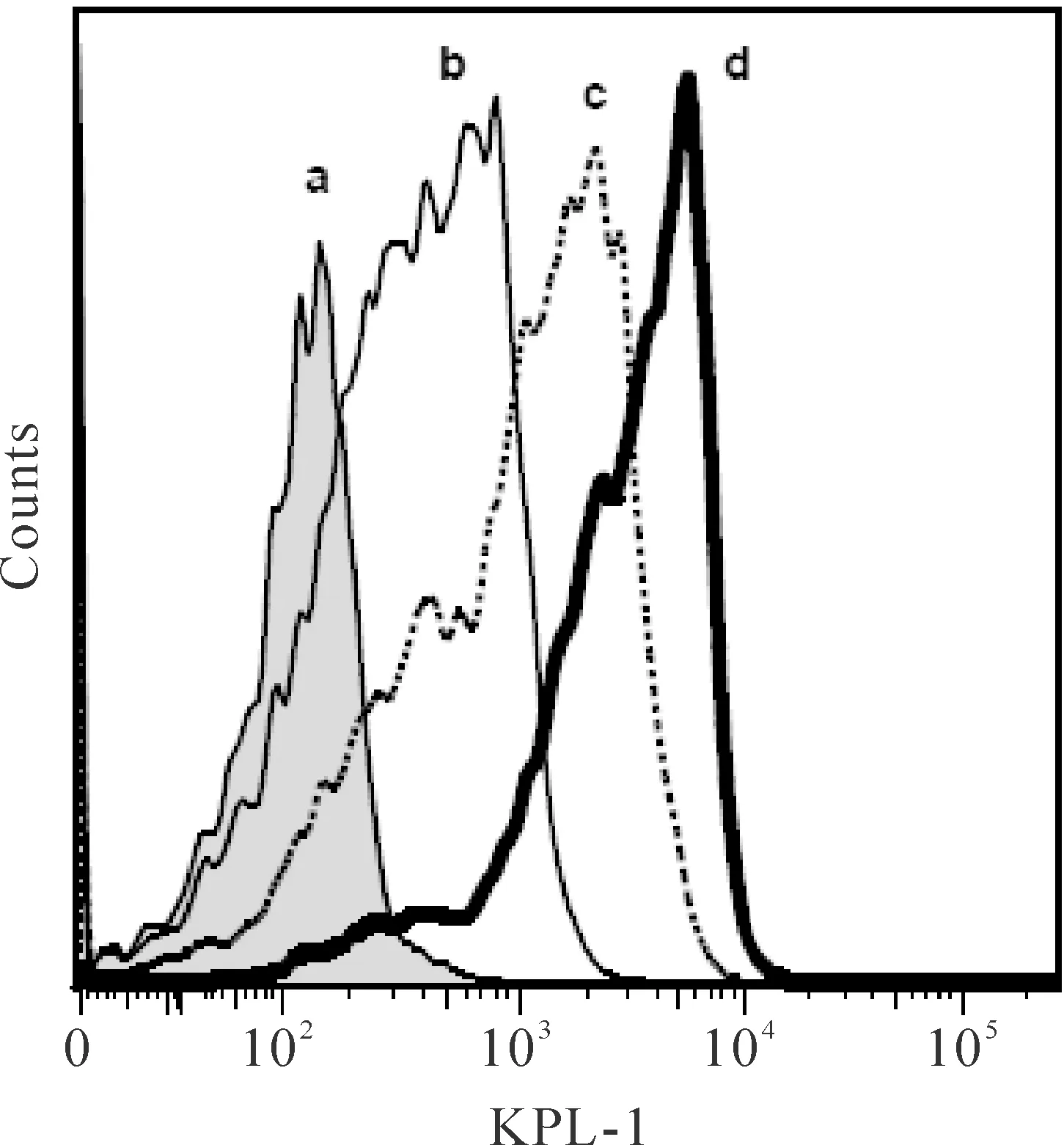
Figure 2.The effects of TAP-SSL5 and SSL5 on the binding of KPL-1 to Jurkat cells. a: isotype control; b,c: Jurkat cells pre-incubated with 10 mg/L SSL5(b) or TAP-SSL5(c), the binding of KPL-1 was significantly inhibited; d:Jurkat cells express abundant of PSGL-1, the binding rate of KPL-1 was 92.8%.
