鰱對藻類攝食效應的室內模擬研究
2015-03-03 08:02:25璽王斌梁夏春香畢永紅陳胡征宇
水生生物學報 2015年5期
王 璽王斌梁夏春香畢永紅陳 磊,胡征宇
(1. 中國科學院水生生物研究所, 中國科學院藻類生物學重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049;3. 中國長江三峽集團公司樞紐運行管理局, 三峽工程魚類資源保護湖北省站點實驗室, 宜昌 443133)
鰱對藻類攝食效應的室內模擬研究
王 璽1,2王斌梁1,2夏春香1畢永紅1陳 磊1,2,3胡征宇1
(1. 中國科學院水生生物研究所, 中國科學院藻類生物學重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049;3. 中國長江三峽集團公司樞紐運行管理局, 三峽工程魚類資源保護湖北省站點實驗室, 宜昌 443133)
為研究鰱(Hypophthalmichthys molitrix)對藻類的攝食作用, 采用小環藻(Cyclotella)、小球藻(Chlorella)、微囊藻(Microcystis)和東湖原水開展了小規格魚種的養殖試驗, 對鰱攝食和排泄物進行了研究。結果顯示: 在3種純培養藻種投喂下鰱存活率從高到低依次是小環藻組>小球藻組>微囊藻組; 相應地, 鰱對藻的平均攝食率排序也是小環藻組>小球藻組>微囊藻組。在顯微鏡下觀察糞便發現大部分小環藻為空殼, 一部分小球藻外部形態結構不完整, 外部形態完整的小球藻則色澤暗淡、內部結構紊亂, 微囊藻沒有明顯變化; 3種藻類細胞的受損率分別為20.04%、7.13%和1.97%。葉綠素熒光活性顯示, 糞便中小環藻基本失去光合活性, 小球藻活性極顯著降低(P<0.01), 微囊藻活性明顯降低(P<0.05), 東湖原水處理組光合活性較對照極顯著降低(P<0.01)。對東湖水飼喂試驗組排泄物的檢測發現, 浮鞘絲藻Planktolyngya、魚腥藻Anabeana(有破損)、假魚腥藻Pseudoanabaena和顫藻Oscillatoria細胞受損嚴重; 空星藻Coelastrum和盤星藻Pediastrum細胞部分破損; 針桿藻Synedra、小環藻、橋彎藻Cymbella和舟形藻Navicula多為空殼。魚糞中平裂藻Merismopedia、微囊藻、浮鞘絲藻、魚腥藻、顫藻、卵囊藻Oocystis、柵藻Scenedesmus和衣藻Chlamydomonas在重新培養后恢復了生長。本研究顯示硅藻尤其是小環藻、針桿藻和舟形藻對鰱攝食最敏感; 柵藻、卵囊藻和平裂藻對攝食壓力不敏感。研究表明鰱捕食對藻類群落結構有明顯的改變效應, 利用鰱的濾食進行藻類群落調節和水華防控具有可行性。
藻類; 浮游植物群落; 種群; 鰱; 攝食率
當前富營養化已經成為影響水體環境質量的主要難題[1]。近十幾年來, 我國幾乎所有的江湖河海和局部地下水或多或少遭受氮磷污染[2], 富營養化趨勢顯著。水體的富營養化會引發藻類暴發性增殖(水華), 水中溶解氧降低, 水生生物大量死亡, 水生生態系統紊亂, 進而影響城市供水、景觀旅游和水產品養殖業等。治理水體富營養化根本上是要解決其中的氮磷等營養鹽負荷過高的問題[3], 采取的方法可分為物理方法、化學方法和生物方法等。物理方法相對安全但運行成本高、難以高效除藻; 化學方法中使用的化學試劑效果顯著但易造成二次污染;生物方法是指利用食物鏈傳遞過程使得營養鹽得到降解和轉化, 該凈化技術具有環保低耗的特色。20世紀 70年代 Shapiro等[4]提出生物操縱的概念, 80年代有學者提出了利用魚類下行作用模型及生物操縱來防治湖泊富營養化[5]; 生物操縱的核心內容是利用浮游動物控制藻類[5], 我國科學家對魚類與水域環境間的關系也有大量研究[6,7], 謝平[8]提出的利用鰱鳙控制藍藻技術稱之為非經典生物操縱。
非經典生物操縱在湖泊富營養化的治理上已取得一定效果。謝平等[9]在對武漢東湖的圍格實驗表明鰱鳙魚對微囊藻有顯著地抑制作用。并且在巢湖、滇池、千島湖和山東東周水庫等水污染治理中, 這項應用也展現了較好的研究成果[10—12]。但另有研究發現, 在較淺的水域中, 隨著濾食性魚類的攝食活動和生理代謝的增加, 促進了氮磷的釋放反而有利于浮游植物的增長[13]; 并且還有研究認為魚類對大型浮游植物的攝取導致浮游植物整體趨于小型化,微型浮游生物比重顯著增加[14]。研究認為利用鰱鳙控制水華的成功與其放養的時間和密度有重大關系[14]。法國的 Domaizon等[15]發現低密度放養白鰱時葉綠素含量較低, 隨著密度增大葉綠素含量也增高。在波蘭的Wamiak湖中放養鰱的密度為30—90 g/m3時,浮游植物生物量和藍藻比例明顯減小[16]。劉建康等[9]提出東湖放養鰱的密度在46—50 g/m3以上, 能夠顯著控制微囊藻的增殖。而 Domaizon等[15]實驗表明鰱的放養閾值為26 g/m3, 超過則反助于小型藻的暴發性滋長。另外浮游植物的沉降速率很低, 在淺水中由于魚類的活動其沉降更慢; 并且魚類也會排泄氮磷等營養物質到水體中, 這可能引起水體初級生產的增加[17]。
利用濾食性魚類控制藻類的技術實行結果具有較大差異, 與實驗條件、放養密度及具體水域實際情況有關。為深入探明生物操縱的作用, 本研究直接從濾食性魚類本身的攝食效應出發, 考察濾食性魚類對藻類種群與群落的處置效果, 確認生物操縱對修復生態環境的實際效果。
