不同碳源對重組畢赤酵母表達嗜熱β-半乳糖苷酶的影響
2014-12-03 09:17:50李洪波郭祥前張子政
懷化學院學報 2014年11期
李洪波,郭祥前,陳 靜,龍 騰,張子政
(懷化學院1.生命科學系;2.民族藥用植物資源研究與利用湖南重點實驗室,湖南 懷化 418008)
β-半乳糖苷酶(EC3.2.1.23)學名為β-D-半乳糖苷半乳糖水解酶,常用名為乳糖酶,廣泛存在于各種動物、植物、微生物中,可催化乳糖分解成一分子的葡萄糖和一分子的半乳糖[1,2].利用乳糖酶水解乳制品中的乳糖,可將乳糖含量降低70%-80%,解決乳糖不耐癥患者的乳品消費問題[2-4].熱穩定性好的β-半乳糖苷酶具有較高的開發價值,目前多數耐熱β-半乳糖苷酶的最適溫度在60℃以上,高溫下熱穩定性良好,能夠有效降低生產工藝中微生物污染的風險[5].嗜熱β-半乳糖苷酶主要來源于高溫菌,已有許多研究報道表明通過基因重組技術,將嗜熱β-半乳糖苷酶基因導入適合的宿主細胞,可以高效地產生熱穩定性好的重組β-半乳糖苷酶[6-8].碳源是影響重組蛋白在組成型畢赤酵母中分泌表達水平的最主要因素.本研究分別以葡萄糖、果糖、丙三醇、乳糖、蔗糖、麥芽糖六種碳源,分析碳源對組成型重組酵母分泌表達源自嗜熱菌Pyrococcus furiosuβ-半乳糖苷酶的影響,確定組成培養基的最佳碳源.
1 材料與方法
1.1 材料
表達嗜熱菌Pyrococcus furiosuβ-半乳糖苷酶的組成型畢赤酵母轉化子由佛羅里達大學李侍武教授饋贈,胰蛋白胨及酵母提取物購自OXIOD公司,YP 液體培養基包含1%酵母提取物和2%蛋白胨(添加1.8%瓊脂即為YP 固體培養基).鄰-硝基酚-β-D-半乳糖苷 (ONPG)、考馬斯亮藍G-250 及小牛血清(BSA)購自Sigma公司.葡萄糖、丙三醇、果糖、蔗糖、麥芽糖和乳糖為國產分析純.
1.2 方法
1.2.1 碳源對菌體生長速度的影響
將重組酵母轉接至含2%葡萄糖的YP 固體培養基平板活化,28℃培養3 d,挑取重組酵母轉化子單菌落接種至含2%葡萄糖的YP 液體培養基中,28℃、200 rpm 培養36 h.按2%的接種量分別接至含2%以下6種碳源的YP 液體培養基中:葡萄糖、丙三醇、果糖、蔗糖、麥芽糖和乳糖,28℃、200 rpm 搖床培養4 d,每24 h 取樣一次.測定在λ =600 nm 處測定待測液菌體濃度.設畢赤酵母出發菌株在果糖中的培養為對照.
1.2.2 碳源對重組酵母分泌蛋白的影響
分別取在不同碳源條件下的發酵液上清,以牛血清白蛋白為標品,采用Bradford 法測定蛋白濃度,蛋白濃度測定方法參照文獻進行[9].
1.2.3 測定相對酶活
采用鄰硝基苯酚法測定酶活,參照標準進行[9],分別取樣品上清100 μL,快速加入1.4 mL 0.25% ONPG溶液,立即搖勻將樣品置于65℃水浴鍋中水浴3 min.立即加入0.5 mL NaCO3溶液終止反應.空白對照為加入待測液后先加入NaCO3溶液,后加入ONPG 溶液,同樣水浴3 min 后,以空白為對照,在分光光度計λ =420 nm 處測定各樣品的OD值.以酵母出發菌株在2%葡萄糖的YP 液體培養基中培養的菌液上清為對照.
1.2.4 目標蛋白的SDS-PAGE 分析
取不同碳源在不同培養時間下的重組酵母發酵液上清液TCA 沉淀后,100%乙醇洗滌蛋白沉淀3次,含8 M 尿素的PBS 溶解蛋白沉淀,加入SDS 上樣緩沖液,煮沸熱變性5 min,12% SDS-PAGE 電泳分析.
2 結果與分析
2.1 碳源對菌體生長速度的影響
不同碳源下菌體生長曲線如圖1所示,丙三醇中的酵母菌體濃度最高,麥芽糖最低.丙三醇中的酵母第1 d 即進入對數期,第2-4 d 維持在穩定期,菌體濃度在培養4 d 時為各碳源最高;果糖、葡萄糖及丙三醇的菌體生長曲線趨同.以蔗糖、乳糖及麥芽糖為碳源,培養2 d 內處于停滯期,之后進入對數期;以蔗糖為碳源于第3 d 進入穩定期,以麥芽糖和乳糖為碳源,培養4 d 后仍處于菌體生長的對數期.菌體生長曲線說明,單糖和丙三醇為碳源,菌體的生長明顯快于雙糖,當以乳糖和麥芽糖為碳源時菌體生長的停滯期和以數生長期最長.出發菌株在果糖中的生長曲線和菌體濃度與重組酵母在果糖中的情況基本一致.在不同碳源中,重組酵母菌的菌體生長情況是:丙三醇>果糖>葡萄糖>蔗糖>麥芽糖>乳糖.
