三峽庫區(qū)5個鰱群體遺傳變異的微衛(wèi)星分析
2015-03-03 08:02:06龐美霞俞小牧童金茍
水生生物學報 2015年5期
關鍵詞:分析
龐美霞俞小牧童金茍
(1. 中國科學院水生生物研究所, 淡水生態(tài)與生物技術國家重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049)
三峽庫區(qū)5個鰱群體遺傳變異的微衛(wèi)星分析
龐美霞1,2俞小牧1童金茍1
(1. 中國科學院水生生物研究所, 淡水生態(tài)與生物技術國家重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049)
研究利用 10個高度多態(tài)的微衛(wèi)星標記對三峽水庫秭歸、巫山、云陽、忠縣、木洞等 5個庫區(qū)鰱(Hypophthalmichthys molitrix)的野生群體進行了遺傳多樣性分析。檢測到161個等位基因, 群體共有等位基因84個, 每個微衛(wèi)星位點的等位基因數(shù)7—29不等。平均觀測雜合度Ho為0.784—0.846, 平均期望雜合度He為0.828—0.847, 平均多態(tài)信息含量PIC為0.797—0.817。Fst值為–0.001—0.009, 表明5個鰱群體間沒有遺傳分化。Hardy-Weinberg平衡檢驗表明巫山、云陽、木洞群體在一些位點上偏離遺傳平衡。Bottleneck分析顯示長江三峽庫區(qū)江段的鰱群體可能在歷史上經(jīng)歷了遺傳瓶頸。5個群體間的遺傳相似系數(shù)為0.891—0.950, 遺傳距離為 0.050—0.115, 根據(jù) Nei’s遺傳距離所繪制的聚類圖, 表明鰱群體間的遺傳距離與其地理距離基本一致。貝葉斯分析結(jié)果也證實三峽庫區(qū) 5個鰱群體可視為一個類群。盡管沒有檢測到遺傳分化, 數(shù)據(jù)清晰地表明三峽庫區(qū)的鰱群體仍有很高的遺傳多樣性, 研究結(jié)果為三峽地區(qū)和長江上游的鰱種質(zhì)資源保護和種群評估提供了參考。
鰱; 微衛(wèi)星; 野生群體; 遺傳多樣性; 三峽水庫
鰱(Hypophthalmichthys molitrix)是我國“四大家魚”之一, 廣泛分布于我國長江、黑龍江、珠江等各大水系, 是我國重要的經(jīng)濟魚類。長江流域是我國鰱天然資源的主要產(chǎn)地, 其養(yǎng)殖性能明顯優(yōu)于珠江、黑龍江等其他水系[1]。然而由于長期的過度捕撈、水體污染、水利建設等一系列人為因素導致近年來長江水系自然環(huán)境劇烈變化[2], 從而引起包括鰱在內(nèi)的“四大家魚”資源量的嚴重衰退[3]。群體遺傳分析是合理評估魚類種群遺傳多樣性和種群分化的有效手段。過去已有一些針對長江流域鰱野生群體遺傳多樣性分析的報道[4—8], 但主要集中在長江中下游地區(qū), 有關長江中上游地區(qū)的相關報道少見[6]。長江三峽水庫已經(jīng)建成, 針對這一河流型水庫中鰱野生種群的遺傳結(jié)構分析方面的資料還非常缺乏,這種狀況不利于魚類資源的評估和合理利用。
微衛(wèi)星標記是一種以1—6 bp的核苷酸序列為核心序列的簡單重復序列[9], 由于其具有共顯性遺傳、數(shù)量多且分布廣泛均勻、多態(tài)信息含量高及便于檢測等諸多優(yōu)點, 已被廣泛應用于群體遺傳分析[10,11]。本研究采用本實驗室開發(fā)的4—5核苷酸重復的10個多態(tài)性鰱微衛(wèi)星標記對來自三峽水庫從庫首到庫尾的5個庫區(qū)(秭歸、巫山、云陽、忠縣、木洞)的鰱樣本進行分析, 旨在掌握三峽庫區(qū)以及長江上游鰱野生群體的遺傳多樣性和遺傳結(jié)構現(xiàn)狀, 為長江流域“四大家魚”種質(zhì)遺傳資源的保護和利用提供理論依據(jù)。
1 材料與方法
1.1 試驗材料及DNA提取
本實驗所分析的 5個野生鰱群體樣本采集于2012—2013年, 采樣點位于湖北省秭歸(ZG)、重慶市巫山(WS)、云陽(YY)、忠縣(ZX)和木洞鎮(zhèn)(MD),樣本總數(shù)227尾(表1、圖1)。剪取每尾魚的少量鰭條用無水酒精保存, 基因組 DNA采用經(jīng)典的酚-氯仿法[12]提取。
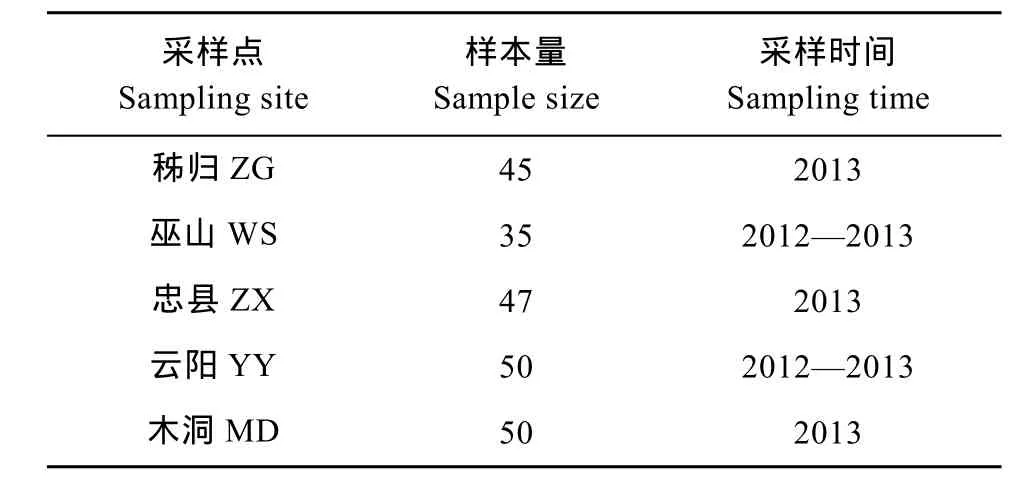
表1 三峽庫區(qū)5個鰱群體樣本的基本信息Tab. 1 The basic information of five silver carp populations in the Three Gorges Reservoir
1.2 標記選取及PCR擴增
在本研究中使用的標記均來自于本實驗室開發(fā)的鰱微衛(wèi)星標記。為了能夠清晰地分型便于數(shù)據(jù)統(tǒng)計, 本文所選取的 10個鰱微衛(wèi)星標記[13]均是 4—5堿基重復的高度多態(tài)性位點(表2)。根據(jù)Li-Cor 4300 DNA 分析系統(tǒng)的檢測要求, 在合成每一對微衛(wèi)星引物時, 將一段19堿基( 5′-CACGACGTTGTAAAA CGAC-3′)的接頭序列添加到每一個微衛(wèi)星位點的正向引物序列 5′端[14]。在PCR進行擴增時, 帶有接頭序列的微衛(wèi)星上游引物既可與目的片段結(jié)合, 又可與 5′端帶有熒光接頭的 IR-M13F引物[M13 Forward(-29) IRDye 700 Primer 或M13 Forward (-29) IRDye 800 Primer] (LI-COR Biosciences, Lincoln, NE, USA) 結(jié)合, 從而把IR-M13F 帶入到擴增反應中, 使得擴增的目的片段末端帶有熒光標記從而通過 4300 DNA 分析儀檢測并成像[15]。 本文所用的微衛(wèi)星引物和熒光標記引物均由上海生工生物工程有限公司合成。