1 材料與方法
1.1 試驗材料
試驗所用鰱魚種購自湖北省鄂州市保安湖扁擔塘東風農場。試驗前魚苗先在室溫22℃下暫養3d。之后選擇體重相當的魚種, 暫養在曝氣自來水的水族缸(130 L)中饑餓處理2d, 待魚腸清空并稱重作為研究材料。試驗所用藻類(微囊藻、小環藻和小球藻)均來自于中國科學院水生生物研究所藻種庫, 試驗時選擇穩定生長期的藻細胞來投喂魚類。
1.2 魚種存活率及排泄物觀察
試驗前離心收集藻細胞, 加入到10 L的玻璃水族箱中調節水體中的藻類密度, 定期鏡檢確定水族箱的藻類細胞密度; 每個處理組設置 3個重復和 3個對照組。微囊藻(Microcystis aeruginosa)密度為928.5×105cells/L, 小環藻(Cyclotella)密度為726.3× 105cells/L, 小球藻(Chlorella)密度組為822.8×105cells/L。3種處理組的水族箱里分別飼養 50條魚種(體重0.254—0.272 g, 平均體長為2.6 cm)通氣培養。3個對照組細胞密度與試驗組相同, 全部對照組未放入魚種。溫度控制在22℃左右, 光照強度45 μmol/(m2·s)。每天記錄魚種存活數量, 10d后從水族箱移出并稱重。收集魚糞, 4000 r/min下離心。稱量0.1 g蒸餾水稀釋到1 mL。在顯微鏡下統計完整的藻細胞數與非完整的藻細胞數。
1.3 藻細胞的攝食率
試驗前離心收集藻細胞, 加入到 8 L的玻璃水族箱中調節密度, 設置3個重復組和2個對照組。微囊藻組的密度設定為1.857×106cells/L, 小環藻組的密度為 1.495×106cells/L, 小球藻組的為 1.525× 106cells/L。3種藻及其重復組的水族箱里放養 20條饑餓處理的魚種(體重 0.225—0.232 g, 平均體長2.6 cm)通氣培養。對照組的藻液密度與試驗組相同,未放入魚種。溫度控制在 22℃左右, 光照強度低于2 μmol/(m2·s)。5h后將魚苗移出, 在顯微鏡下統計藻細胞的密度。
攝食率的計算公式為: I=V×{Ceo–[Ceo×(Cco–Cct)/Cco]–Cet}/W·t
其中V是水的體積, W是魚的濕重, Ceo和Cet分別為試驗組藻細胞的起始和終止時的密度; Cco和 Cct是對照組藻細胞的起始和終止時的密度[18]。
1.4 東湖水喂養魚種的攝食及排泄
以東湖水注入水族槽中(測定藻細胞密度), 每組10 L (共三組)均等放入 20條小魚種[總重(4.5± 0.021) g]通氣培養。試驗期間同等投喂魚體重 2%的飼料。另設 3組水族箱不加魚種的東湖水用作對照,試驗溫度22℃左右, 光照強度10 μmol/(m2·s)。每隔3天測定葉綠素含量。20d后測定水體浮游植物的密度。另設一組10 L東湖水放養10條小魚種, 不投喂其他飼料, 每3天收集魚糞用顯微鏡下觀察鏡檢藻類。
1.5 魚糞飼喂和再培養
設兩組10 L曝氣自來水放養的魚種(10條), 試驗前同樣饑餓處理 2d。每天按魚體重的 5%僅投喂前東湖水試驗收集的糞便(濕重)。1周后收集糞便再觀察。
將收集的魚糞稱重加入到BG11和D1培養液,置于光照培養箱, 光強50 μmol/(m2·s), 12D︰12L光暗周期、22℃的條件下再培養 2周, 觀察藻類活性的恢復情況。
1.6 魚糞的光合活性檢測
在清水槽中收集魚糞離心, 用蒸餾水等倍稀釋。分別吸取3 mL樣品利用 WATER-PAM(德國)檢測最大光化學效率Fv/Fm。
1.7 數據分析
利用數據采用Excel 2013、SPSS 17.0 和origin 9軟件進行統計分析和制圖, 使用 t-Test、ANOVA統計方法對試驗處理組進行統計意義檢驗。
2 結果
2.1 純培養微藻飼喂鰱的現存量
由圖1可見, 微囊藻試驗組的魚種20d內死亡數量最多, 最終平均現存量為 14條, 平均體重(0.256±0.005) g。小球藻試驗組的魚種20d后存活數平均為18條, 體重平均值(0.26±0.004) g。小環藻試驗組的魚苗20d內死亡數最低, 最終平均現存量19條, 平均體重(0.262±0.006) g。經檢驗, 微囊藻組的魚種存活數顯著低于其他兩組(P<0.05)。并且小環藻組的存活數顯著高于小球藻組(P<0.05)。
2.2 純培養微藻飼喂鰱的排泄物分析
由表1可知不同藻喂養魚種排泄物中藻細胞的平均密度。其中小環藻密度高達2.66×106cells/g, 小球藻次之, 微囊藻密度相對最低, 僅有1.68×106cells/g。受損細胞比率的檢測結果顯示, 小環藻在排泄物中的完整細胞數最少, 微囊藻細胞多保持完整形態。由鰱對小環藻、小球藻和微囊藻在水溫22℃左右的平均攝食率可知小環藻的攝食率最高。
2.3 東湖水飼養鰱的藻細胞密度
東湖水試驗組在 20d后, 密度顯著變化。表 2是以東湖水飼養小魚, 對照組和試驗組的細胞密度變化情況。一共鑒定出硅藻7屬。試驗組的硅藻細胞總密度相比于對照組減少 9.88×105cells/L; 其中小環藻細胞密度最高, 試驗組與對照組相比有顯著降低(P<0.01); 針桿藻細胞密度與對照組無顯著差異; 舟形藻屬的密度顯著減少(P<0.01)。藍藻鑒定到9屬, 結果顯示藍藻細胞顯著減少(P<0.01); 浮鞘絲藻、藍纖維藻、顫藻和微囊藻的細胞密度降低; 與對照組相比, 藍纖維藻屬、微囊藻屬、澤絲藻屬和顫藻屬的數量有顯著性減小(P<0.01)。綠藻有 8屬,柵藻、空星藻、盤星藻和卵囊藻的細胞密度與對照組相比有顯著降低(P<0.01)。
