貴州東部常綠落葉闊葉混交林碳素積累及其分配特征
2015-03-15 07:16:19丁訪軍潘忠松崔迎春周鳳嬌
生態學報 2015年6期
丁訪軍,潘忠松,吳 鵬,崔迎春,周鳳嬌
1 貴州省林業科學研究院, 貴陽 550005 2 貴州省三都縣林業局, 三都 558100
貴州東部常綠落葉闊葉混交林碳素積累及其分配特征
丁訪軍1,*,潘忠松2,吳 鵬1,崔迎春1,周鳳嬌1
1 貴州省林業科學研究院, 貴陽 550005 2 貴州省三都縣林業局, 三都 558100
以雷公山自然保護區常綠落葉闊葉混交林為研究對象,對其碳素含量、碳密度及分配特征進行了研究。結果表明:生態系統碳素含量表現為喬木層(418.58 g/kg)>灌木層(387.26 g/kg)>草本層(382.80 g/kg)>枯落物層(378.11 g/kg)>土壤層(31.48 g/kg),差異極顯著(P﹤0.01),喬木不同器官表現為干>根>葉>枝,差異不顯著(P>0.05),灌、草層均表現為地上>地下,土壤碳素含量隨土層深度的增加而減少;生態系統碳密度為234.68 t/hm2,表現為土壤層(170.00 t/hm2)>喬木層(57.02 t/hm2)>枯枝落葉層(5.48 t/hm2)>灌木層(1.81 t/hm2)>草本層(0.37 t/hm2),分別占生態系統碳密度的72.44%、24.30%、2.34%、0.77%和0.16%;植被層碳密度為58.79 t/hm2,占了生態系統碳密度的25.09%;喬木層各器官以樹干的碳密度最高,占了喬木層碳密度52.43%;灌木層、草本層地上部分碳密度分別是地下部分的2.85倍1.64倍;土壤表層(0—20 cm)碳密度為70.40 t/hm2,顯著高于其它各層(P﹤0.001),占了土壤(0—80 cm)碳密度的41.41%,有很強的表聚性,因此,防止地表的水土流失,可有效保持土壤對碳的吸存。
碳素含量; 碳密度; 常綠落葉闊葉混交林; 雷公山
自工業革命以來,人類大量燃燒化石燃料和改變土地的利用方式等因素導致大氣中的CO2濃度不斷升高[1],由此而引起的氣候變暖問題已嚴重威脅到人類的生存和發展,全球碳循環也因此成了當今研究的熱點問題之一。森林作為陸地生態系統的主體,是地球上最大的陸地碳庫,在調節全球碳平衡、減緩大氣中CO2等溫室氣體濃度上升以及維護全球氣候等方面具有不可替代的作用[2-3]。熱帶、亞熱帶森林在全球森林中占有舉足輕重的地位[4], 其生物量的較小波動即可引起全球碳循環的顯著變化[5]。我國亞熱帶地區植被類型豐富,生物多樣性高,對維持全球碳平衡具有極其重要的作用。近年來,有關亞熱帶典型森林生態系統的碳含量及碳密度研究已取得了較多成果[6-15],但主要集中在針葉林、針葉混交林和常綠闊葉林,而對于常綠落葉闊葉混交林的研究較少,特別是貴州東部中亞熱帶地區尚屬空白,加之森林生態系統的碳儲量和碳密度有很大的空間變異性[16-17]。因此,研究貴州東部中亞熱帶常綠落葉闊葉混交林生態系統的碳素含量、碳密度及其分配特征,對于區域及國家尺度上估算森林生態系統碳庫和碳平衡以及制定森林資源管理策略具有重要意義。
1 研究地概況