PCR反應總體積為12.5 μL, 反應混合物包含以下組分: 30—50 ng模板DNA, 0.4 U Taq DNA聚合酶, 1.25 μL 10×PCR buffer, 0.4 μL dNTP (2.5 mmol/L), 0.4 μL正反向混合引物(各 2.5 μmol/L), 0.3 μL IR-M13F引物, 10 μL滅菌去離子水。PCR擴增條件如下: 94℃預變性5min; 擴增35個循環(huán), 每個循環(huán)94°C變性35s, 48—60℃退火35s(退火溫度見表2), 72℃延伸40s; 72℃終延伸10min。
1.3 微衛(wèi)星分型及數(shù)據(jù)統(tǒng)計
在 PCR擴增結(jié)束后, 在含有擴增產(chǎn)物的薄壁管中加入 7 μL變性劑( 98%去離子甲酰胺, pH 8.0, 10 mmol/L EDTA, 0.25%溴酚藍), 95℃變性10min后立即將置于冰水中冷卻。變性產(chǎn)物在 Li-Cor 4300 DNA分析儀中進行電泳檢測并自動成像。MS-TOOLS軟件用于計算觀測雜合度(Ho)、期望雜合度(He)、等位基因數(shù)(Na)等多樣性參數(shù), 以及計算多態(tài)信息含量(PIC)。PopGene 32軟件計算各個群體間的Nei’s遺傳距離[16]和相似系數(shù)。并根據(jù)Nei’s遺傳距離用 MEGA 4軟件[17]對 5個鰱群體進行聚類分析。Arlequin 3.1軟件[18]計算各群體間的分化系數(shù) Fst, 同時用于進行各群體的哈迪-溫伯格平衡(Hardy-Weinberg equilibrium)檢驗、連鎖不平衡(Linkage disequilibrium)檢驗。進行多重比較時, 概率的顯著性水平需要進行 Bonferroni校正[19]。利用分子變異分析(AMOVA)方法[20]計算群體遺傳結(jié)構。根據(jù)各位點等位基因頻率, 使用BOTTLENECK3.4軟件[21]進行瓶頸效應分析。基于無限等位基因模型(Infinite allele model, IAM), 采用符號檢驗(Sign test)分析雜合度過剩情況, 依此推測群體數(shù)量動態(tài)變化。利用Structure 2.2[22]軟件中的貝葉斯方法混合模型來構建群體遺傳結(jié)構圖。
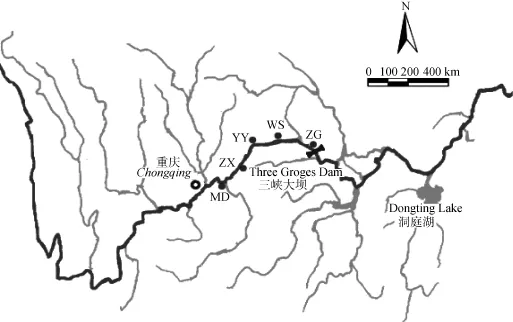
圖1 三峽水庫鰱群體采樣點分布示意圖Fig. 1 Distribution of sampling sites for silver carps in the Three Gorges Reservoir
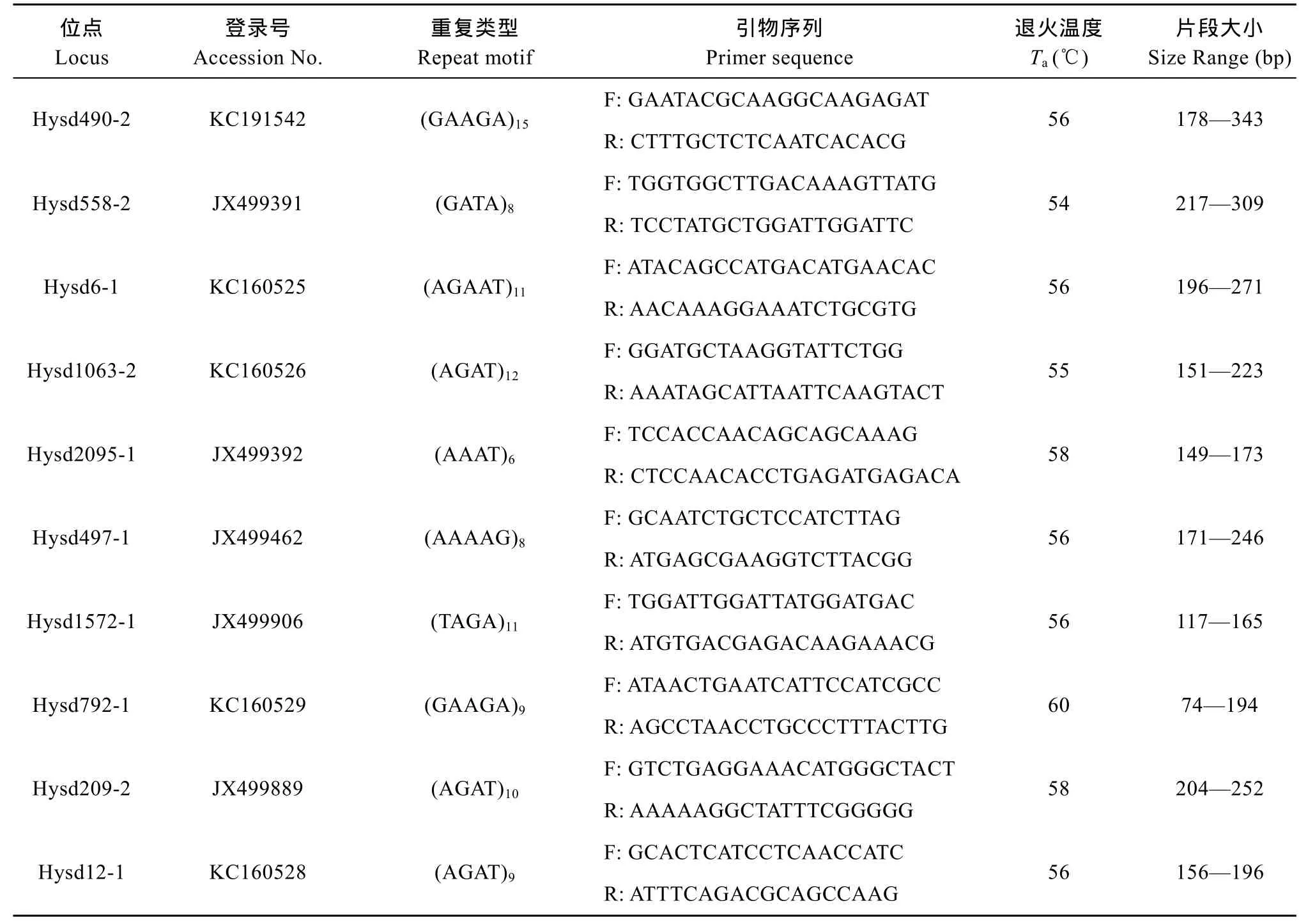
表2 本研究所用鰱微衛(wèi)星標記的基本信息Tab. 2 The microsatellite markers of silver carps used in this study
2 結(jié)果
2.1 鰱各群體的遺傳多樣性特征及群體動態(tài)分析
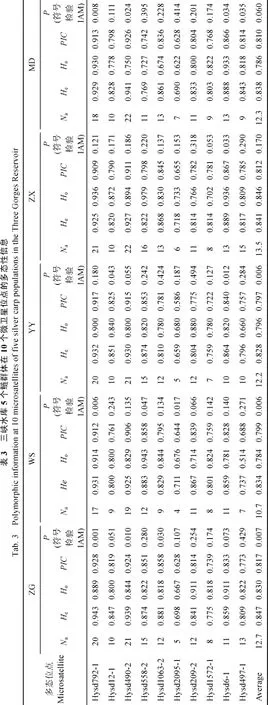
5個鰱群體各位點的等位基因數(shù)(Na)、期望雜合度(He)、觀測雜合度(Ho)以及多態(tài)信息含量(PIC)統(tǒng)計結(jié)果見表3。在10個微衛(wèi)星位點上共檢測到161個等位基因, 其中ZG、WS、YY、ZX、MD群體分別測出127、107、122、135和123個等位基因, 5個鰱群體共有等位基因84個, 并且ZG、WS、YY、ZX、MD群體分別有4、3、5、5和2個特殊等位基因。每個微衛(wèi)星位點的等位基因數(shù)7—29不等。ZX群體的平均Na最高(13.5), 巫山群體最低(10.7)。平均 Ho、He和 PIC分別為 0.784—0.846、0.828—0.847和0.797—0.817, 其中WS群體Ho最低, ZX群體Ho最高; YY群體He和PIC在所有群體中均最低,而ZG群體He和PIC在所有群體中均最高。
對各個鰱群體進行哈迪-溫伯格平衡檢驗并用Bonferroni校正后發(fā)現(xiàn), WS群體在 Hysd792-1、Hysd490-2、Hysd2095-1、Hysd209-2、Hysd497-1 位點, YY群體在 Hysd2095-1位點, MD群體在Hysd490-2、Hysd1063-2 位點, 在 0.05置信水平上偏離哈迪-溫伯格平衡。而連鎖不平衡分析結(jié)果顯示,所有10個微衛(wèi)星標記在5個鰱群體均不存在連鎖不平衡。遺傳瓶頸效應分析結(jié)果(表3)顯示, 在IAM模型下, 除 ZX群體外其他群體均表現(xiàn)出雜合過度(P<0.05), 表明這些群體近期可能經(jīng)歷了瓶頸效應。
2.2 群體間的遺傳分化