3 TAP-SSL5抑制ADP激活的血小板與Jurkat細胞的結合
在baseline組,未激活血小板與Jurkat細胞的結合率為(8.85±2.79)% ;在control組,血小板經20 μmol/L ADP 激活后與Jurkat細胞的結合率增加至 (11.86±4.49)% ,與baseline組比較差異有統計學意義(P<0.05);Jurkat細胞經1 mg/L TAP-SSL5預處理后,其與ADP 激活血小板的結合率降至(8.15±2.43)%; Jurkat細胞經10 mg/L TAP-SSL5 或10 mg/L SSL5預處理后,其與ADP 激活血小板的結合率分別為(6.73±2.71)% 和 (5.69±1.74)%,與control組比較均顯著降低 (P<0.05);說明10 mg/L 的TAP-SSL5 和 SSL5均可顯著抑制血小板與Jurkat細胞的結合,見圖3。
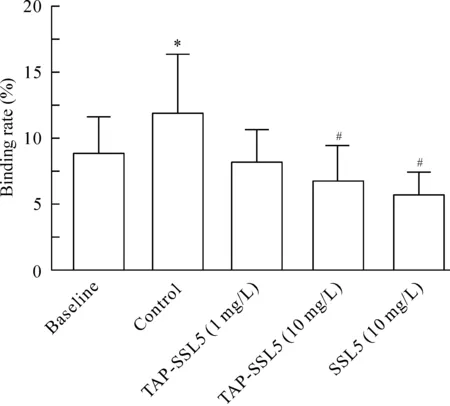
Figure 3.The effect of TAP-SSL5 on the binding of ADP-activated platelets to Jurkat cells. Mean±SD. n=5. *P<0.05 vs baseline; #P<0.05 vs control.
4 TAP-SSL5抑制ADP激活的血小板與外周血淋巴細胞的結合
在baseline組,未激活血小板與淋巴細胞的結合率為(4.99±0.64)%;在control組,血小板經20 μmol/L ADP 激活后與淋巴細胞的結合率增加至 (8.32±1.00)%,與baseline組比較差異有統計學意義(P<0.05);淋巴細胞經1 mg/L TAP-SSL5預處理后,其與ADP 激活血小板的結合率降至(7.02±0.82)%,淋巴細胞經10 mg/L TAP-SSL5 或10 mg/L SSL5預處理后,其與ADP 激活血小板的結合百分率分別為(5.51±0.70)% 和 (6.15±0.57)%,與control組比較均顯著降低 (P<0.05);說明1 mg/L、10 mg/L 的TAP-SSL5 和10 mg/L的 SSL5均可顯著抑制血小板與淋巴細胞的結合,見圖4。
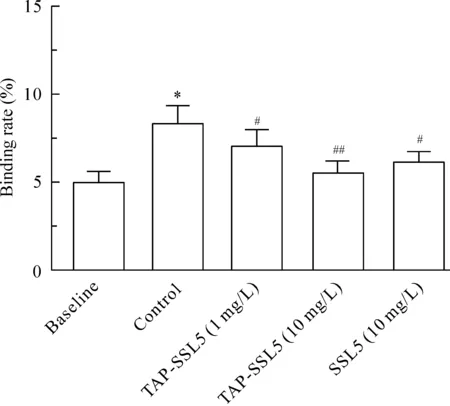
Figure 4.The effect of TAP-SSL5 on the binding of ADP-activated platelets to lymphocytes.Mean±SD. n=3. *P<0.05 vs baseline; #P<0.05,##P<0.01 vs control.
討 論
傳統觀點認為動脈粥樣硬化源于血管壁的脂質沉積,但越來越多的證據顯示炎癥反應在AS的病理過程中發揮重要作用[7-8],同時它還受到免疫機制的嚴密調控[9]。血小板和淋巴細胞是血栓和免疫炎癥過程中的重要細胞成分,它們間的相互作用在血栓形成、炎癥反應、機體免疫等病理生理過程中都顯示出重要的調控作用[1]。血小板和淋巴細胞間可以通過直接接觸和分泌可溶性介質的方式影響彼此功能。血小板可以促進輔助性T細胞(helper T cells,TH)、細胞毒性T細胞(cytotoxic T cells,Tc)、B淋巴細胞及自然殺傷細胞(natural killer,NK)的黏附和滲透;血小板還促進TC細胞的增殖和細胞毒力[10],影響TH細胞的因子分泌和免疫抑制活性[6],引起B淋巴細胞的表型轉換和抗體分泌[10],還可以減輕NK細胞的吞噬能力[11]。這些在動脈粥樣硬化的進程中起到重要的調控作用,針對血小板-淋巴細胞相互作用的干預措施,可能為動脈粥樣硬化的防治提供新的思路和方向。
Li等[12]的研究指出,在血小板同淋巴細胞的結合中,P-選擇素結合PSGL-1是基礎,GPIIb/IIIa、CD11b、CD40L對結合有輔助作用。SSL5是金黃色葡萄球菌的分泌型蛋白,研究發現SSL5可與粒細胞表面的PSGL-1結合而抑制粒細胞在P-選擇素或激活的內皮細胞表面的滾動和黏附[2],SSL5還能夠與HL-60細胞表面的PSGL-1結合而抑制其同血小板或內皮細胞的結合[13]。鑒于炎癥反應和血栓形成在心血管急癥發生中的關鍵作用,在前期研究中,我們將SSL5與凝血因子Xa 的強效抑制劑TAP聯合,成功構建了具有抗炎抗凝雙效功能的融合蛋白TAP-SSL5,并證實TAP-SSL5可抑制粒細胞在P-選擇素表面的黏附,還可抑制激活的血小板與粒細胞的結合。在此基礎上,本研究進一步探討了融合蛋白TAP-SSL5對血小板與淋巴細胞結合的影響。
