不同肌肉特異性啟動子IGF2表達載體構建及對牛骨骼肌衛星細胞增殖的影響
2015-03-22 08:48:15孫曉麗李樹峰佟慧麗張偉偉殷紅艷嚴云勤
畜牧獸醫學報 2015年4期
關鍵詞:檢測
孫曉麗,李樹峰,佟慧麗,張偉偉,殷紅艷,嚴云勤
(東北農業大學生命科學學院,哈爾濱 150030)
不同肌肉特異性啟動子IGF2表達載體構建及對牛骨骼肌衛星細胞增殖的影響
孫曉麗,李樹峰,佟慧麗,張偉偉,殷紅艷,嚴云勤*
(東北農業大學生命科學學院,哈爾濱 150030)
旨在獲得帶有高效特異性啟動子的IGF2表達載體,研究其對牛骨骼肌衛星細胞增殖的影響。本研究將構建的3種含有IGF2肌肉特異性啟動子的真核表達載體轉染牛骨骼肌衛星細胞和胎兒成纖維細胞,RT-PCR檢測IGF2的相對表達量,從而篩選高效特異性的載體,并進行EdU試驗及細胞周期相關基因RT-PCR檢測。結果,成功構建了不同啟動子desminpro、CMV-MyoGpro和MyoGpro-double表達IGF2的載體。RT-PCR檢測結果表明,pGL3-desminpro-IGF2是較高效的并具有特異性的載體。載體轉染體外培養細胞提高了細胞增殖速度,并上調了細胞周期相關基因CDK6和IGF2的表達。這為轉基因肉牛的生產奠定了重要基礎。
IGF2;肌肉特異性啟動子;骨骼肌衛星細胞;細胞增殖
胰島素樣生長因子2 (Insulin-like growth factor 2,IGF2)是由67個氨基酸殘基組成的蛋白質[1-2],是骨骼肌生長發育的重要調控因子[3],它調控細胞增殖、分化以及程序性細胞死亡,對個體生長和發育具有重要作用[4-5]。研究表明,與野生型小鼠相比,IGF2基因敲除小鼠的體重減少了40%[6],S.Hayashi等得到了同樣的研究結果,證明IGF2 在牛的骨骼肌再生過程中發揮著重要的作用[7]。近年來,對IGF2在促增殖方面的研究越來越多,但通過轉基因方法探索IGF2對家畜肌肉生長影響的研究還較少。
為了提高外源基因在肌肉組織中的表達效率,啟動子的選擇是一個重要的因素。特異高效啟動子可以驅動外源基因在真核細胞中表達[8]。目前,巨細胞病毒(CMV)啟動子是外源基因在真核細胞中表達常選用的啟動子之一,CMV啟動子雖然高效但幾乎沒有細胞特異性,而且考慮到轉基因的生物安全性,不能將其應用于轉基因肉牛的生產中。本研究構建出3種含有肌肉特異性啟動子的IGF2真核表達載體,通過細胞轉染及IGF2表達量檢測,篩選出具有較高活性和較強肌肉特異性的IGF2表達載體,使其能夠促進肌肉細胞的增殖,為提高肌肉產量的轉基因肉牛的生產提供重要幫助。
1 材料與方法
1.1 試驗材料
1.1.1 菌株和載體 菌株DH5α、pMD18-T克隆載體購于大連TaKaRa生物技術有限公司;pcDNA3.1(+),pMD18-T-desminpro,pMD18-T-CMV-MyoGpro pMD18-T-MyoGpro-double菌株由本實驗室提供。1.1.2 工具酶、試劑盒和其他試劑 限制性內切酶和 LATaq 聚合酶等購于大連TaKaRa生物技術有限公司;Tissue DNA Kit I和Endo-Free PlasmidMini kit II購于Omega公司;Plasmid Mini Kit I和Gel Extraction Kit I購于北京TransGen生物技術有限公司;Trizol試劑購于 Invitrogen 公司;SYBR?PrimeScriptTMRT-PCR Kit II 購于大連TaKaRa生物技術有限公司;Cell-LightTMEdU Apllo 488InvitroKit購于廣州銳博生物科技有限公司;蛋白胨、瓊脂糖、酵母提取物、氯化鈉等購自Sigma公司,DMEM、Opti-MEM、胎牛血清和馬血清購自Gibco公司。