雷公山是貴州苗嶺山脈東段主峰,最高峰海拔 2178.8 m,位于東經108°5′—108°24′;北緯 26°15′—26°32′。屬中亞熱帶季風山地濕潤氣候區,冬無嚴寒,夏無酷暑,雨量充沛。最冷月(1月)平均溫山頂-0.8 ℃,山麓4—6 ℃,最熱月(7月)山頂17.6 ℃,山麓23—25.5 ℃,年平均溫度山頂9.2 ℃,山麓14.7—16.3 ℃,年降雨量在1300—1600 mm之間。地帶性植被屬我國中亞熱帶東部偏濕性常綠闊葉林,植被垂直分布明顯。海拔1350 m以下是常綠闊葉林,1350—2100 m是山地常綠落葉闊葉混交林,2100 m以上是高山灌叢。常綠落葉闊葉混交林中落葉樹種以水青岡(Faguslongipetiolata)、亮葉水青岡(Faguslucida)、多脈青岡(Cyclobalanopsismultinervis)、白辛樹(Pterostyraxpsilophyllus)等為主,常綠樹種以栲(Castanopsisfargesii)、石櫟(Lithocarpusglaber)、木蓮(Manglietiafordiana)、木荷(Schimasuperba)等為主;自然保護區內植被保護較好,常綠落葉闊葉混交林物種豐富,群落結構復雜,優勢種不明顯。群落年齡結構也十分復雜,平均年齡約在40a左右。林下灌木主要有狹葉方竹(Chimonobambusaangustifolia)、柃木(Euryajaponica)、圓錐繡球(Hydrangeapaniculata)等。林下草本有樓梯草(Elatostemaumbellatum)、矮冷水花(Pileapeploides)、禾本科(Poaceae)、莎草科(Cyperaceae)、菊科(Asteraceae)及蕨類(Pteridophyta)植物等。
2 研究方法
2.1 樣地設置與調查
本研究于2011年8月在全面踏查的基礎上,選擇林相相對整齊、立地條件相對一致的常綠落葉闊葉混交林代表性地段設置了5塊樣地,樣地面積為30 m×30 m。在每個樣地四角和樣地對角線交叉點按梅花型設置5個2 m×2 m的灌木小樣方、5個1 m×1 m的草本和枯落物層小樣方,調查樣地內喬木樹種(胸徑≥5 cm)的種類、胸徑和樹高,灌、草種類及蓋度等,樣地基本情況見表1。
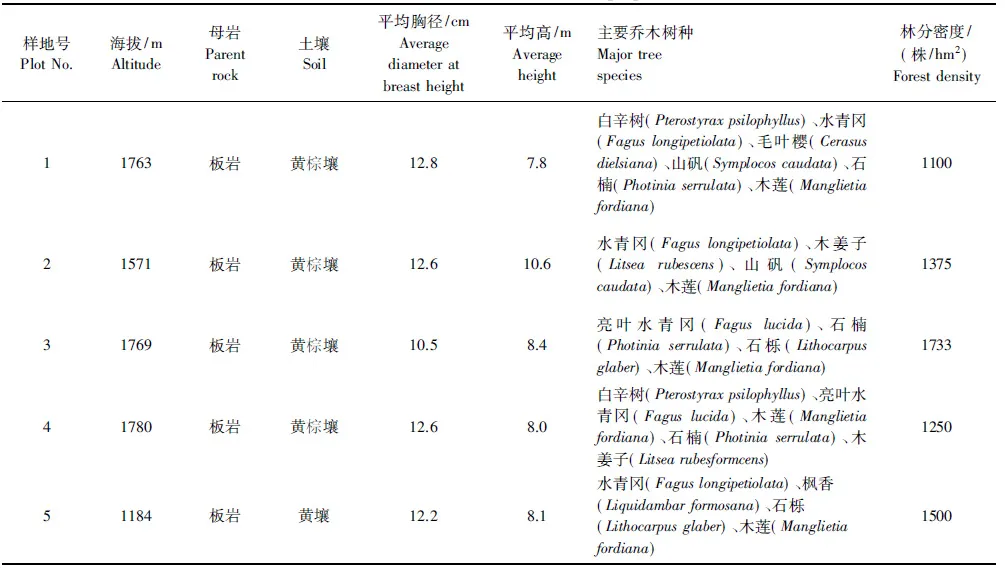
表1 樣地基本情況Table 1 General Situations of the Sample plots
2.2 生物量及碳素含量測定
由于研究樣地位于國家級自然保護區, 不宜進行大量的破壞性采伐,因此在對樣地每木檢尺統計分析的基礎上,根據胸徑總體分布情況,分別常綠和落葉以2—4 cm為一個徑級選擇解析木,株數較多的中央徑級選取2—3株,其他徑級各選取1—2株,共21株解析木(落葉11株,常綠10株)建立混合相對生長方程(表2),并以此計算喬木層不同器官生物量。灌、草層和枯落物層生物量采用樣方收獲法測定。在測定生物量的同時對喬木層的干、枝、葉和根分別取樣,灌木層的葉、莖和根分別取樣,草本層的地上部分和地下部分分別取樣。枯枝落葉層分為未分解層和半分解層分別取樣。樣品的碳含量采用重鉻酸鉀-外加熱法測定[6,13-15]。

表2 生物量相對生長方程Table 2 Equation of Relative Biomass Growth