根據體積轉化法推算了試驗組的生物量變化量(表2)。生物量顯著變化的有針桿藻、浮鞘絲藻、藍纖維藻、柵藻和盤星藻(P<0.05)。
2.4 鰱排泄物中藻類鑒定與再培養
表3顯示的是魚糞中觀察到的藻細胞及其存在狀態, 17種藻中浮鞘絲藻的數量高達86.4%, 其次是平裂藻和假魚腥藻的數量相對較多。平裂藻多保持完整, 假魚腥藻大多斷裂。其他細胞受損率高的藻類還有魚腥藻、顫藻和盤星藻。硅藻的受損細胞數最高(空殼細胞計入受損細胞數)。
表4是以鰱魚糞投喂饑餓處理的魚種后魚糞收集觀察和再培養的結果。一共鑒定到13種藻類。另外糞便再培養后仍觀察到10種藻。柵藻和平裂藻的細胞數最高。硅藻暫未發現。魚糞再培養的生長情況與投喂前收集觀察的結果一致。
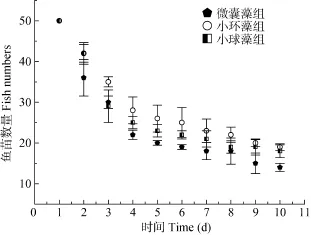
圖1 不同微藻飼喂試驗組的魚種存活數Fig. 1 The standing crop of three microalgae feeding groups

表1 純培養微藻飼喂鰱的排泄物藻類分析Tab. 1 The cell density and digestion in the feces of silver carp fed with different algal species
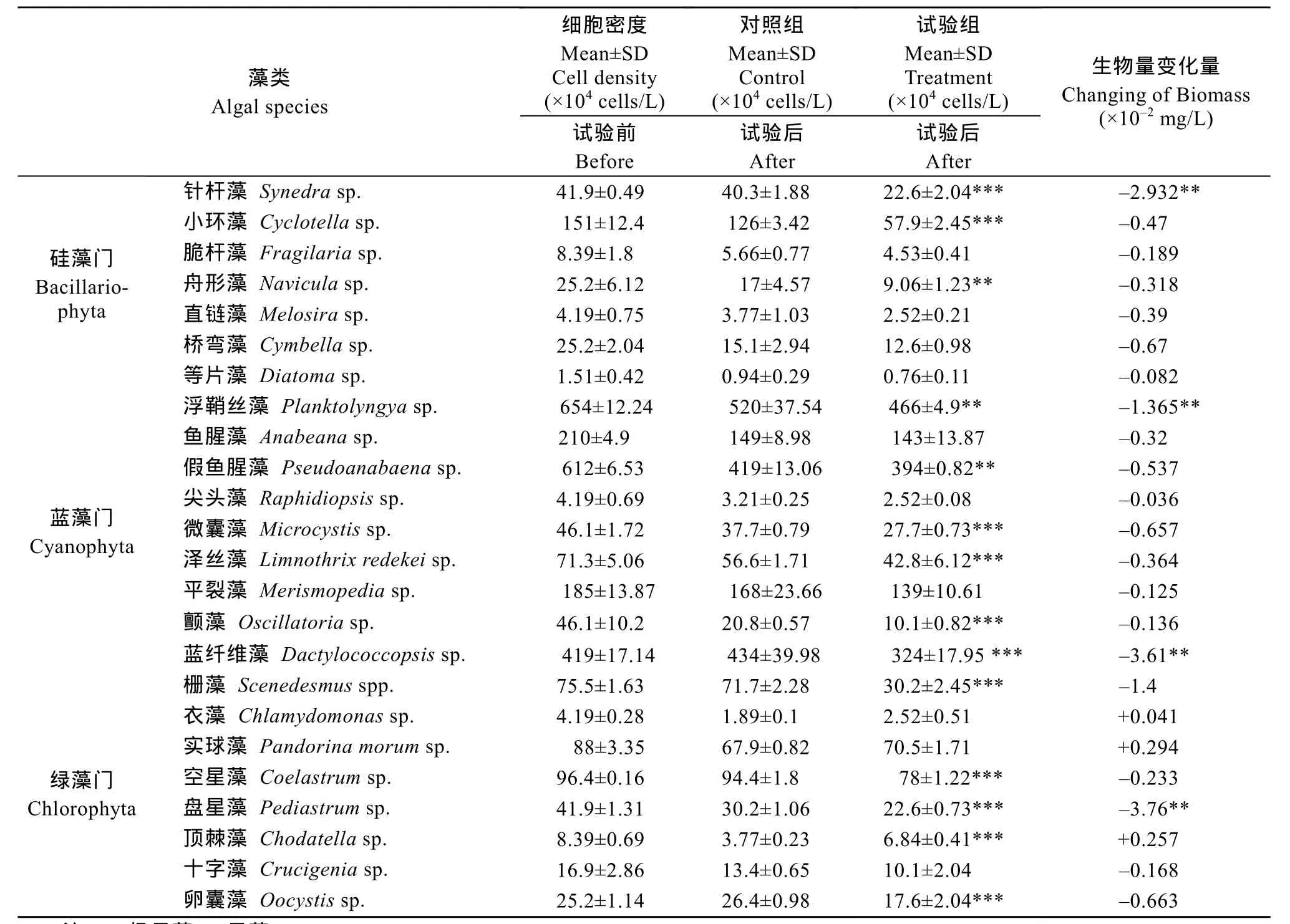
表2 東湖水飼養鰱的藻細胞密度變化情況Tab. 2 The change in cells density in the aquarium microcosms
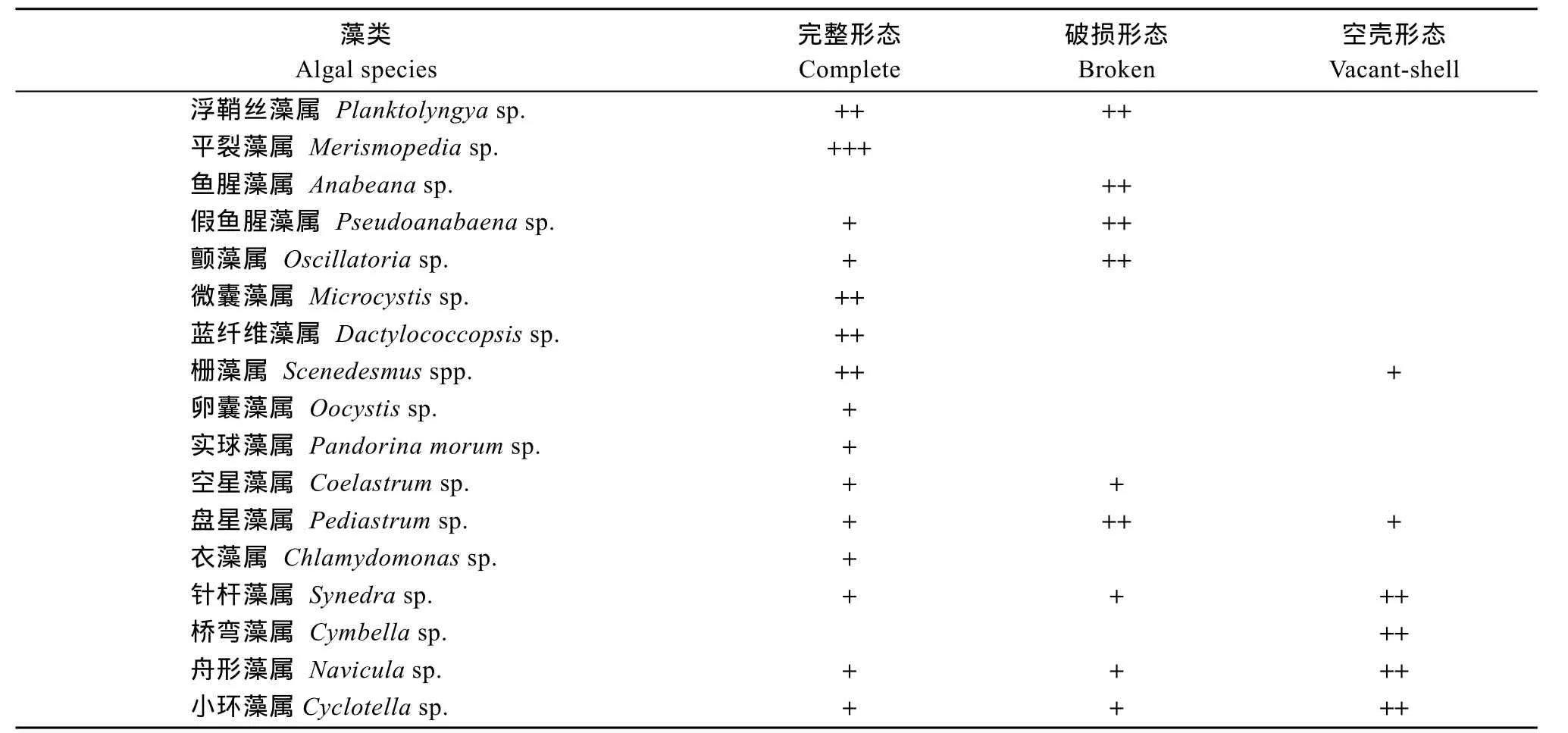
表3 東湖水試驗組魚糞中的藻類組成Tab. 3 The species and shapes of the algae in sliver carp feces
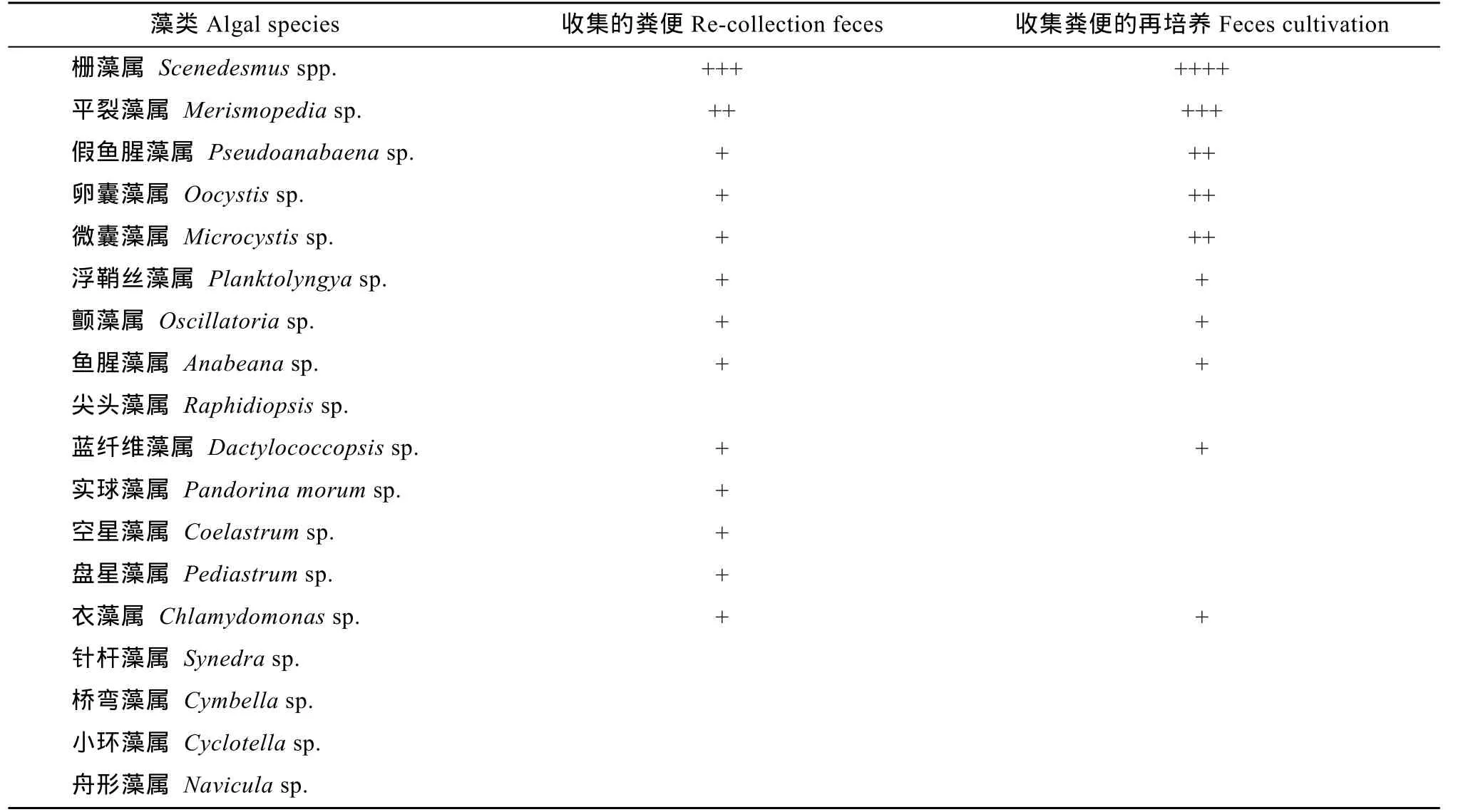
表4 魚糞投喂饑餓魚種后的糞便收集與再培養情況Tab. 4 The re-collection and re-cultivation of feces discharged by the hungry silver carp fed with feces
采用東湖水飼養鰱的水體浮游植物密度和鰱排泄物的結果顯示, 水體中的密度變化顯著的有: 小環藻、舟形藻、針桿藻、微囊藻、藍纖維藻、澤絲藻、顫藻、假魚腥藻、浮鞘絲藻、柵藻、空星藻、盤星藻和卵囊藻。糞便中完全受損的藻類有浮鞘絲藻、魚腥藻(有破損)、假魚腥藻和顫藻; 盤星藻破損的較多, 空星藻有小部分破損; 針桿藻、小環藻、橋彎藻和舟形藻多為空殼。糞便中的柵藻、平裂藻、微囊藻、浮鞘絲藻、魚腥藻、顫藻、卵囊藻和衣藻在重新培養后恢復了生長。由此可見, 硅藻, 尤其是小環藻、針桿藻和舟形藻對濾食性魚類的攝食最為敏感。
2.5 藻類光合系統活性
圖2為不同微藻以及東湖水投喂鰱前后及其魚糞再培養液的最大光化學效率(Fv/Fm)。可見, 糞便中的小環藻基本失去活性, 小球藻活性極顯著降低(P<0.01), 微囊藻活性明顯降低(P<0.05), 東湖原水處理組糞便光合活性較對照極顯著降低(P<0.01)。經過再培養的魚糞, 其光合活性不同程度地恢復; 微囊藻的活性甚至高于投喂前的對照, 小球藻和東湖原水處理組均有較大幅度的活性恢復, 但小環藻的活性恢復能力最弱。
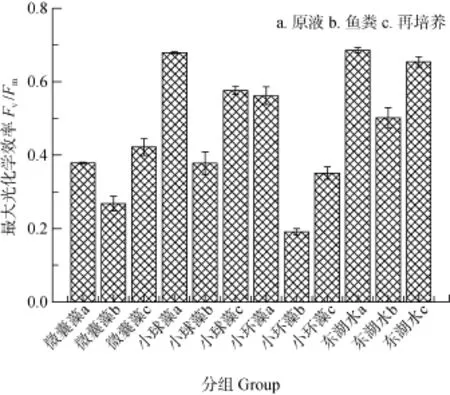
圖2 不同處理組飼養鰱的葉綠素熒光活性變化Fig. 2 Variation of chlorophyll fluorescence in different treatment groups
3 討論
史為良等[17]的網箱實驗結果顯示, 放養魚密度大的處理組其優勢種多為柵藻、小球藻和藍纖維藻等; 朗宇鵬等[19]的研究表明鰱對藍藻生長具有很好的控制力。本試驗發現鰱能夠有效濾食部分藍藻。雖然排出的藻類仍有部分活性, 但需要良好的環境條件才能恢復生長; 在自然環境條件下, 魚糞會吸引浮游動物和原生動物等攝食分解, 進一步增加了其活性恢復和種群復壯的難度。在本研究中投放魚種的年齡小, 由于試驗時間較短, 盡管對藻類物種的影響有限, 但也顯示了其對藍藻生物量的顯著改變作用。從這個角度看, 濾食性魚類能夠攝食利用部分藍藻的結論是正確的。
有研究認為攝食百分比與水體中浮游植物的密度具有一定相關性, 通常密度越大則攝食效果越明顯[19]。在本研究的結果中, 投喂鰱的純培養微藻細胞密度相當, 但攝食卻存在顯著的差異, 攝食結果并沒有顯示與藻類密度的相關性, 鰱對硅藻的攝食消化情況好于綠藻和藍藻。對純培養微藻飼喂鰱的排泄物鏡檢發現, 微囊藻細胞受損率最小, 這與之前的研究者所認為的被膜藻類不易被鰱消化[20,21]的結果相同。盡管小環藻的存活數量最高但卻大部分是完全空殼細胞, 周潔和林峰[22]曾提到鰱最易消化的硅藻是異端硅藻和舟形藻等, 倪達書[21]列舉了硅藻門細胞構成濾食性魚類的主要食物。在顯微鏡下常可見硅質殼破損并且正在釋放色素的半消化細胞。在東湖原水的飼養試驗中, 盡管藍藻浮鞘絲藻細胞密度最大, 但經過飼養試驗后, 盤星藻、藍纖維藻和針桿藻的生物量變化最大, 也就是它們被攝食利用的最多, 由此可見, 鰱對硅藻的消化利用能力顯著地高于藍藻; 濾食性魚類的攝食與水環境中的細胞密度并沒有明顯的相關性, 而是具有物種的特異性。