1.2 方法
1.2.1 牛IGF2基因的克隆與測序 根據GenBank公布的牛IGF2基因序列AC_000186.1,設計IGF2引物,上游引物F:ccggaattcTCAATGGGGATCACAGCAGGAAAGT(小寫部分為KpnⅠ酶切位點);下游引物R:cccaagcttGCTCACTTCTAATCGCTGGATGCCT(小寫部分為EcoRⅠ酶切位點),引物由上海生工生物工程有限公司合成。以牛全基因組為模板擴增IGF2基因全序列。用1%瓊脂糖凝膠電泳檢測,并用凝膠回收試劑盒進行回收。回收PCR產物與PMD-18T克隆載體連接,雙酶切鑒定正確后命名為pMD-18T-IGF2,送由北京英駿公司測序。
1.2.2 表達載體pcDNA3.1(+)-IGF2的構建 用限制性內切酶KpnⅠ和EcoRⅠ雙酶切獲得目的片段,用同樣的酶雙酶切pcDNA3.1(+)載體,回收純化后,用T4 DNA連接酶連接轉化至DH5α感受態細胞中,挑選單克隆,提取質粒并酶切鑒定,成功構建pcDNA3.1(+)-IGF2載體。
1.2.3 pGL3-desminpro-IGF2、pGL3-CMV-MyoGpro-IGF2、pGL3-MyoGpro-Double-IGF2 重組質粒的構建 用限制性內切酶NheⅠ和Hind Ⅲ分別雙酶切pMD18-T-desminpro、pMD18-T-CMV-MyoGpro、pMD18-T-MyoGpro-double獲得desminpro、CMV-MyoGpro、MyoGpro-double 3種肌肉特異性啟動子,膠回收小片段,用同樣的酶雙酶切pcDNA3.1(+)-IGF2載體,用T4 DNA連接酶連接,轉化DH5α感受態細胞中,挑選單克隆,提取質粒并酶切鑒定,構建3種中間載體,然后用NheⅠ和XhoⅠ分別雙酶切3個中間載體膠回收小片段,用同樣的酶雙酶切pGL3-Basic,用T4 DNA連接酶連接轉化DH5α感受態細胞中,挑選單克隆,提取質粒并酶切鑒定,構建3種最終載體。
1.2.4 熒光定量RT-PCR檢測相關基因的表達變化 Lipofectamine 2000 Reagent將構建3種重組質粒分別轉染牛胎兒成纖維細胞和牛骨骼肌衛星細胞,同時設置陽性對照、陰性對照和空白對照。48 h后用TRIzol法提取各細胞總RNA,反轉錄后進行RT-PCR檢測。分別檢測IGF2、P27、CDK6基因mRNA相對表達量的變化情況,每個樣品設置3個重復。RT-PCR擴增體系:SYBR Premix Ex TaqTMⅡ 10.0 μL,上下游引物(10 μmol·L-1)各1 μL,cDNA 2.0 μL,ROX Reference DyeⅡ 0.4 μL,無菌水補平體系至20 μL。RT-PCR反應條件: 95 ℃ 2 min;95 ℃ 15 s,60 ℃ 40 s,共45個循環;95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。引物序列見表1。
1.2.5 EdU檢測 用Lipofectamine 2000 Reagent將pGL3-Basic和pGL3-desminpro-IGF2兩種質粒轉染接種于12孔板中細胞匯合度為90%的牛骨骼肌衛星細胞,48 h后,用EdU 試劑盒檢測細胞增殖率。
表1 熒光定量引物序列及擴增長度