式中,W代表生物量,D代表胸徑
2.3 土壤樣品采集及分析
在每個樣地內按“S”型隨機挖取5個土壤剖面,按0—20 cm、20—40 cm、40—60 cm和60—80 cm進行分層土樣采集。樣品的有機碳含量采用重鉻酸鉀-外加熱法測定,土壤容重采用環刀法測定。
2.4 碳密度的計算
植被層碳密度為植被層各組分生物量與相應組分的碳素含量的乘積。
土壤有機碳密度計算公式[18]如下:
SOCi=OiHiBi(1-Gi)/100
式中,SOCi是第i層土壤有機碳密度(kg/m2),Hi是第i層土壤的厚度(cm),Bi是第i層土壤的容重(g/cm3),Oi是i層土壤的有機碳含量(g/kg),Gi為直徑≥2mm的石礫所占的體積百分比(%)。
數據處理及分析采用Excel2003和SPSS16.0軟件。
3 結果與分析
3.1 雷公山常綠落葉闊葉混交林生態系統各組分碳素含量
3.1.1 喬木層碳素含量
喬木層不同器官的平均碳素含量為418.58g/kg,表現為干(428.12g/kg)>根(421.18g/kg)>葉(418.91g/kg)>枝(406.11g/kg),不同器官碳素含量的變異系數介于2.09%—8.12%之間(表3),差異不顯著(P>0.05)。喬木層的平均碳素含量低于杉木[13]、馬尾松[7,9-10]、樟樹[11]等亞熱帶地區主要優勢樹種的平均碳素含量。這主要是由于組成樹種不同,導致林木不同器官中的碳素含量明顯不同,即使是同一森林類型,在不同的氣候帶,林木各器官碳素含量也有一定的差異[19]。
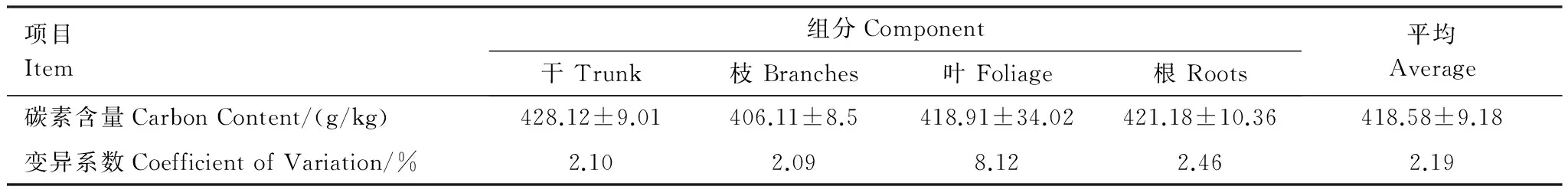
表3 喬木層碳素含量及分配Table 3 Carbon content and distribution among the tree layer
3.1.2 林下植被層及土壤層碳素含量
灌木層不同器官的碳素含量介于370.92—408.17 g/kg之間,平均為387.26 g/kg,不同器官碳素含量表現為枝干>葉>根,變異系數介于6.08%—13.91%之間,草本層不同器官碳素含量表現為地上部分(416.27 g/kg)>地下部分(349.33 g/kg),平均為382.80 g/kg,變異系數介于12.23%—13.56%之間(表4),灌木層和草本層不同器官的碳素含量差異均不顯著(P>0.05)。不論灌木和草本,其地上部分碳素含量均高于地下部分。灌木層的碳素含量高于草本層,表現出隨植物個體高度或組織木質化程度的降低,其碳素含量相應減少的趨勢[6,13]。枯落物層碳素含量表現為未分解層(382.25 g/kg)>半分解層(373.98 g/kg),平均為378.11 g/kg,變異系數介于2.24%—2.63%之間(表4),各層的碳素含量差異不顯著(P>0.05)。