鰱排泄物觀察結果與周潔和林峰[22]的結果相近。從攝食數量和攝食后形態變化情況來看, 純培養的微藻飼養結果顯示了硅藻對攝食最敏感, 藍藻對攝食最不敏感; 東湖原水飼養的結果顯示, 對鰱攝食敏感的藍藻有假魚腥藻、浮鞘絲藻和顫藻; 敏感的綠藻有盤星藻。而在濾食性魚類強大的攝食壓力下, 以微囊藻為代表的物種難以被攝食壓力破壞而保存了較高的種群數量。卵囊藻的消化情況與水體密度變化并非完全一致。其存在形態較為完整并且在適當的培養條件可以恢復生長, 但水體中的密度卻有顯著降低, 可能與試驗光照及溫度控制有關。
大量研究結果均支持濾食性魚類對藻類增殖的抑制作用[8,9,23—25]。另一些研究則認為, 濾食性魚類會攝食浮游動物從而減弱了浮游動物對浮游植物的攝食壓力, 間接導致浮游植物生物量增加, 并且可能促進小型藻類暴發生長[26—28]。從本研究結果來看,鰱對藻類的攝食能力得到驗證, 僅在0.1 mL魚糞中就觀察到 104個數量級以上的藻細胞被攝食。以東湖原水飼喂魚種發現, 浮游植物細胞密度和生物量明顯降低。有人發現引入鰱不能完全控制浮游植物,但會使浮游植物組成發生顯著改變, 細胞數量變化并不大[29]。出現這樣的情況可能與具體水域有關,試驗所采用的水體以及藻類群落的差異可能導致結果略有不同。
1987年有學者發現鰱糞便中的微囊藻在特定培養基上可以生長[30]。Kolmakov等[31]測定了鯽排泄物中活性藻類的生物量及光合效率, 發現微囊藻的生長與光合活性都顯著增加。王銀平等[32]曾對雜食性魚類排泄物進行了藻類光能活性研究; 結果表明魚類攝食對微囊藻生長及葉綠素熒光參數均有顯著影響, 并且光合活性和生長活性經過再培養后會逐漸恢復。本試驗同樣發現攝食排泄后各組的 Fv/Fm值顯著變化, 糞便中的微囊藻光合系統活性顯著降低, 但再培養后的活性竟然高于對照, 據此推測適度的攝食壓力對微囊藻具有刺激作用, 也從另一個角度說明了微囊藻具有較強的攝食抗性, 對其種群調控具有相當的難度; 其他藻類和東湖水光合活性也有不同程度的恢復, 但均難以恢復到對照的水平,尤其是硅藻, 光合活性恢復較困難。由此可見, 經濾食性魚類攝食后的不同藻類具有不同的恢復能力,這種恢復能力的差異最終將導致水體浮游植物種群動態的顯著差別。
[1] ILEC/Lake Biwa Research Institute. Survey of the state of the World’s Lakes [C]. Nairobi, International Lake Environment Committee, Otsu and United Nations Environment Programme. 1993, Volumes I-IV
[2] Lu K H, Yan W J, Su S A. Environmental and ecological engineering on control and remediation of eutrophicated waterbodies: by using ameliorated alum plasma and fishes to control blue- green blooms of qiaodun reservoir [J]. Acta Scientiae Circumstantiae, 2002, 22(6): 732—737 [陸開宏,晏維金, 蘇尚安. 富營養化水體治理與修復的環境生態工程—利用明礬漿和魚類控制橋墩水庫藍藻水華. 環境科學學報, 2002, 22(6): 732—737]
[3] Wang Y, Liu L S, Fang Y D, et al. The research advances about biomanipulation method control of eutrophicated lake [J]. Progress in Natural Science, 2009, 19(12): 1296—1301 [王瑜, 劉錄三, 方玉東, 等. 生物操縱方法調控湖泊富營養化研究進展. 自然科學進展, 2009, 19(12): 1296—1301]
[4] Shapiro J, Lamarra V, Lynch M. Biomanipulation: an ecosystem approach to lake restoration [J]. Proceedings ofthe Symposium on Water, 1975, 21(6): 85—96
[5] Liu E S. Analysis on biomanipulation, non-traditional biomanipulation and discussion of the termeasures of biomanipulation application in waters [J]. Journal of Lake Sciences, 2010, 22(3): 307—314 [劉恩生. 生物操縱與非經典生物操縱的應用分析及對策探討. 湖泊科學, 2010, 22(3): 307—314]