Table1 The sequences of qRT-PCR primers and the product length

基因Gene引物序列(5'-3')Primersequence擴增長度/bpProductlength退火溫度/℃AnnealingtemperatureIGF2S:GCCTTCGCCTCGTGCTGCTAA:ACGGCTGCGTCGGTTTATGC14460P27S:GGCGTCAGACGTAAAA:CTTGGAGTCAGCGATA12149CDK6S:ATAAGCCAAGTTTCCCCAAGTGA:TGAGTGCTAAGGCAACCAGACA30258RPS18S:GTGGTGTTGAGGAAAGCAGACAA:TGATCACACGTTCCACCTCATC7960
1.2.6 統計分析 應用SPSS16.0軟件包計算各組試驗數據平均值、標準差及標準誤差,并對各組試驗數據之間的差異顯著性進行單因素方差分析。
2 結 果
2.1 牛IGF2基因的克隆與中間載體pcDNA3.1(+)-IGF2的構建
以牛骨骼肌衛星細胞基因組DNA為模板,用IGF2引物進行PCR擴增,1%瓊脂糖凝膠電泳檢測,在相應位置出現明亮條帶(圖1)。測序結果分析表明,IGF2測序結果與GenBank發表序列的同源性為100%,表明成功擴增牛的IGF2基因。將pcDNA3.1(+)-IGF2重組質粒用KpnI和EcoR I進行雙酶切鑒定,電泳結果顯示,在5 428和551 bp左右處各有一條特異性條帶(圖1),與預期結果相符。
2.2 不同啟動子表達牛IGF2載體的構建與鑒定
pGL3-desminpro-IGF2、pGL3-CMV-MyoGpro-IGF2 和pGL3-MyoGpro- Double-IGF2 3種重組質粒分別用NheⅠ和XhoⅠ進行雙酶切鑒定,電泳結果顯示,pGL3-desminpro-IGF2在4 818和851 bp處各有一條特異性條帶,pGL3-CMV-MyoGpro-IGF2在4 818和1 373 bp處有一條特異性條帶,pGL3-MyoGpro- Double-IGF2在4 818 和1 090 bp處各有一條特異性條帶(圖1),結果表明,3種肌肉特異性啟動子和IGF2基因的CDS區成功連接到重組質粒中,與預期結果相符。
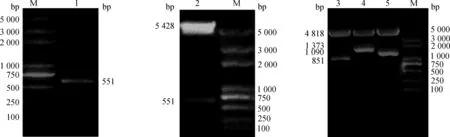
M.DNA相對分子質量標準DL5000;1.PCR擴增IGF2 CDS區結果;2.pcDNA3.1(+)-IGF2的Kpn I和EcoR I雙酶切結果;3.pGL3-desminpro-IGF2的NheⅠ和XhoⅠ雙酶切結果;4.pGL3-CMV-MyoGpro-IGF2的NheⅠ和Xho Ⅰ雙酶切結果;5.pGL3-MyoGpro- Double-IGF2的NheⅠ和XhoⅠ雙酶切結果M.DL5000 marker;1. PCR product of IGF2 CDS;2.pcDNA3.1(+)-IGF2 plasmid digested with Kpn Ⅰ and EcoR Ⅰ;3.pGL3-desminpro-IGF2 plasmid digested with Nhe Ⅰ and Xho Ⅰ;4.pGL3-CMV-MyoGpro-IGF2 plasmid digested with Nhe Ⅰ and Xho Ⅰ;5.pGL3-MyoGpro- Double-IGF2 plasmid digested with Nhe Ⅰ and Xho Ⅰ圖1 PCR擴增結果和重組質粒的雙酶切結果Fig.1 The result of PCR amplification and the recombinant plasmid by double digested with restriction enzymes