流式細胞儀檢測結果顯示,人T淋巴細胞株Jurkat細胞表面有豐富的PSGL-1的表達。細胞毒性試驗顯示,終濃度為30 mg/L或以下的融合蛋白TAP-SSL5及10 mg/L的SSL5作用于細胞24 h后,對Jurkat細胞存活率無明顯影響,提示該濃度的蛋白適合于淋巴細胞的體外實驗。10 mg/L的TAP-SSL5或SSL5預處理細胞后,KPL-1同Jurkat細胞的結合明顯減低,提示TAP-SSL5及SSL5能夠同Jurkat細胞表面的PSGL-1結合而競爭性抑制KPL-1的結合。血小板激活后,大量P-選擇素由α-顆粒體轉移到細胞表面,進而加強其同白細胞結合的能力。為探討激活血小板同淋巴細胞的結合作用及TAP-SSL5的影響,我們應用免疫磁珠分選的淋巴細胞同洗滌血小板進行研究,純化了反應體系,能夠更加精確分析這2種細胞間的相互作用。本研究小組前期的研究表明,融合蛋白TAP-SSL5具有抗凝血肽TAP的抗凝血活性[3]。本研究進一步顯示,血小板在經ADP激活后,其與淋巴細胞的結合率明顯增加; TAP-SSL5和SSL5可以顯著抑制激活的血小板與淋巴細胞的結合,說明融合蛋白TAP-SSL5保留了SSL5的功能,即通過與細胞表面PSGL-1的結合發揮抑制炎癥的作用[2]。TAP-SSL5及SSL5對激活血小板同淋巴細胞結合的抑制作用主要來源于對“P-選擇素-PSGL-1”相互作用的抑制;課題組前期研究還發現TAP-SSL5及SSL5可與血小板表面的GPIbα結合[14-15],而GPIbα可同白細胞表面的Mac-1結合,由此推測TAP-SSL5及SSL5可能還通過阻斷“GPIbα-Mac-1”途徑抑制血小板同淋巴細胞的結合。
淋巴細胞從循環血流中遷移到炎癥局部,是其發揮對動脈粥樣硬化免疫調控作用的基礎。同其它白細胞一樣,淋巴細胞的遷移由選擇素介導的接觸、滾動,整合素介導的黏附和隨后的跨內皮滲透所實現[1]。血小板和淋巴細胞的結合,不僅加強淋巴細胞在動脈血流條件下同血管內皮的黏附[5],還易化了它們間的物質代謝和相互影響。因此,針對血小板-淋巴細胞結合的干預措施,有望成為動脈粥樣硬化等炎癥性疾病防治的新手段。本研究證實了融合蛋白TAP-SSL5能夠有效抑制激活血小板同淋巴細胞的結合,這種抑制作用在淋巴細胞的不同亞群間是否存在差異,以及對淋巴細胞激活、增殖和分化等功能的影響有待后續進一步研究。
[1] Li N. Platelet-lymphocyte cross-talk[J]. J Leukoc Biol, 2008, 83(5):1069-1078.
[2] Bestebroer J, Poppelier MJ, Ulfman LH, et al. Staphylococcal superantigen-like 5 binds PSGL-1 and inhibits P-selectin-mediated neutrophil rolling[J]. Blood, 2007, 109(7):2936-2943.
[3] 曲小龍,胡厚源,李 敏,等. 抗炎、抗凝雙效融合蛋白TAP-SSL5表達載體的構建及其功能研究[J]. 第三軍醫大學學報, 2010, 32(1): 5-8.
[4] 龔麗莎,房兆飛,胡厚源,等. 融合蛋白TAP-SSL5對血小板與粒細胞結合的影響[J]. 第三軍醫大學學報, 2013, 35(8):754-758.
[5] Spectre G, Zhu L, Ersoy M, et al. Platelets selectively enhance lymphocyte adhesion on subendothelial matrix under arterial flow conditions[J]. Thromb Haemost, 2012, 108(2):328-337.
[6] Gerdes N, Zhu L, Ersoy M, et al. Platelets regulate CD4+T-cell differentiation via multiple chemokines in humans[J]. Thromb Haemost, 2011, 106(2):353-362.