土壤不同層次碳素含量在17.08—58.23 g/kg之間,平均值為31.48 g/kg,變異系數介于16.82%—43.03%之間,0—20 cm土層的碳素含量最高,為58.23 g/kg,表現為隨土層深度的增加而減少的趨勢(表4),且差異極顯著(P< 0.01),說明森林土壤碳含量的分布有較強的表聚性。這主要是由于地表凋落物的積累和分解使得土壤表層有機質具有穩定而豐富的來源,同時植被根系對土壤表層有機碳的累積發揮了很強的作用[20-21]。多重比較結果表明:0—20 cm和20—40 cm、40—60 cm、60—80 cm土層的有機碳含量差異極顯著(P<0.01);20—40 cm和40—60 cm、60—80 cm土層有機碳含量差異顯著(P<0.05);40—60 cm和60—80 cm土壤有機碳含量差異不顯著(P>0.05)。

表4 林下植被層及土壤層碳素含量Table 4 Carbon Content in the Understory Vegetation and Soil
總的看來,雷公山常綠落葉闊葉混交林生態系統碳素含量表現為喬木層>灌木層>草本層>枯落物層>土壤層,差異極顯著(P<0.01)。植被層各層及枯落物層碳素含量均與土壤層碳素含量差異極顯著(P<0.01),喬木層與枯落物層碳含量差異顯著(P<0.05),而與其它層差異均不顯著(P>0.05),灌木層與草本層及枯落物層的碳含量差異均不顯著(P>0.05)。
3.2 生態系統碳密度及分配
3.2.1 生態系統碳密度及分配
通過表5計算得出生態系統碳密度為234.69 t/hm2,各層次碳密度表現為土壤層>喬木層>枯落物層>灌木層>草本層。土壤層的碳密度最大,占了生態系統碳密度的72.44%,喬木層的碳密度次之,占了生態系統碳密度的24.30%,枯落物層、灌木層和草本層的碳密度均較小,分別只占了生態系統碳密度的2.34%、0.77%和0.16%,說明土壤層和喬木層是影響生態系統碳密度最重要的兩個部分。同時,土壤層(0—80 cm)的碳密度是植被層和枯落物層的2.87和30.96陪,這說明土壤層是決定生態系統碳密度大小的關鍵作用層。
3.2.2 植被層與枯落物層碳密度及分配
從表5看出,喬木層碳密度為57.02 t/hm2,各器官的碳密度表現為干(29.89 t/hm2)>根(13.06 t/hm2)>枝(12.05 t/hm2)>葉(2.01 t/hm2)。在喬木層各器官中,樹干的碳密度最高,占了喬木層碳密度的52.43%,這充分說明,樹干在喬木層各器官中對碳的積累能力和貯存量均有重要的地位及意義。根與枝的碳密度次之,分別占了喬木層碳密度的22.90%和21.14%,葉的碳密度最小,僅占了3.53%。從表5還可以看出,灌木層的碳密度為1.81 t/hm2,表現為枝干(1.04 t/hm2)>根(0.47 t/hm2)>葉(0.30 t/hm2),各器官的碳密度占了灌木層碳密度的16.52%—57.52%,灌木層地上部分碳密度為1.34 t/hm2,占了灌木層碳密度的74.04%,是地下部分的2.85倍;草本層的碳密度為0.37 t/hm2,表現為地上部分(0.23 t/hm2)>地下部分(0.14 t/hm2),分別占了62.16%和37.84%,地上部分碳密度是地下部分的1.64倍;枯落物層的碳密度為5.48 t/hm2,表現為半分解層(2.98 t/hm2)>未分解層(2.50 t/hm2),分別占了54.43%和45.57%,半分解層的碳密度是未分解層的1.19倍。
3.2.3 土壤層碳密度及分配