2.3 RT-PCR檢測3種重組質粒轉染細胞后IGF2的表達變化
將3種重組質粒轉染牛骨骼肌衛星細胞和胎兒成纖維細胞,同時設置pcDNA3.1(+)-IGF2為陽性對照,pGL3-Basic為陰性對照,control為空白對照,48 h以后收集細胞。熒光定量RT-PCR結果顯示,pGL3-desminpro-IGF2組的IGF2的表達水平顯著高于其他兩組(P<0.05),且具有較好的肌肉特異性(圖2),所以選擇效果較好的pGL3-desminpro-IGF2質粒用于后續試驗。
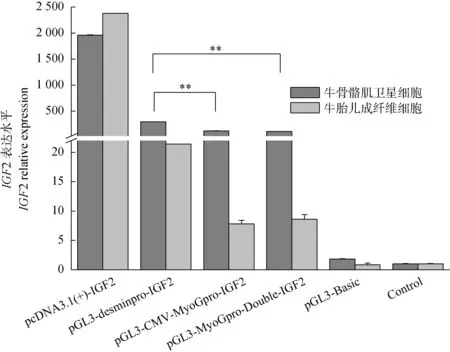
*.差異顯著(P<0.05);** .差異極顯著(P<0.01)。圖4、圖5同*.Means differ significantly (P<0.05);** .Means highly differ significantly (P<0.01).The same as Figure 4 and Figure 5圖2 各組間IGF2 mRNA的相對表達量Fig.2 IGF2 relative expression in different groups
2.4 EdU檢測轉染pGL3-desminpro-IGF2后對細胞增殖的影響
重組質粒pGL3-desminpro-IGF2和對照組pGL3-Basic轉染牛骨骼肌衛星細胞,48 h后用EdU試劑盒檢測增殖變化(圖3),與對照組相比,轉染pGL3-desminpro-IGF2后細胞增殖率為17.95%,轉染pGL3-Basic后細胞增殖率為14.10%,pGL3-desminpro-IGF2與對照組相比增殖率為27.31%(圖4),結果表明pGL3-desminpro-IGF2能夠促進牛骨骼肌衛星細胞增殖。
2.5 RT-PCR檢測轉染pGL3-desminpro-IGF2后細胞周期相關基因的表達變化
用pGL3-desminpro-IGF2轉染牛骨胳肌衛星細胞,48 h后,熒光定量RT-PCR檢測IGF2、P27和CDK6的表達變化。與對照組比較,試驗組細胞P27表達下降,而CDK6表達上升,差異均顯著(P<0.05)(圖5)。
3 討 論
近年來,IGF2促進肌肉增殖的相關研究越來越受到關注,細胞試驗證明,IGF2是肌細胞生長過程中的自分泌信號。A.Levinovitz等報道,在大鼠的骨骼肌再生過程中,IGF2 mRNA的表達水平顯著上調[9]。IGF2對細胞生長的調控作用主要通過對細胞周期的調控實現。IGF2可以刺激DNA合成和細胞復制,推動細胞完成增殖周期[10-11]。本試驗利用的是實驗室前期構建的3種具有特異性且活性相對較高的3種啟動子,Desminpro(300)是牛結蛋白基因大小為300 bp啟動子,它活性較高,其啟動基因表達的能力是人骨骼肌α-actin啟動子和肌酸激酶啟動子的2倍[12];CMV-MyoGpro是MyoG基因啟動子之前連接了一段CMV增強子序列,在C2C12細胞中CMV-MyoGpro復合啟動子的轉錄活性較牛MyoG啟動子而言提高了9倍左右,達到了強啟動子CMV啟動子的72%,相比牛骨骼肌衛星細胞,CMV增強子提高轉錄活性的效果更明顯;MyoGpro-double 是含有兩個拷貝數調控元件的MyoG啟動子片段,其轉錄活性顯著高于普通MyoG基因啟動子,約為MyoG基因啟動子轉錄活性的3.8倍[13],3種啟動子都具有較高的活性和較好的肌肉特異性[12-14]。