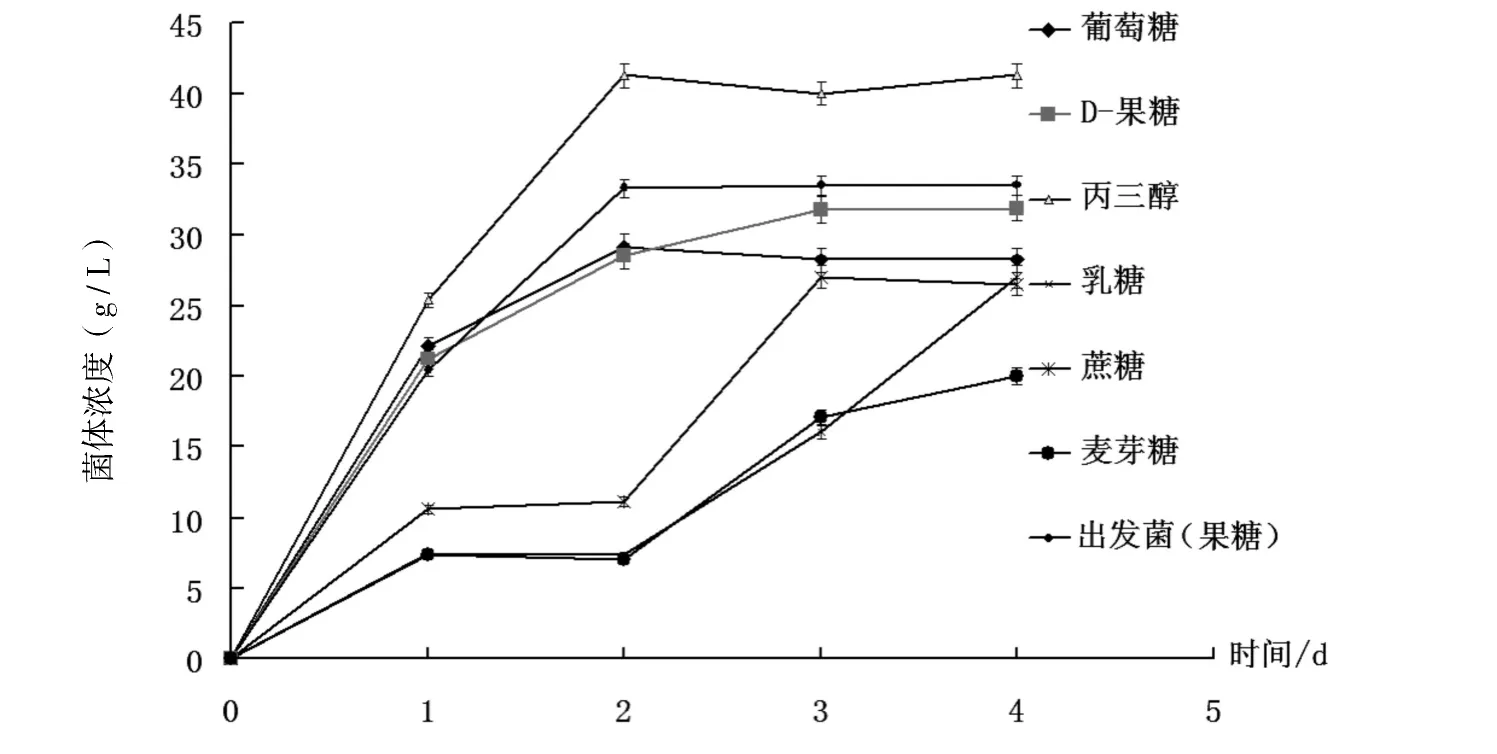
圖1 不同碳源下酵母的菌體生長曲線
2.2 碳源對重組酵母分泌目標蛋白的影響
不同碳源培養的培養基上清中的總蛋白濃度如圖2所示,以葡萄糖、果糖和丙三醇為碳源,酵母分泌的總蛋白在第2 d 迅速上升,之后維持在一個相對穩定的水平;乳糖在1-2 d 蛋白濃度較低,而3-4 d 總蛋白濃度迅速上升,并與上述3種碳源達相近水平.以麥芽糖和蔗糖為碳源時,發酵液中的總蛋白濃度在4 d 內均處于較低水平.菌體分泌蛋白的曲線說明,單糖和丙三醇為碳源菌體,發酵液中的總蛋白濃度較高,重組酵母菌分泌蛋白的能力在3種碳源中相近,當以乳糖為碳源,在培養的第4 d 達到與以上3種碳源接近的水平,當以麥芽糖和蔗糖為碳源時在培養時間范圍內,重組酵母轉化子僅能分泌少量蛋白.