土壤碳密度為170.00 t/hm2(0—80 cm),各層土壤碳密度在27.40—70.40 t/hm2之間,且隨著土壤深度增加而降低(表5),差異極顯著(P<0.01)。多重比較結果表明:0—20 cm和20—40 cm、40—60 cm、60—80 cm土層的碳密度差異極顯著(P<0.001);20—40 cm和40—60 cm土層的碳密度差異不顯著(P>0.05),而與60—80 cm土層的碳密度差異顯著(P<0.05);40—60 cm和60—80 cm土層差異不顯著(P>0.05)。表層(0—20 cm)土壤的碳密度占了土壤總碳密度(0—80 cm)的41.41%,其他各層占了土壤總碳密度(0—80 cm)的16.12%—24.00%(表5),表層(0—20 cm)土壤的碳密度顯著高于其他它各層,說明雷公山常綠落葉闊葉混交林土壤碳密度有很強的表聚性。這主要是由于植物根系集中分布在土壤表層,凋落物和腐殖層以及土壤微生物的分解對土壤有機碳的貢獻主要作用于地表,且隨土壤深度的增加而減弱,因而表層土壤的碳密度大[20-21]。
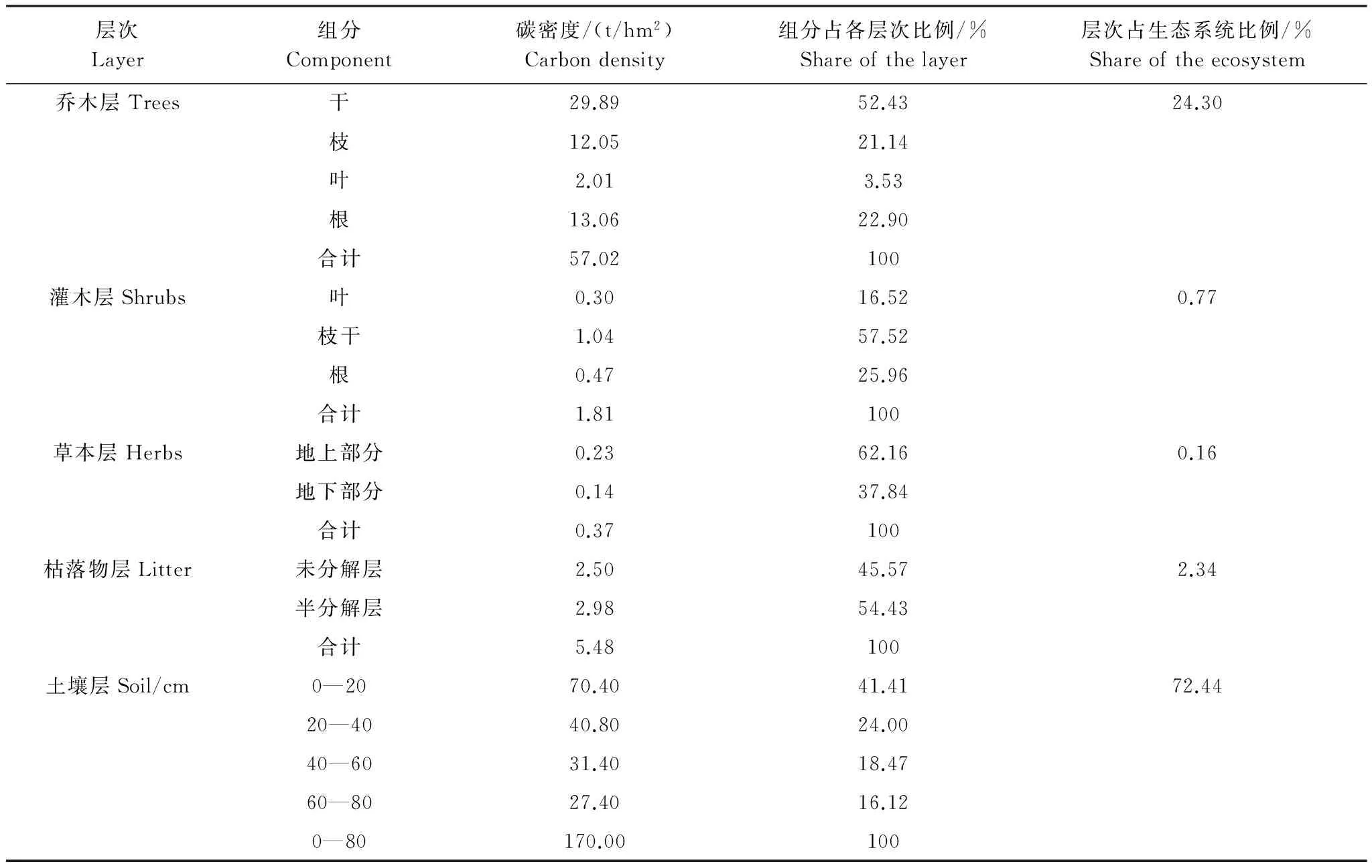
表5 生態系統碳密度及分配Table 5 Carbon Density and the Distribution in the Ecosystem
4 討論
4.1 碳素含量
本研究所測貴州東部常綠落葉闊葉混交林林下植被層的碳素含量低于喬木層碳素含量。這是由于喬木層有利于植物的光合作用,合成、積累較多的有機物,因而碳含量較高;而灌草層的光合作用減弱,有機物合成、積累少,碳含量相對較低。由于枯落物中的部分有機物被分解,使得處于半分解狀態的枯落物碳素含量低于未分解枯落物。而植被層平均碳素含量為40.02%,比國內外通常采用的碳素含量值(50%或45%)要低,因此在估算該區常綠落葉闊葉混交林植被層的碳儲量時,采用40%的轉換因子較為準確。
4.2 生態系統碳密度
本研究所得常綠落葉闊葉混交林生態系統碳密度約低于我國森林生態系統平均碳密度(258.83 t/hm2)[22],同時也低于江西九連山[12]與鼎湖山[8,14]亞熱帶常綠闊葉林地帶性植被的碳密度,但大于湖南鷹嘴界中亞熱帶常綠闊葉林[15]地帶性植被及馬尾松林[7,9-10]、樟樹林[11]、針葉混交林[6]、杉木林[13]等人工林的碳密度。說明研究區域、森林類型、樹種組成、立地條件等不同,導致生態系統碳儲量存在較大差異。植被層碳密度約高于我國森林植被平均碳貯量(57.07 t/hm2)[22],這主要是由于亞熱帶較好的水熱條件利于植被生物量的累積。但該植被層碳密度遠低于亞熱帶常綠闊葉林[8,12,14-15]植被層碳密度,說明該區植被層的碳匯能力較低,還有較大的碳匯潛力。植被層碳密度主要取決于植被層生物量的多少,而生物量又受林分年齡、密度和立地因子等方面的影響存在較大差異。喬木層碳密度占了植被層碳密度的96.99%,說明植被層碳密度主要取決于喬木層的生物量。但研究區地處中亞熱帶,水、熱條件豐富,加之屬國家級自然保護區,受人為破壞較少,常綠落葉闊葉混交林喬木層物種豐富,在計算喬木層的生物量時采用混合相對生長方程,其精度還有待進一步驗證和深入研究。