5個鰱群體的遺傳分化分析結(jié)果表明, 群體間的Fst值(表4)為–0.001—0.009, 群體兩兩間Fst差異均不顯著(P>0.05), 表明他們沒有遺傳分化。AMOVA分析結(jié)果顯示群體間的遺傳變異僅占0.47% , 而群體內(nèi)的遺傳變異占99.53% 。對群體內(nèi)的各個體進行混合模型分析結(jié)果顯示, 當K值(理論群)取2時, 5個鰱群體遺傳結(jié)構高度一致; 當K值取3—5時, WS群體少部分個體遺傳結(jié)構相對獨立, 5個鰱群體遺傳結(jié)構基本一致。鰱各個群體間的遺傳距離和相似系數(shù)如表 5所示。Nei氏遺傳距離和相似系數(shù)分別為 0.050—0.115、0.891—0.950, 以 ZX群體和MD群體之間的遺傳距離最小, 而MD群體與WS群體之間的遺傳距離最大。
依據(jù)計算出來的Nei氏遺傳距離(表5), 5個野生鰱群體的聚類分析結(jié)果(圖2)顯示, 鄰接樹中共有兩大分枝, 其中ZG和WS聚為一枝, 其余3個群體聚為另一大分枝。這個聚類結(jié)果基本與群體間的地理距離關系一致。
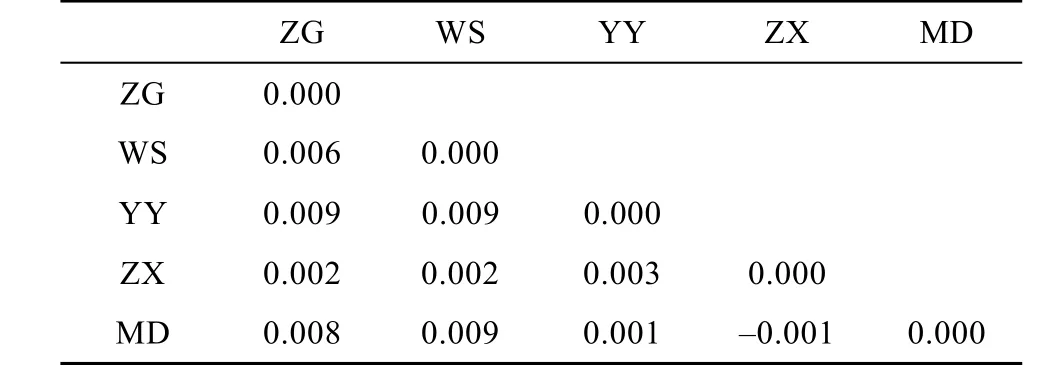
表4 三峽庫區(qū)5個鰱群體兩兩間Fst值Tab. 4 Pairwise Fstvalues of five silver carp populations in the Three Gorges Reservoir
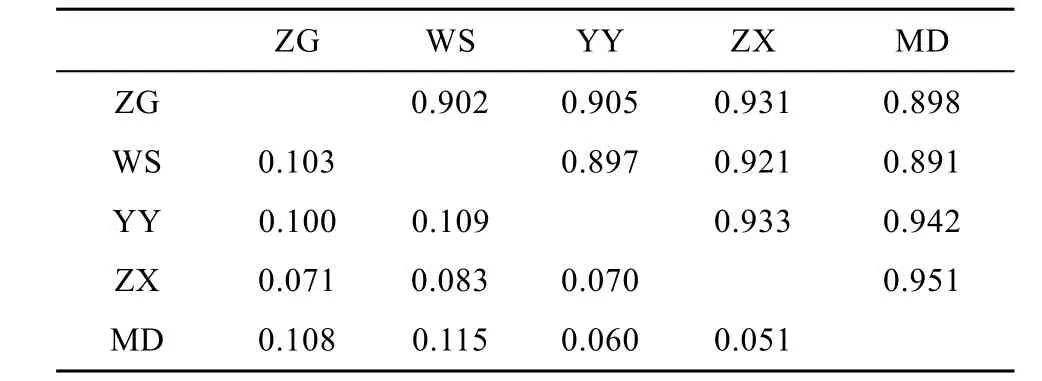
表5 五個鰱群體兩兩間Nei遺傳一致性(對角線上方)和遺傳距離(對角線下方)Tab. 5 Nei’s genetic identity (above diagonal) and genetic distance (below diagonal) among five silver carp populations
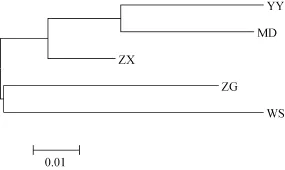
圖2 依據(jù)Nei氏遺傳距離構建的三峽庫區(qū)鰱5個鰱群體間的鄰接樹Fig. 2 Neighbour-Joining tree of five silver carp populations in the Three Gorges Reservoir of the Yangtze River based on Nei’s genetic distance
3 討論
3.1 微衛(wèi)星標記的適用性
微衛(wèi)星標記具有共顯性遺傳、數(shù)量多且在基因組中分布廣泛、多態(tài)信息含量高及便于檢測等諸多特點, 是物種種群遺傳分析的最常用分子遺傳標記。微衛(wèi)星標記的多態(tài)性來源于微衛(wèi)星核心重復序列重復次數(shù)的變化[23], 與其他諸如RAPD、AFLP、SNP等標記相比, 微衛(wèi)星標記具有更高的分辨率。本研究采用的10個鰱微衛(wèi)星標記, 均是4—5堿基重復序列。有研究表明, 相比于二核苷酸重復的微衛(wèi)星標記, 多核苷酸重復的微衛(wèi)星標記的擴增穩(wěn)定性更高, 能忠實地反映DNA序列遺傳本質(zhì)[24,25]。在長江三峽庫區(qū) 5個野生鰱群體中, 每個微衛(wèi)星位點的等位基因數(shù)從7—29不等(表3), 表明這10個微衛(wèi)星均為較高多態(tài)性的座位。平均多態(tài)信息含量(PIC)是衡量等位基因片段多態(tài)性的理想指標, 當PIC>0.5時該位點表現(xiàn)為高度多態(tài), 當 0.25<PIC<0.5時該位點表現(xiàn)為中度多態(tài), 當PIC<0.25 時該位點表現(xiàn)為多態(tài)性低下[26]。在本研究中使用的10個微衛(wèi)星標記中, 除了 Hysd2095-1為中度多態(tài)性以外,其余標記的PIC值均大于0.7為高度多態(tài)(表3), 這說明本文所選用的微衛(wèi)星位點是鰱野生群體遺傳多樣性研究的理想分子遺傳標記。
3.2 鰱群體遺傳多樣性及群體動態(tài)分析
基因雜合度表示群體在某座位雜合子的比例。觀測雜合度(Ho)和期望雜合度(He)是反映群體遺傳變異的兩個重要指標。雜合度越大變異越大, 對環(huán)境適應能力越強[7]。本研究發(fā)現(xiàn) 5個鰱野生群體的平均等位基因為 10.7—13.5, 平均有效等位基因數(shù)Ne為 6.6—7.5, 平均觀測雜合度Ho為0.784—0.846,平均期望雜合度He為 0.828—0.847, 平均多態(tài)信息含量PIC為0.797—0.817(表3), 這說明三峽庫區(qū)鰱野生群體遺傳多樣性仍在很高水平, 對長江“四大家魚”種質(zhì)資源保護和恢復有積極意義。關于長江流域鰱群體遺傳多樣性分析, 迄今已有一些報道。朱曉東等[5]利用30個微衛(wèi)星標記對長江中下游5個鰱群體進行遺傳多樣性分析, 發(fā)現(xiàn)這 5個鰱群體的平均有效等位基因數(shù) Ne為 2.444—2.633, 平均觀察雜合度 Ho為 0.323—0.351,平均期望雜合度 He為0.442—0.470,平均多態(tài)信息含量 PIC 為 0.406—0.428。王長忠等[6]對長江中上游萬州和監(jiān)利鰱群體的遺傳多樣性分析顯示, 兩個鰱群體的平均有效等位基因數(shù)Ne分別為4.107和3.395;平均觀測雜合度Ho為0.834和0.775,平均期望雜合度He為0.713和0.623; 兩群體平均多態(tài)信息含量 PIC為 0.617。由于本研究采用的微衛(wèi)星標記絕大部分為高度多態(tài)性標記, 而且本文采用更加靈敏的熒光標記基因分型手段, 因此Ne比過去的報道要高的多。本研究中Ho、He、PIC等遺傳多樣性參數(shù)也遠高于朱曉東等關于長江中下游 5個鰱群體的報道, 而與王長忠等[6]對于長江中上游2個鰱群體的報道較為接近。 這可能在一定程度上表明, 長江中上游相較于長江中下游鰱群體的遺傳多樣性更為豐富。