[6] Liu J K. Ecological Research on East Lake (I) [M]. Beijing: Science Press. 1990, 292—371 [劉健康主編. 東湖生態學研究(一). 北京: 科學出版社. 1990, 292—371]
[7] Liu J K. Ecological research on East Lake (II) [M]. Beijing: Science Press. 1995, 360—415 [劉健康主編. 東湖生態學研究(二). 北京: 科學出版社. 1995, 360—415]
[8] Xie P. Silver Carp and Bighead with Algae Blooms Control [M]. Beijing: Science Press. 2003, 116 [謝平. 鰱、鳙與藻類水華控制. 北京: 科學出版社. 2003,116]
[9] Liu J K, Xie P. Direct control of Microcystis bloom through the use of planktivorous carp-closure experiments and lake fishery practice [J]. Ecologic Science, 2003, 22(3): 193—196 [劉建康, 謝平. 用鰱鳙直接控制微囊藻水華的圍隔試驗和湖泊實踐. 生態科學, 2003, 22(3): 193—196]
[10] Li C Y, Cui Y. The application of biomanipulation in preventing and curing the eutrophication of cultivated water [J]. Marne Fisheries Research, 2002, 23(1): 71—75 [李春雁,崔毅. 生物操縱法對養殖水體富營養化防治的探討. 海洋水產研究, 2002, 23(1): 71—75]
[11] Chen L S, Hong R H, He G X. The application of non-traditional biomanipulation in controlling the blooms [J]. Fishery Modernization, 2006, 3: 45—46 [陳來生, 洪榮華,何光喜. 運用非經典生物操縱技術治理水華. 漁業現代化, 2006, 3: 45—46]
[12] Li Q, Li D S, Xiong B X, et al. Influence of silver carp (Hypophthalmicthys molitrix C et V) on plankton community in reservoir enclosures [J]. Acta Ecologica Sinica, 1993, 13: 30—37 [李琪, 李德尚, 熊邦喜, 等. 放養鰱魚(Hypophthalmicthys molitrix C et V) 對水庫圍隔浮游生物群落的影響. 生態學報, 1993, 13: 30—37]
[13] Gu X H, Liu G Y. Impact of sliver carp and bighead carp in fish ponds on plankton [J]. Journal of Ecology and Rural Environment, 1996, 12(1): 6—10, 41 [谷孝鴻, 劉桂英. 濾食性鰱鳙魚對池塘浮游生物的影響. 農村生態環境, 1996, 12(1): 6—10, 41]
[14] Yan Y H, Zhong C H, Deng C G. Progress of non-traditional bio-manipulation in the controlling of eutrophication [J]. Journal of Anhui Agricultural Sciences, 2007, 35(12): 3459—3460 [閆玉華, 鐘成華, 鄧春光. 非經典生物操縱修復富營養化的研究進展. 安徽農業科學, 2007, 35(12): 3459—3460
[15] Domizon I, Devaux J. Impact of moderate silver carp biomass gradient on zooplankton communities in a eutrophic reservoir. Consequences for the use of silver carp in biomanipulatin [J]. Academie Des Sciences Paris Comptes Rendus Serie Generale La Vie Des Sciences, 1999, 322(7): 621—628