灌草層和枯落物層的碳密度均極低,分別只占了生態系統碳密度的0.93%和2.34%,但林下植被在維持地力、保持水土等方面有著重要的作用[23],林下植被一旦遭受破壞,不僅造成植被碳的損失,而且加劇了地表徑流對表土層的沖刷,進而導致土壤碳流失。而枯落物層是土壤-植物系統碳循環的聯結庫,對森林生態系統的碳循環起到非常重要的作用。土壤層碳密度(0—80 cm)最高,是植被層碳密度的2.87倍,但土壤碳素的來源主要是地表枯枝落葉、土壤動物及植被根系的輸送,這說明了植被層及枯枝落葉層對土壤碳的貢獻也是非常大的。因此,加強森林保護,充分發揮植被在維持地力、保持水土等方面作用,減少地表徑流對表土層的沖刷,進而減少土壤碳散失,以維持和增加土壤碳貯量,對減緩大氣CO2濃度上升有著重大意義。
4.3 土壤碳密度
本研究所得土壤碳密度(0—80 cm)低于周玉榮等[22]報道的我國森林土壤碳密度平均水平,同時也低于貴州西部[20]華山松林、杉木林、柳杉林、樺木林和黔中喀斯特地區[21]常綠落葉混交林土壤層碳密度,但高于解憲麗等[24]報道的森林土壤碳密度全國平均水平,同時也高于鼎湖山亞熱帶常綠闊葉林[8,14]、黔中喀斯特地區灌木林和針闊混交林[21]、湖南鷹嘴界常綠闊葉林[15]及江西九連山常綠闊葉林[12]土壤層碳密度,說明森林土壤碳密度具有很強的空間異質性。這主要是由于森林類型的差異造成地表枯落物、地下微生物、植物根系和林下植被種類等的差異及各地區土壤發育的母巖、土壤類型,降雨量與林地管護等不同影響所致。表層(0—20 cm)土壤碳密度高于我國森林土壤(0—20 cm)土壤平均碳密度(42.4 t/hm2)[24],一方面主要是由于該區屬于亞熱帶地區,雨量充沛,森林植被結構復雜,林下植被豐富,凋落物現存量充足且分解轉化較快[20-21],另一方面雷公山屬于國家級保護區,森林管護較好,林地受破壞較少,表土層碳損失較少。表層(0—20 cm)土壤碳密度占了土壤層(0—80 cm)總碳密度的41.41%,顯著高于其它各層,而(0—40 cm)土壤碳密度占了土壤層(0—80 cm)總碳密度的65.41%,高于Detwiler[25]的熱帶和亞熱帶地區0—40 cm所貯存的碳占0—100 cm的比例,這充分說明該區森林土壤有機碳密度具有很強的表聚性,意味著森林土壤表層碳貯量貢獻較大,防止地表的水土流失,可有效保持土壤對碳的吸存。
5 結論
生態系統各組分碳素含量有較明顯的規律,表現為喬木層>灌木層>草本層>枯落物層>土壤層。植被層平均碳素含量為40.02%。土壤層碳素含量以0—20 cm土層最高,碳素含量隨土層深度的增加而減少。生態系統碳密度為234.69 t/hm2,各層次碳密度表現為土壤層>喬木層>枯枝落葉層>灌木層>草本層,分別占了生態系統碳密度的72.44%、24.30%、2.34%、0.77%和0.16%。植被層碳密度為58.79 t/hm2,占了生態系統碳密度的25.09%,植被層碳密度主要取決于喬木層的生物量。土壤層碳密度(0—80 cm)最高,林地土壤是一個極重要的碳庫,其中表層(0—20 cm)土壤碳密度顯著高于其它各層,土壤碳密度具有很強的表聚性。
[1] Tans P P. How can global warming be traced to CO2. Scientific American, 2006, 295(6): 124-124.