本文在所檢測的微衛(wèi)星位點中, WS群體在5個位點、YY群體在1個位點、MD群體在2個位點偏離哈迪-溫伯格平衡。可見WS群體內(nèi)有較多位點的基因型頻率發(fā)生了較大改變, 而這種改變可能與多種因素有關, 例如突變、選擇、遷移和雜交等。當一個群體的基因位點出現(xiàn)雜合度過剩及不足的概率大致均等時, 表明這個群體處于突變-漂移平衡的狀態(tài)[27]。而當群體經(jīng)歷瓶頸效應則會表現(xiàn)出雜合度過剩的現(xiàn)象, 因此雜合度過剩可以衡量群體數(shù)量下降的瞬間效應[28]。本研究采用 IAM 模型符號檢驗發(fā)現(xiàn), ZG、WS、YY、MD 4個群體顯示出顯著雜合子過剩(表 3), 說明以上 4 個群體可能在近期經(jīng)歷了不同程度的瓶頸效應, 群體數(shù)量發(fā)生一定程度的下降。在長江三峽水庫建成運行的條件下, 庫區(qū)過度人工捕撈等因素尤其應該引起重視, 以免對鰱等“四大家魚”的遺傳資源造成不可逆的破壞。
3.3 鰱群體遺傳分化
基因分化系數(shù)是衡量群體間等位基因的遺傳分化程度的重要參數(shù)。當 Fst為 0—0.05 時, 群體間遺傳分化很弱; 0.05—0.15 時, 遺傳分化中等; 0.15—0.25時, 遺傳分化較大;大于0.25時, 表示群體間分化極大[29]。在 本研究中各群體間的 Fst值均小于0.05(表4), 表明三峽庫區(qū)5個鰱野生群體間幾乎沒有分化。AMOVA分析結(jié)果也支持這一結(jié)果: 群體間的遺傳變異只占0.47% , 而群體內(nèi)的遺傳變異高達 99.53% , 表明三峽庫區(qū)鰱的遺傳變異基本來自于群體內(nèi)部。利用 structure軟件中混合模型, 根據(jù)設定的分群數(shù)(K)進行群體遺傳結(jié)構研究, K=2—5時均顯示 5個鰱群體間遺傳分化不明顯, 基本可劃分為一個類群。而ZG、WS、YY、ZX、MD 5個群體各有4、3、5、5和2個特殊等位基因, 這些特殊基因可能為 5個群體間將來出現(xiàn)遺傳分化提供方向。過去的一些研究報道認為長江流域中各鰱群體或江段之間存在一定程度的遺傳分化。如王淞等[7]對4 個鰱群體進行了mtDNA D-loop的PCR-RFLP分析, 結(jié)果顯示長江干流的監(jiān)利和瑞昌兩個群體與長沙及寧河群體間有一定的分化; 朱曉東等[5]對長江中下游5個鰱群體進行了微衛(wèi)星分析發(fā)現(xiàn)5個鰱群體間存在中等程度的分化; 而王長忠等[6]對長江中上游萬州和監(jiān)利兩個鰱群體進行了微衛(wèi)星分析認為萬州和監(jiān)利鰱群體間存在顯著遺傳分化, 應隸屬于不同的種群。本研究認為三峽庫區(qū)的5個鰱群體間沒有遺傳分化, 可以看成一個天然群體。這一結(jié)果也與郭穩(wěn)杰[13]對長江中上游9個鰱群體的微衛(wèi)星分析結(jié)果類似。郭穩(wěn)杰[13]認為除湖口外, 其他群體間只呈現(xiàn)出較弱的群體分化。本文所研究的三峽庫區(qū)的 5個采樣點, 盡管有少數(shù)采樣點在長江支流,但整體來說這5個群體之間的地理距離都不是太大,目前也沒有明顯的地理隔離, 故我們認為這些水域的鰱群體之間進行了較頻繁的基因交流。
根據(jù)Nei氏遺傳距離(表5)所繪制的5個野生鰱群體的鄰接樹(圖2)顯示, YY與MD的遺傳距離最小, 而與 WS最大, 表明群體間的遺傳距離與地理距離之間有一定的關聯(lián)性。但也并非完全地理位置越遠的群體間遺傳分化程度越大, 例如 YY在地理位置上與ZX和WS更近, 結(jié)果顯示YY與MD的聚類關系反而更近, 這可能是由于 YY部分位點偏離哈迪溫伯格平衡, 導致部分基因型缺失, 從而在一定程度上影響了其遺傳多樣性; 而YY與WS的聚類關系反而最遠, 可能是由于 WS獨特的地理環(huán)境導致WS群體與其他群體間的基因交流受到一定影響; 也有可能是由于 WS群體受到較大的捕撈壓力導致基因型頻率受到影響。
遺傳多樣性是種內(nèi)個體之間或群體內(nèi)不同個體的遺傳變異總和, 是生物適應環(huán)境的基礎, 同時種內(nèi)的遺傳變異程度又決定其進化的潛勢[30]。對一個物種來說, 遺傳多樣性越高, 則其對生存環(huán)境變化所產(chǎn)生的適應能力越強, 進化的潛力也越大。由于過去對長江上游尤其是三峽庫區(qū)鰱野生群體的遺傳多樣性和遺傳結(jié)構的現(xiàn)狀了解不多, 人類對該地區(qū)鰱、鳙等經(jīng)濟魚類的種群動態(tài)和種質(zhì)資源評價方面長期缺乏群體遺傳學分析數(shù)據(jù)。本研究的結(jié)果證實長江三峽庫區(qū)范圍內(nèi)的鰱基本可看作一個天然群體,也在一定程度上給出了該地區(qū)鰱群體仍有較高的遺傳多樣性的事實。本研究分析結(jié)果也提示三峽地區(qū)可能多個鰱群體曾發(fā)生過遺傳瓶頸效事件, 這也應引起魚類保護生物學工作者的注意。本文對于長江上游以及整個長江流域的鰱野生資源保護或增殖放流效果評估等研究有積極作用。