[16] Kajak Z, Rybak J L. Influence of the planktonivorous fish, Hypoph-thalmichthys molitrix, on the plankton and benihos of eutrophic lake Poi. Arch [J]. Archiv Fur Hydrobiologie, 1975, 22: 301—310
[17] Shi W L, Jin W H, Wang D Q. Effect of stocking of sliver carp and bighead carp on the eutrophication in waters [J]. Journal of Dalian Fisheries College, 1989, 4(3): 11—24 [史為良, 金文洪, 王東強. 放養鰱鳙對水體富營養化的影響.大連水產學院學報,1989, 4(3): 11—24]
[18] Lu K H, Jin C H, Dong S L, et al. Feeding and control of blue-green algal blooms by tilapia (Oreochromis niloticus) [J]. Hydrobiologica, 2006, 568: 111—120
[19] Lang Y P, Zhu L, Liu C G, et al. Bio- control of silver carp on phytoplankton in freshwater [J]. Journal of Agro-Environment Science, 2006, 25(Supplement): 683—686 [郎宇鵬, 朱琳, 劉春光, 等. 鰱魚對淡水浮游植物的抑制作用研究.農業環境科學學報, 2006, 25(增刊): 683—686]
[20] He Z H, Li Y H. On the food of the silver carp [J]. Acta Hydrobiologica Sinica, 1975, 5(4): 541—548 [何志輝, 李永函. 論白鰱的食物問題. 水生生物學報, 1975, 5(4): 541—548]
[21] Lin D S, Jiang Y Z. On the food of the silver carp and bighead [J]. Current Zoology, 1954, 6(1): 59—71 [倪達書,蔣燮治. 花鰱和白鰱的食料問題. 動物學報, 1954, 6(1): 59—71]
[22] Zhou J, Lin F. The feeding habit of silver carp and bighead and their digestion of algae [J]. Acta Hydrobiologica Sinica, 1990, 14(2): 170—177 [周潔, 林峰. 鰱、鳙的食性及其對藻類的消化利用. 水生生物學報, 1990, 14(2): 170—177]
[23] Liu X J, Xie P, Wang S M, et al. Studies on the causes of resumption and disappearance of Microcystis bloom in donghu lake: stocking experiments in enclosures [J]. Journal of Lake Sciences, 1994, 6(3): 245—256 [劉學君, 謝平, 王少梅, 等. 武漢東湖微囊藻水華形成及消失原因的探討一圍圈放養實驗. 湖泊科學, 1994, 6(3): 245—256]
[24] Wang H Z, Liu Y D, Xiao B D, et al. Ecological meaning and bloom controlling of different density of silver carp followed with potamogeton crispus in enclosures [J]. Acta Hydrobiologica Sinica, 2004, 28(2): 141—147 [王海珍, 劉永定, 肖邦定, 等. 圍隔中鱗鰱和菹草控藻效果及其生態學意義. 水生生物學報, 2004, 28(2): 141—147]
[25] Stewart T W, Winer J G, Lowe R L. An experimental analysis of crayfish (Orconectes rustieus) effeets on a Dreissena dominated benthic macroinvertebrate community in western lake Erie [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(4): 1040—1050