本研究克隆了長度為551 bp牛IGF2基因的CDS序列,并構建出帶有相應肌肉特異性啟動子IGF2真核表達載體,即pGL3-desminpro-IGF2、pGL3-CMV-MyoGpro-IGF2和pGL3-MyoGpro-Double-IGF2。pGL3-desminpro-IGF2載體轉染牛骨骼肌衛星細胞后能夠高效特異性表達,且能夠促進牛骨骼肌衛星細胞增殖。IGF2通過下調CDKs蛋白抑制因子P27的表達,上調CDK6的表達,加速細胞從G1期進入到S期的轉變[15-16],進而發揮其對牛骨骼肌衛星細胞的增殖作用。本研究通過熒光定量RT-PCR檢測P27和CDK6的表達,結果顯示,P27表達下降,而CDK6表達上升,均呈顯著性差異(P<0.05),這與文獻報道一致。pGL3-desminpro-IGF2在牛骨骼肌衛星細胞中高效特異性表達,并促進成肌細胞的生長,推測其原因除了在desminpro肌肉特異性啟動子的啟動作用之外,IGF2的高表達在肌肉細胞生長方面發揮了重要的作用,因此該表達載體的應用能夠為轉基因肉牛的生產提供重要幫助。
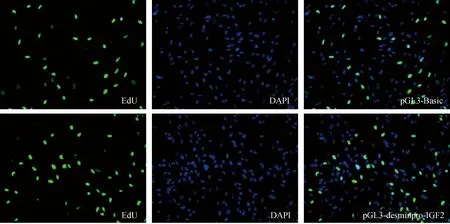
細胞轉染pGL3-Basic和pGL3-desminpro-IGF2,48 h后用EdU試劑盒檢測增殖變化,綠色代表EdU陽性細胞,藍色為DAPI染色的細胞核Cells transfected with pGL3-Basic and pGL3-desminpro-IGF2 were detected by EdU kit for proliferation changes,green is for EdU positive cells,blue is DAPI staining of nuclei圖3 EdU增殖試驗結果 200×Fig.3 Proliferation results of EdU 200×
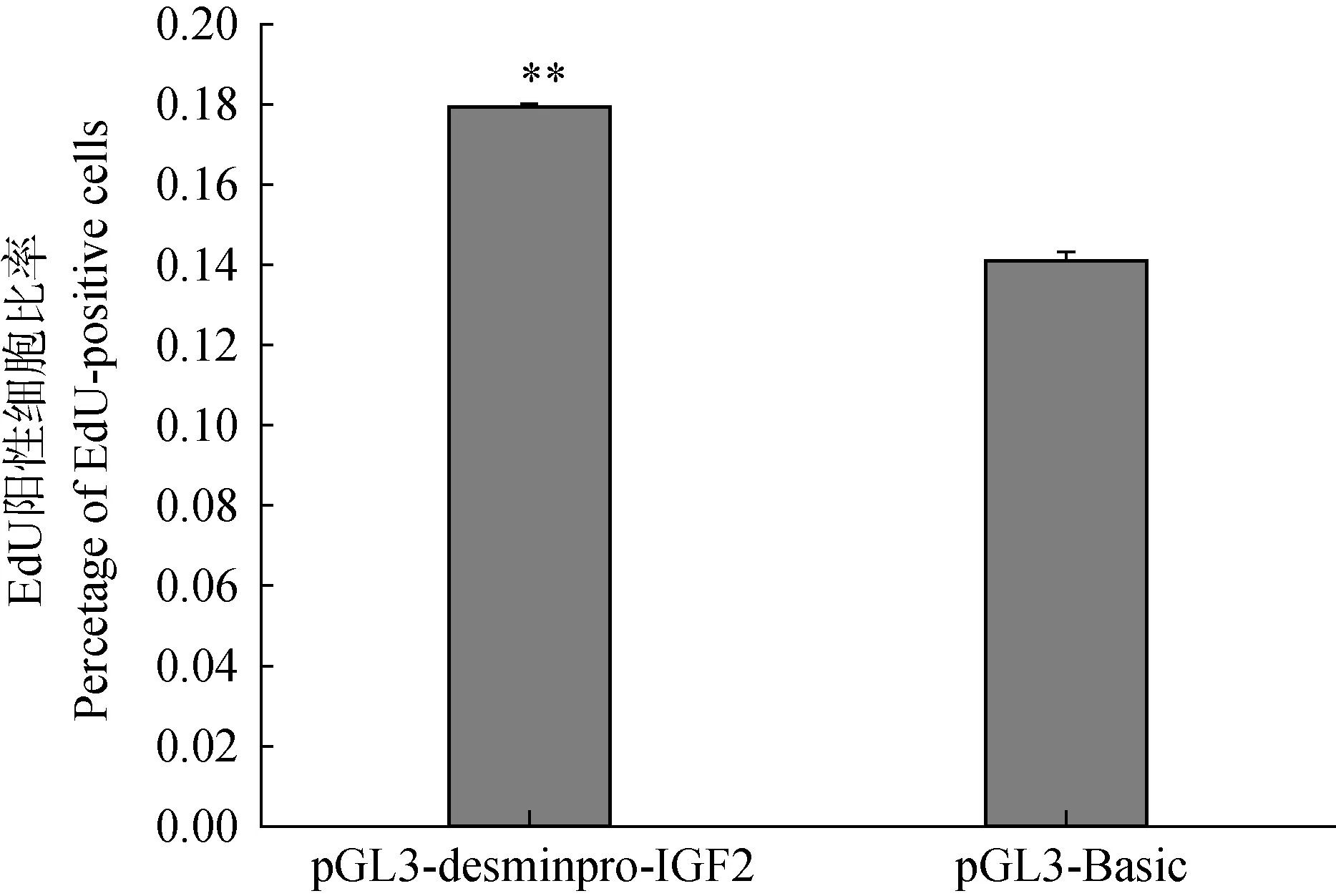
圖4 EdU陽性細胞百分率Fig.4 Percentage of EdU-positive cells
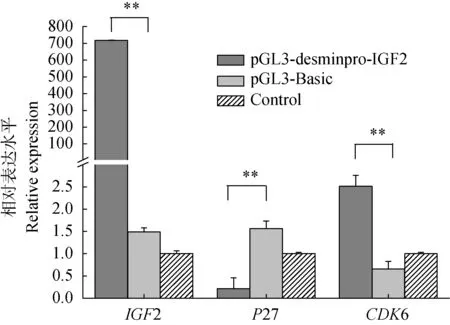
圖5 各組間不同基因mRNA相對表達量Fig.5 Different genes relative expression in different groups
4 結 論
本研究獲得了能夠在牛骨骼肌衛星細胞中高效特異性表達的真核表達載體pGL3-desminpro-IGF2,其在牛骨骼肌衛星細胞的高效特異性表達,并促進成肌細胞的增殖。
[1] MICKE G C,SULLIVAN T M,MCMILLEN I C,et al.Protein intake during gestation affects postnatal bovine skeletal muscle growth and relative expression of IGF1,IGF1R,IGF2 and IGF2R[J].MolCellEndocrinol,2011,332(1):234-241.