[1] Li S F, Wu L Z, Wang Q, et al. Changjiang River, Zhujiang River, Heilongjiang River Silver Carp, Bighead Carp, Grass Carp Germplasm Resource Research [M]. Shanghai: Shanghai Science & Technology Press. 1990, 25—50 [李思發(fā), 吳力釗,王強, 等. 長江、珠江、黑龍江鰱、鳙、草魚種質(zhì)資源研究.上海: 上海科學技術出版社. 1990, 25—50]
[2] Zhang M Y, Xu D P, Liu K, et al. Analysis of genetic diversity among silver the artificially released populations of carps in the lower reach of Yangtze River with microsatellite markers [J]. Acta Agriculturae Universitatis Jiangxiensis, 2012, 34(1): 141—146 [張敏瑩, 徐東坡, 劉凱, 等. 長江下游放流鰱群體遺傳多樣性的微衛(wèi)星標記分析. 江西農(nóng)業(yè)大學學報, 2012, 34 (1): 141—146]
[3] Liu S P, Chen D Q, Duan X B, et al. Monitoring of the four famous Chinese Carps resources in the middle and upper reaches of the Yangtze River [J]. Resources and Environment in the Yangtze Basin, 2004, 13(2): 183—186 [劉紹平,陳大慶, 段辛斌, 等. 長江中上游四大家魚資源監(jiān)測與漁業(yè)管理. 長江流域資源與環(huán)境, 2004, 13(2): 183—186]
[4] Zhang S M, Deng H, Wang D Q, et al. Population structure and genetic diversity of Silver carp and Grass carp from populations of Yangtze River system revealed by RAPD [J]. Acta Hydrobiologica Sinica, 2001, 25(4): 324—330 [張四明,鄧懷, 汪登強, 等. 長江水系鰱和草魚遺傳結(jié)構及變異性的 RAPD 研究. 水生生物學報, 2001, 25(4): 324—330]
[5] Zhu X D, Geng B, Li J, et al. Analysis of genetic diversity among silver carp populations in the middle and lower Yangtze River using thirty microsa-tellite markers [J]. Hereditas (Beijing), 2007, 29(6): 705—713 [朱曉東, 耿波,李嬌, 等. 利用 30 個微衛(wèi)星標記分析長江中下游鰱群體的遺傳多樣性. 遺傳, 2007, 29(6): 705—713]
[6] Wang C Z, Liang H W, Zou G W, et al. Genetic variation analysis of two silver carp populations in the middle and upper Yangtze River by microsatellite [J]. Hereditas (Beijing), 2008, 30(10): 1341—1348 [王長忠, 梁宏偉, 鄒桂偉, 等. 長江中上游兩個鰱群體遺傳變異的微衛(wèi)星分析.遺傳, 2008, 30(10): 1341—1348]
[7] Ji C H, Gu J J, Mao R X, et al. Analysis of genetic diversity among wild silver carp (Hypophthamlichthys molitrix) populations in the Yangtze, Heilongjiang and Pearl Rivers using microsatellite markers [J]. Journal of fisheries of China, 2009, 33(3): 354—371 [姬長虹, 谷晶晶, 毛瑞鑫,等. 長江、珠江、黑龍江水系野生鰱遺傳多樣性的微衛(wèi)星分析. 水產(chǎn)學報, 2009, 33(3): 354—371]