[26] Chen S L, Liu X F, Hua L. The role of silver carp and bighead in the cycling of nitrogen a nd phosphorus in the east lake ecosystem [J]. Acta Hydrobiologica Sinica, 1991, 15(1): 8—26 [陳少蓮, 劉肖芳, 華俐. 鰱、鳙在東湖生態系統的氮、磷循環中的作用. 水生生物學報, 1991, 15(1): 8—26]
[27] Reinertsen H. Effcet of fish removal on to the limonitic ecosystem of an eutrophic lake [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1990, 47: 166—173
[28] Shapiro J, Wright D I. Lake restoration by biomanipulation: Round Lake, Minnesota, the first two years [J]. Freshwater Biology, 1984, 14: 371—383
[29] Pogozhev P I, Gerasimova T N. The effect of zooplankton on microalgae blooming and water eutrophication [J]. Water Resources, 2001, 7/8: 420—427
[30] Kharitonova N N, Sirenko L A, Stetsenko L I. Viability of planktonic algae passed intestines of herbivorous fish (case of bighead) [J]. Rybnoie khoziaistvo, 1987, 41: 1—80
[31] Kolmakov V I, Gladyshev M I. Growth and potential photosynthesis of cyanobacteria are stimulated by viable gut passage in crucian carp [J]. Aquatic Ecology, 2003, 37: 237—242
[32] Wang Y P, Zhao Y, Zeng Q F, et al. Changes in the photosynthetic activity of Microcystis colonies after gut passage through omnivorous fish [J]. China Environmental Science, 2013, 33(3): 524—529 [王銀平, 趙勇, 曾慶飛,等. 雜食性魚類排泄物中藻類光能活性研究. 中國環境科學, 2013, 33(3): 524—529]
THE PREDATOR EFFECTS OF SLIVER CARP ON DIFFERENT ALGAL SPECIES INSIDE AQUARIUM
WANG Xi1,2, WANG Bin-Liang1,2, XIA Chun-Xiang1, BI Yong-Hong1, CHEN Lei1,2,3and HU Zheng-Yu1
(1. Key Laboratory of Algal Biology, Institute of Hydrobiology, Chinese Academy of Sciences, 430072 Wuhan, China; 2. University of Chinese Academy of Sciences, 100049 Beijing, China; 3. Hubei Key Laboratory of Three Gorges Project for Conservation of Fish, Three Gorges Construction and Operation Management Department, China Three Gorges Corporation, Yichang 443133, China)
To study the ingestion and digestion of different algal species by sliver carp, we conducted feeding and feces experiments in aquarium microcosms. Sliver carps were fed with three species of algae (Cyclotella, Chlorella and Microcystis aeruginosa), and the ingestion rates per fish weight for the three species were 2.70×104, 1.99×104and 1.06×104cells/(g·h) respectively. In feces, the ratios of the complete cells to the imperfect cells were 20.04%, 7.13% and 1.97% for the three species