[7] 王建麗,高春榮,李 莉,等. 冠狀動脈外膜炎癥在動脈粥樣硬化病灶形成中的作用[J]. 中國病理生理雜志, 2006, 22(3): 431-434.
[8] 徐索文,劉培慶. C-反應蛋白與動脈粥樣硬化炎癥的關系[J]. 中國病理生理雜志, 2009, 25(12): 2487-2490.
[9] Libby P, Ridker PM,Hansson GK. Progress and challenges in translating the biology of atherosclerosis[J]. Nature, 2011, 473(7347):317-325.
[10]Elzey BD, Tian J, Jensen RJ, et al. Platelet-mediated modulation of adaptive immunity. A communication link between innate and adaptive immune compartments[J]. Immunity, 2003, 19(1):9-19.
[11]Kopp HG, Placke T,Salih HR. Platelet-derived transforming growth factor-beta down-regulates NKG2D thereby inhibiting natural killer cell antitumor reactivity[J]. Cancer Res, 2009, 69(19):7775-7783.
[12]Li N, Ji Q, Hjemdahl P. Platelet-lymphocyte conjugation differs between lymphocyte subpopulations[J]. J Thromb Haemost, 2006, 4(4):874-881.
[13]Walenkamp AM, Bestebroer J, Boer IG, et al. Staphylococcal SSL5 binding to human leukemia cells inhibits cell adhesion to endothelial cells and platelets[J]. Cell Oncol, 2010, 32(1-2):1-10.
[14]Hu H, Armstrong PC, Khalil E, et al. GPVI and GPIbα mediate staphylococcal superantigen-like protein 5 (SSL5) induced platelet activation and direct toward glycans as potential inhibitors[J]. PLoS One, 2011, 6(4):e19190.
[15]程 彥,房兆飛,曲小龍,等. 融合蛋白TAP-SSL5對人血小板功能的影響[J]. 第三軍醫大學學報, 2012, 34(6):477-480.
Effect of TAP-SSL5 fusion protein on binding of activated platelets to human lymphocytes
PENG Song, BEI Jun-jie, HU Hou-yuan, CHEN Qiang
(DepartmentofCardiology,SouthwestHospital,ThirdMilitaryMedicalUniversity,Chongqing400038,China.E-mail:houyuanhu@hotmail.com)
AIM: To study the effect of tick anticoagulant peptide-staphylococcal superantigen like protein 5 (TAP-SSL5), an anti-inflammatory and anticoagulant fusion protein, on the binding of activated platelets to human lymphocytes.METHODS: Human periphery lymphocytes were isolated by magnetic activated cell sorting (MACS). The toxicity of TAP-SSL5 on the viability of Jurkat cell was assessed by CCK-8 assay. Flow cytometry was applied to detect the expression of CD162 (PSGL-1) on the Jurkat cells (human peripheral blood leukemia T lymphocyte cell line) and the inhibitory effect of TAP-SSL5 on the binding of mouse anti-human CD162 monoclonal antibody (KPL-1) to Jurkat cells. Platelets were activated by ADP at concentration of 20 μmol/L, the binding rates of activated platelets to Jurkat cells or human lymphocytes were assayed by flow cytometry. RESULTS: The concentration of TAP-SSL5 below 30 mg/L didn’t affect the viability of Jurkat cells. TAP-SSL5 at 10 mg/L competitively inhibited KPL-1 binding to Jurkat cells. The binding rates of activated platelets to Jurkat cells or lymphocytes were (11.86±4.49)% and (8.32±1.00)%, respectively, which decreased to (6.73±2.71)% and (5.51±0.70)% after the Jurkat cells and lymphocytes were pre-incubated with 10 mg/L TAP-SSL5 (P<0.05).CONCLUSION: TAP-SSL5 binds to PSGL-1 expressed on lymphocyte surface and directly inhibits the binding of activated platelets to human lymphocytes, which may be one of the anti-inflammatory mechanisms of TAP-SSL5.
Tick anticoagulant peptide; Staphylococcal superantigen like protein 5; Fusion protein; Platelets; Lymphocytes; P-selectin glycoprotein ligand 1
1000- 4718(2015)01- 0023- 05
2014- 08- 12
2014- 09- 30
國家重大新藥創制課題(No. 2013ZX09103003-001);國家自然科學基金資助項目(No. 81270362)
△通訊作者 Tel: 023-68765167; E-mail: houyuanhu@hotmail.com
R363; R331
A
10.3969/j.issn.1000- 4718.2015.01.005
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
今日農業(2021年19期)2022-01-12 06:16:36
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年11期)2021-08-22 03:15:44
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
無線電工程(2020年11期)2020-10-29 01:25:46