[8] Wang S, Cao X X, Tanniguchi N. PCR-RFLP analysis on mtDNA D-loop region of four populations of silver carp (Hypophthalmichthys molitrix) [J]. Freshwater Fisheries, 2010, 40(4): 3—15 [王淞, 曹曉霞, 谷口順彥, 等. 4 個群體鰱 mtDNA D-loop的 PCR-RFLP 分析.淡水漁業(yè), 2010, 40(4): 3—15]
[9] Tautz D, Renz M. Simple sequences are ubiquitous repetitive components of eukaryotic genomes [J]. Nucleic Acids Research, 1984, 12(10): 4127—4138
[10] Liu L, Liu H Q, Guo Y S, et al. Isolation of microsatellite markers in the common snapping turtle (Chelydra serpentina L.) and its application in genetic diversity analysis in a cultured population [J]. Acta Hydrobiologica Sinica, 2013, 37(4): 669—677 [劉麗, 劉海情, 郭昱嵩, 等. 蛇鱷龜微衛(wèi)星標記的開發(fā)及一個養(yǎng)殖群體的遺傳多樣性分析. 水生生物學報, 2013, 37(4): 669—677]
[11] Wang J J, Tong J G, Zhang Y G, et al. Study on the genetic diversity of two wild populations of Megalobrama pellegrini (teleostei, cyprinidae) [J]. Acta Hydrobiologica Sinica, 2014, 38(5): 975—979[王瑾瑾, 童金茍, 張耀光, 等. 厚頜魴兩個野生群體遺傳多樣性分析. 水生生物學報, 2014, 38(5): 975—979]
[12] Sambrook J, Russell D W. Molecular Cloning: A Laboratory Manual [M]. NewYork: Cold Spring Harbor Laboratory Press, 2001
[13] Guo W J. Studies on population genetic structure and comparative genome mapping in silver carp [D]. Thesisi for Doctor of Science. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, 2013 [郭穩(wěn)杰. 鰱群體遺傳結(jié)構及比較基因組作圖研究. 博士學位論文. 中國科學院水生生物研究所, 武漢, 2013]
[14] Wang Y W, Samuels T D, Wu Y Q. Development of 1030 genomic SSR markers in switchgrass [J]. Theoretical and Applied Genetics, 2011, 122(4): 677—686
[15] Zhang J R, Wei C L. Establishment a SSR-PCR System of Tea Plant Based on the 4300 DNA Analysis System [J]. Journal of Tea Science, 2014, 34(5): 481—488 [張潔茹, 韋朝領. 基于 4300 DNA 分析系統(tǒng)的茶樹 SSR 發(fā)掘方法優(yōu)化與建立. 茶葉科學, 2014, 34(5): 481—488]
[16] Nei M, Li W H. Mathematical model for studying genetic variation in terms of restriction endonucleases [J]. Proceedings of the National Academy of Sciences, 1979, 76(10): 5269—5273
[17] Tamura k, Dudley J, Nei M, et al. MEGA4: Molecular evolutionary genetics aualysisi (MEGA) sofeware version 4.0 [J]. Molecular Biology & Evolution, 2007, 24(8): 1596—1599
[18] Excoffier L, Laval G, Schneider S. Arlequin(version 3.1): An integrated software package for population genetics dada analysis [J]. Evolutionary Bioinformatics Onlion, 2006, 1(1): 47—50