respectively. Microcosm experiments were conducted in aquariums filled with water from the Donghu Lake for two weeks. Each aquarium had 20 fish [total weight (4.5±0.021) g] inside, and there were three replicates in one experiment. We found that after 20 days the cell density in the aquariums was significantly reduced compared to the control. Planktolyngbya accounted for 87% of the phytoplankton density in feces, and Merismopedia sp., Pseudoanabaena sp., Oscillatoriaceae sp. and Microcystis sp. accounted for 11%, but Raphidiopsis sp., Scenedesmus spp., Coelastrum sp. only formed a very small portion. The morphology of Oscillatoria sp., Spirulina sp., and Pediastrum sp. appeared to be incomplete, while Synedra sp., Navicula sp. and Cyclotella sp. had a nearly vacant-shell shape. We collected the feces to feed the hungry sliver carps and did not observe any Pediastrum sp., Cyclotella sp. or Synedra sp. in the new discharge. However, we still found Planktolyngbya sp., Anabeana sp., Pseudoanabaena sp., Merismopedia sp., Microcystis sp., Raphidiopsis sp., Oscillatoriaceae sp., Microcystis sp., Scenedesmus spp., Oocystis sp., Pandorina morum sp., Coelastrum sp., Chlamydomonas sp. and Cymbella sp. After a 10d culture of the feces in the medium, we observed the revival of Planktolyngbya sp., Oscillatoriaceae sp., Merismopedia sp., Anabeana sp., Pseudoanabaena sp., Microcystis sp., Scenedesmus spp., Chlamydomonas sp. and Oocystis sp. The level of chlorophyll a decreased slightly at first and then increased in fish groups. Our study suggested that sliver carp could filter phytoplankton with the size of 2—50 μm, and that sliver carp might mainly feed on diatoms, of which the ingestion and digestion was easier than that of green algae and blue-green algae. The results also indicated that Cyclotella sp. and Navicula sp. were sensitive to the predator stress. Therefore silver carp could be an efficient tool to regulate the algal population and the phytoplankton communities.
Algae; Phytoplankton community; Population; Sliver carp; Ingestion rate
Q178.1
A
1000-3207(2015)05-0940-08
10.7541/2015.123
2015-03-02;
2015-04-28
中國長江三峽集團公司科研項目(CT-12-08-01)資助
王璽( 1991— ), 女, 安徽六安人; 碩士研究生; 研究方向為藻類生態學; E-mail: 516738801@qq.com
畢永紅, E-mail: biyh@ihb.ac.cn