[2] 劉國華, 傅伯杰, 方精云. 中國森林碳動態及其對全球碳平衡的貢獻. 生態學報, 2000, 20(5): 733-740.
[3] Watson R T, Noble I R, Bolin R. Land Use, Land-Use Change and Forestry. Cambridge: Cambridge University Press, 2000: 30-30.
[4] Dixon R K, Brown S, Houghton R A, Solomon A M, Trexler M C, Wisniewski J. Carbon pools and flux of global forest ecosystem. Science, 1994, 263(5144): 185-190.
[5] Phillips O L, Malhi Y, Higuchi N, Laurance W F, Nunez P V, Vasquez R M, Laurance S G, Ferreira L V, Stern M, Brown S, Grace J. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science, 1998, 282(5388): 439-442.
[6] 康冰, 劉世榮, 張廣軍, 常建國, 溫遠光, 馬姜明, 郝文芳. 廣西大青山南亞熱帶馬尾松、杉木混交林生態系統碳素積累和分配特征. 生態學報, 2006, 26(5): 1320-1329.
[7] 巫濤, 彭重華, 田大倫, 閆文德. 長沙市區馬尾松人工林生態系統碳儲量及其空間分布. 生態學報, 2012, 32(13): 4034-4042.
[8] 王斌, 楊校生. 4種典型地帶性森林生態系統碳含量與碳密度比較. 湖南農業大學學報: 自然科學版, 2010, 36(4): 464-469, 473-473.
[9] 尉海東, 馬祥慶. 不同發育階段馬尾松人工林生態系統碳貯量研究. 西北農林科技大學學報: 自然科學版, 2007, 35(1): 171-174.
[10] 方運霆, 莫江明. 鼎湖山馬尾松林生態系統碳素分配和貯量的研究. 廣西植物, 2002, 22(4): 305-310.
[11] 雷丕鋒, 項文化, 田大倫, 方晰. 樟樹人工林生態系統碳素貯量與分布研究. 生態學雜志, 2004, 23(4): 25-30.
[12] 邸月寶, 王輝民, 馬澤清, 楊風亭, 張文江, 付曉莉, 梁躍龍, 周華. 亞熱帶森林生態系統不同重建方式下碳儲量及其分配格局. 科學通報, 2012, 57(17): 1553-1561.
[13] 康冰, 劉世榮, 蔡道雄, 盧立華. 南亞熱帶杉木生態系統生物量和碳素積累及其空間分布特征. 林業科學, 2009, 45(8): 147-153.
[14] 莫江明, 方運霆, 彭少麟, Brown S, 周國逸. 鼎湖山南亞熱帶常綠闊葉林碳素積累和分配特征. 生態學報, 2003, 23(10): 1970-1976.
[15] 宮超, 汪思龍, 曾掌權, 鄧仕堅, 陳建平, 龍康壽. 中亞熱帶常綠闊葉林不同演替階段碳儲量與格局特征. 生態學雜志, 2011, 30(9): 1935-1941.
[16] 馮宗煒, 王效科, 吳剛. 中國森林生態系統的生物量和生產力. 北京: 科學出版社, 1999.
[17] Fang J Y. Forest productivity in China and its response to global climate change. Acta Phytoecologica Sinica, 2000, 24(5): 513-517.
[18] Yang Y H, Mohammat A, Feng J M, Zhou R, Fang J Y. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry, 2007, 84(2): 131-141.
[19] 馬欽彥, 陳遐林, 王娟, 藺琛, 康峰峰, 曹文強, 馬志波, 李文宇. 華北主要森林類型建群種的含碳率分析. 北京林業大學學報, 2002, 24(5/6): 96-110.
[20] 丁訪軍, 高艷平, 周鳳嬌, 潘明亮, 吳鵬. 貴州西部4種林型土壤有機碳及其剖面分布特征. 生態環境學報, 2012, 21(1): 38-43.
[21] 丁訪軍, 潘忠松, 周鳳嬌, 吳鵬. 黔中喀斯特地區3種林型土壤有機碳含量及垂直分布特征. 水土保持學報, 2012, 26(1): 161-164.
[22] 周玉榮, 于振良, 趙士洞. 我國主要森林生態系統碳貯量和碳平衡. 植物生態學報, 2000, 24(5): 518-522.