[2] 朱 嬋,徐勇飛.IGF2 的生物學特性及其與疾病關系的研究進展[J].國際檢驗醫學雜志,2010,(9):966-969. ZHU C,XU Y F.Biological characteristics of IGF2 and Research progress of its relationship with disease [J].InternationalJournalofLaboratoryMedicine,2010,(9):966-969.(in Chinese)
[3] GERRARD D E,OKAMURA C S,RANALLETTA M A,et al.Developmental expression and location of IGF-I and IGF-II mRNA and protein in skeletal muscle[J].JAnimSci,1998,76(4):1004-1011.
[4] 彭鳳蘭.胰島素樣生長因子 2 (Igf2) 研究進展[J].實用預防醫學,2008,14(6):1963-1966. PENG F L.The research progress of insulin-like growth factor2 (Igf2) [J].PracticalPreventiveMedicine,2008,14(6):1963-1966.(in Chinese)
[5] CONSTA^NCIA M,HEMBERGER M,HUGHES J,et al.Placental-specific IGF-II is a major modulator of placental and fetal growth[J].Nature,2002,417(6892):945-948.
[6] DECHIARA T M,ROBERTSON E J,EFSTRATIADIS A.Parental imprinting of the mouse insulin-like growth factor II gene[J].Cell,1991,64(4):849-859.
[7] HAYASHI S,ASO H,WATANABE K,et al.Sequence of IGF-I,IGFⅡ,and HGF expression in regenerating skeletal muscle [J].HistochemCellBiol,2004,122(5):427-434.
[8] 袁建龍,王 瑋,金 永,等.人工合成啟動子特異啟動 IGF-1真核載體的構建與綿羊成纖維細胞轉染[J].內蒙古大學學報:自然科學版,2010,41(6):678-682. YUAN J L,WANG W,JIN Y,et al.The construction of synthetic promoter promote IGF-1 eukaryotic vector and transfected into sheep fibroblasts [J].InnerMongoliaUniversity:NaturalScience,2010,41(6):678-682.(in Chinese)
[9] LEVINOVITZ A,JENNISCHE E,OLDFORS A,et al.Activation of insulin-like growth factor II expression during skeletal muscle regeneration in the rat:correlation with myotube formation[J].MolEndocrinol(Baltimore,Md.),1992,6(8):1227-1234.