[19] Rice W. Analyzing tables of statistical tests [J]. Evolution, 1989, 43(1): 223—225
[20] Excoffier L, Smouse P E, Quattro J M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to Human mitochondrial DNA restriction data [J]. Genetics, 1992, 131(2): 479—491
[21] Maruyama T, Fuerst P A. Population bottlenecks and nonequilibrium models in population genetics. II. Number of alleles in a small population that was formed by a recent bottleneck [J]. Genetics, 1985, 111(3): 675—689
[22] Pritchard J K, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data [J]. Genetics, 2000, 155(2): 945—959
[23] Liu M, Yu D H, Huang G J. Isolation and screening of microsatellite DNA markers from silver-lip pearl oyster Pinctada maxima [J]. Marine Science, 2010, 34(8): 1—5 [柳明, 喻達輝, 黃桂菊. 大珠母貝微衛(wèi)星 DNA標記的分離與篩選. 海洋科學, 2010, 34(8): 1—5]
[24] Kijas J M H, Thomas M R, Fowler J C S, et al. Integration of trinucleotide microsatellites into a linkage map of Citrus [J]. Theoretical and Applied Genetics, 1997, 94(5): 701—706
[25] Gastier J M, Pulido J G, Sunden S, et al. Survey of trinucleotide repeats in the human genome: assessment of their utility as genetic markers [J]. Human Molecular Genetics, 1995, 4(10): 1829—1836
[26] Nei M. Genetic distance between populations [J]. American Naturalist, 1972, 106(949): 283—292
[27] Cornuet J M, Luikart G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data [J]. Genetics, 1996, 144(4): 2001—2014
[28] Nei M, Maruyama T, Chakraborty R. The bottleneck effect and genetic variability in populations [J]. Evolution, 1975, 29(1): 1—10
[29] Wright S. Evolution and the Genetics of Population Variability Within and Among Natural Population [M]. Chicago: University of Chicago Press, 1978, 4
[30] Chen J M. Studies on Conservation Genetics of the Rare and Endangered Aquatic plant Caldesia grandis (Alismataceae) [D]. Thesis for Doctor of Science. Wuhan University, Wuhan. 2005 [陳進明. 珍稀瀕危水生植物寬葉澤苔草(澤瀉科)的保護遺傳學研究. 博士學位論文,武漢大學, 武漢. 2005]
THE MICROSATELLITE ANALYSIS OF GENETIC DIVERSITY OF FIVE SILVER CARP POPULATIONS IN THE THREE GORGES RESERVOIR OF THE YANGTZE RIVER
PANG Mei-Xia1,2, YU Xiao-Mu1and TONG Jin-Gou1
(1. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