[23] Liu S R, Li X M, Niu L M. The degradation of soil fertility in pure larch plantations in the northeastern part of China. Ecological Engineering, 1998, 10(1): 75-86.
[24] 解憲麗, 孫波, 周慧珍, 李忠佩. 不同植被下中國土壤有機碳的儲量與影響因子. 土壤學報, 2004, 41(5): 687-699.
[25] Detwiler R P. Land use change and the global carbon cycle: the role of tropical soils. Biogeochemistry, 1986, 2(1): 67-93.
Carbon accumulation and distribution characteristics of the evergreen broad-leaved and deciduous broad-leaved mixed forests in East Guizhou
DING Fangjun1,*, PAN Zhongsong2, WU Peng1, CUI Yingchun1, ZHOU Fengjiao1
1GuizhouForestryAcademy,Guiyang550005,China2SanduCountyForestryBureau,Sandu558100,China
This study was conducted to determine forest carbon content, density, and the characteristics of carbon distribution in the Evergreen Broad-leaved and Deciduous Broad-leaved mixed forest in Lei Gongshan Nature Reserve, Guizhou, China. Results indicated carbon content in the ecosystem varied significantly (P< 0.01) between trees, shrubs, herbs, litter and soil: 418.58 g/kg, 387.26 g/kg, 382.80 g/kg, 378.11 g/kg, and 31.48 g/kg, respectively. Among the various organs of a tree, the carbon content was distributed as follows: trunks > roots > leaves > branches. For shrubs, the carbon content was distributed as: stems > leaves > roots. For herbs, the above ground carbon content was higher than the below ground carbon content, while for all of them, the differences in carbon content among the different organs of a tree, shrub or herb was insignificant (P> 0.05). The average carbon content of all vegetation was 40.02%, when all types (trees, shrubs, and herbs) and all plant organs were considered. The carbon content in the litter of the un-decomposed layer was higher than in the semi-decomposed layer, though the difference was not significant (P> 0.05). The soil carbon content was highest in the soil at a depth of 0 to 20 cm and soil carbon content generally decreased inversely and significantly with soil depth (P< 0.01). The most of the carbon in the entire ecosystem (234.68 t/hm2) was divided between the soil (170.00 t/hm2) and trees (57.02 t/hm2) with the remainder being tied up in the litter (5.48 t/hm2), shrubs (1.81 t/hm2) and herbs (0.37 t/hm2), and accounting for 72.44%, 24.30%, 2.34%, 0.77% and 0.16% of the soil carbon of the entire ecosystem, respectively. The carbon density in the vegetation was 58.79 t/hm2accounting for 25.09% of that in the entire ecosystem. The carbon density among the various organs of a tree could be classified as: tree trunks > roots > branches > leaves and among them the carbon density of the tree trunk was the highest, accounting for 52.43% of the total. This indicated that tree trunks, among all the organs of a tree, were the major contributor not only to the tree′s capacity to accumulate carbon but also determined the quantity of carbon an ecosystem could store. The aboveground carbon density was 2.85 times higher than the underground density for shrubs and the aboveground carbon density was 1.64 times higher than the underground carbon density for the herbs. The carbon density in the soil ranged from 27.40 t/hm2to 70.40 t/hm2and it decreased with an increase in the soil depth. The carbon density in the topsoil (0 to 20 cm deep) was 70.40 t/hm2and was prominently higher than that of any other soil depth (P<0.001), and the carbon density in the topsoil contributed 41.41% of the entire soil carbon mass measured at depths of 0 to 80 cm, which indicated that carbon accumulated significantly near the topsoil. Therefore, the control of soil and water erosion could effectively maintain the soil′s capacity to sequestrate and fix soil organic carbon.
carbon content; carbon density; evergreen broad-leaved and deciduous broad-leaved mixed forests; Lei Gongshan Mountain
國家林業局公益專項子項目(201204101-5); 貴州省林業廳重大項目(黔林科合[2010]重大01)
2013-05-26;
日期:2014-04-25
10.5846/stxb201305261192
*通訊作者Corresponding author.E-mail: ding3920034@163.com
丁訪軍,潘忠松,吳鵬,崔迎春,周鳳嬌.貴州東部常綠落葉闊葉混交林碳素積累及其分配特征.生態學報,2015,35(6):1761-1768.
Ding F J, Pan Z S, Wu P, Cui Y C, Zhou F J.Carbon accumulation and distribution characteristics of the evergreen broad-leaved and deciduous broad-leaved mixed forests in East Guizhou.Acta Ecologica Sinica,2015,35(6):1761-1768.