[10] 李 聰,馬凱麗,劉增力,等.IGFs 及其信號通路研究進展[J].生物技術通報,2013,(9):43-46. LI C,MA K L,LIU Z L,et al.The research progress of IGFs and its signaling pathways [J].BiotechnologyBulletin,2013,(9):43-46.(in Chinese)
[11] DUPONT J,KARAS M,LEROITH D.The potentiation of estrogen on insulin-like growth factor I action in MCF-7 human breast cancer cells includes cell cycle components[J].JBiolChem,2000,275(46):35893-35901.
[12] 杜 巍,李樹峰,佟慧麗,等.牛結蛋白基因啟動子的克隆及功能的初步分析[J].畜牧獸醫學報,2014,45(1):56-61. DU W,LI S F,THONG H L,et al.Cloning and functional analysis of bovine desmin gene promoter[J].ActaVeterinariaetZootechnicaSinica,2014,45(1):56-61.(in Chinese)
[13] WANG X,LU M,FENG L H,et al.Effects of CMV enhancer on activity and specificity of bovineMyoGgene promoter[J].JNortheastAgricUniversity,2013,20(4):34-38.
[14] 王秋華,曹允考,李樹峰,等.牛MyoG基因啟動子的克隆及功能的初步分析[J].畜牧獸醫學報,2012,43(1):37-43. WANG Q H,CAO Y K,LI S F,et al.Cloning and functional analysis of bovineMyoGgene promoter[J].ActaVeterinariaetZootechnicaSinica,2012,43(1):37-43.(in Chinese)
[15] DUPONT J,PIERRE A,FROMENT P,et al.The insulin-like growth factor axis in cell cycle progression[J].HormMetabRes,2003,35(11/12):740-750.
[16] 董雅靜.IGF2 真核表達載體的構建及綿羊骨骼肌細胞轉基因研究[D].楊凌:西北農林科技大學,2012. DONG Y J.The construction of IGF2 eukaryotic vector and the gene transfer in sheep skeletal muscle cells[D].Yangling:Northwest A&F University,2012.(in Chinese)
(編輯 郭云雁)
The Construction of Different Muscle-specific Promoter IGF2 Expression Vector and Its Impact on Bovine Skeletal Muscle Satellite Cell Proliferation
SUN Xiao-li,LI Shu-feng,TONG Hui-li,ZHANG Wei-wei,YIN Hong-yan,YAN Yun-qin*
(CollegeofLifeScience,NortheastAgriculturalUniversity,Harbin150030,China)
The aim of this study was to construct IGF2 expression vector with efficient and specific promoter and study its effects on bovine skeletal muscle satellite cell proliferation.The 3 eukaryotic expression vectors containing IGF2 and muscle-specific promoter were transfected into bovine skeletal muscle satellite cells and fetal fibroblasts,the relative expression of IGF2 was detected by RT-PCR to screen efficient and specific vector.EdU experiments were carried out and RT-PCR was used to detect the cell cycle-related genes.The different promoter (desminpro,CMV-MyoGpro and MyoGpro-double) IGF2 expression vectors were constructed successfully.The test results of RT-PCR showed that pGL3-desminpro-IGF2 was an efficient and specific vector.The vector transfected into cultured cellinvitroincreased the speed of cell proliferation and up-regulated the expression of cell cycle-related genes,CDK6 andIGF2.The result has laid an important foundation for the production of transgenic bovine.
IGF2;muscle-specific promoter;skeletal muscle satellite cells;cell proliferation
10.11843/j.issn.0366-6964.2015.04.007
2014-07-01
國家轉基因專項“高產優質轉基因肉牛新品種培育”(2011ZX08007-002)
孫曉麗(1988-),黑龍江齊齊哈爾人,碩士,主要從事分子克隆與細胞增殖的研究,E-mail:965080867@qq.com
*通信作者:嚴云勤,教授,博士生導師,主要從事動物轉基因的研究,E-mail:yanyunqin@sohu.com
S823;S813.3
A
0366-6964(2015)04-0555-06
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