In this study we employed ten microsatellite markers to analyze the genetic diversity of five silver carp populations including Zigui, Wushan, Yunyang, Zhongxian and Mudong in the Three Gorges Reservoir of the Yangtze River. A total of 161 different alleles were detected, and 84 of them were common in all 5 populations. The number of alleles per locus ranged from 7 to 29. In the five populations, the average values of observed and expected heterozygosity ranged from 0.784 to 0.846 and 0.828 to 0.847 respectively, and the mean PIC was 0.797 to 0.817. The Fstvalues were between –0.001 and 0.009, which indicated that the genetic differentiation was not significant. Chi-square test was used to analyze the genotypes based on the Hardy-Weinberg equilibrium, and the P value indicated that some loci in three populations deviated from the HWE. The bottleneck analysis revealed that the silver carp populations from the Three Gorges Reservoir of the Yangtze River might have undergone a recent genetic bottleneck. The genetic similarity coefficient of the five populations was between 0.891 and 0.950, and the genetic distance of the populations was between 0.050 and 0.115. We also constructed the phylogenetic tree of the five silver carp populations based on Nei’s standard genetic distance, and the results showed that genetic distances between the populations were consistent with their geographical distances. The Bayesian analysis also suggested that the silver carp samples from the 5 locations of the Three Gorges Reservoir could be characterized as a single population. Although we did not observe the genetic differentiation, silver carp populations in the Three Gorges Reservoir displayed high genetic diversity. Our study provided valuable information on the genetic resources of silver carps in the Three Gorges Reservoir and in the upstream of the Yangtze River, which should help develop appropriate policies on the conservation and utilization of silver carps.
Silver carp (Hypophthalmichys molitrix); Microsatellite; Wild population; Genetic diversity; The Three Gorges Reservoir
Q346+.5
A
1000-3207(2015)05-0869-08
10.7541/2015.115
2015-02-12;
2015-03-26
中國長江三峽集團公司科研項目(CT12-08-01); 淡水生態(tài)與生物技術國家重點實驗室自主課題(2011FBZ20)資助
龐美霞(1989—), 女, 湖南長沙人; 博士研究生; 主要從事魚類遺傳學研究。E-mail: pang1mei2xia3@163.com
童金茍, E-mail: jgtong@ihb.ac.cn
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