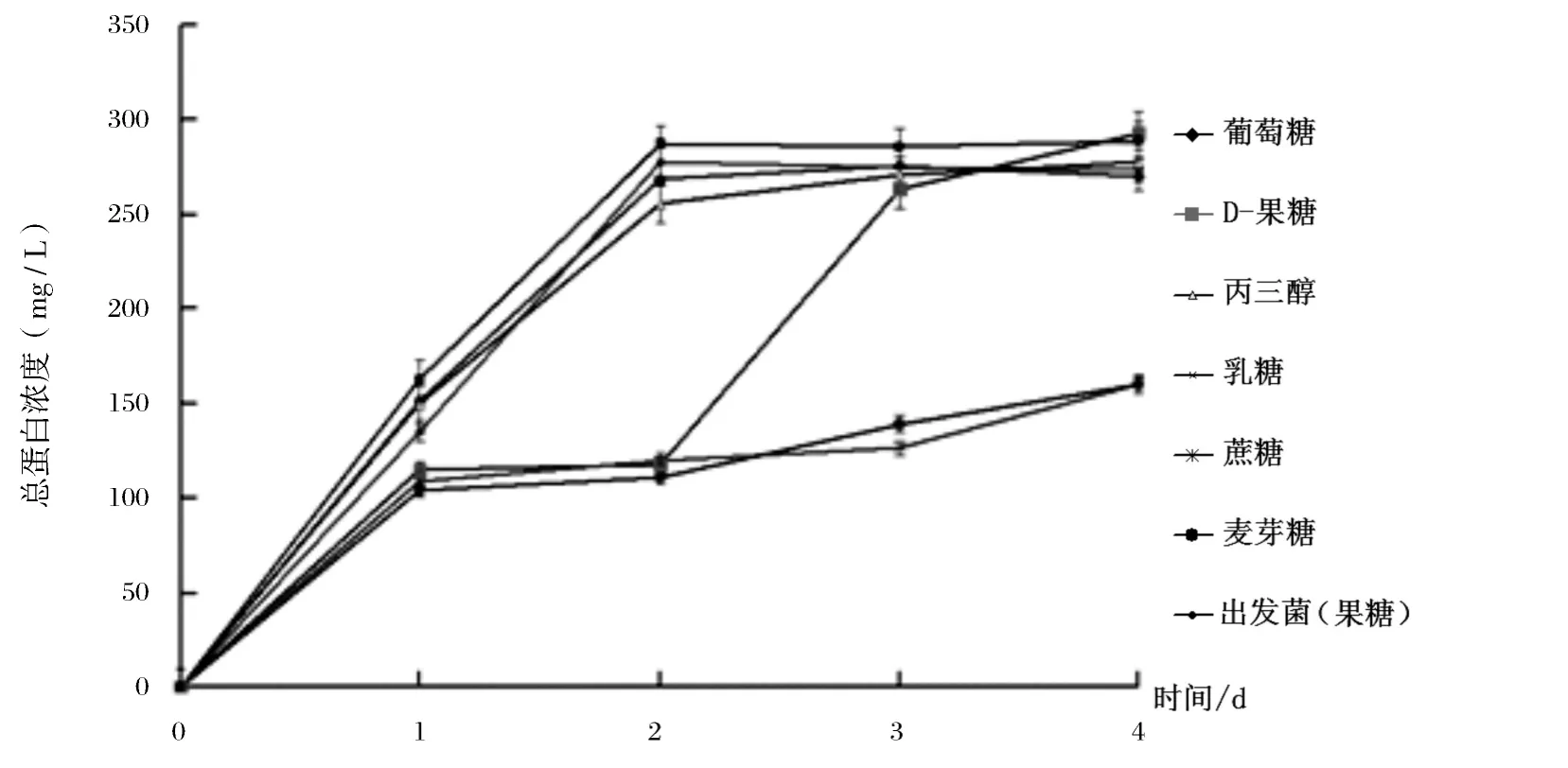
圖2 不同碳源下樣品的總蛋白濃度
2.3 碳源對重組酵母分泌β-半乳糖苷酶酶活的影響
由圖3可知,除畢赤酵母出發菌株的酶活始終很低外,6種碳源都能使重組酵母分泌β-半乳糖苷酶.在培養的第1 d,葡萄糖為碳源酶活最高,而第2 d 果糖為最高;果糖的酶活在第4 d 有所下降,葡萄糖從3 d 開始,酶活稍有下降;以丙三醇為碳源,在培養的4 d 內,酶活一直在增加,但其酶活要低于果糖和葡萄糖.當以蔗糖、麥芽糖和乳糖雙糖為碳源時,酶活在培養時間內始終處于較低水平,且總體穩定,沒有明顯增加.酶活測定結果表明,果糖是6種碳源中最好的碳源,其次是葡萄糖和丙三醇;蔗糖、麥芽糖和乳糖雙糖在培養重組酵母時,其分泌的β-半乳糖苷酶酶活較低.
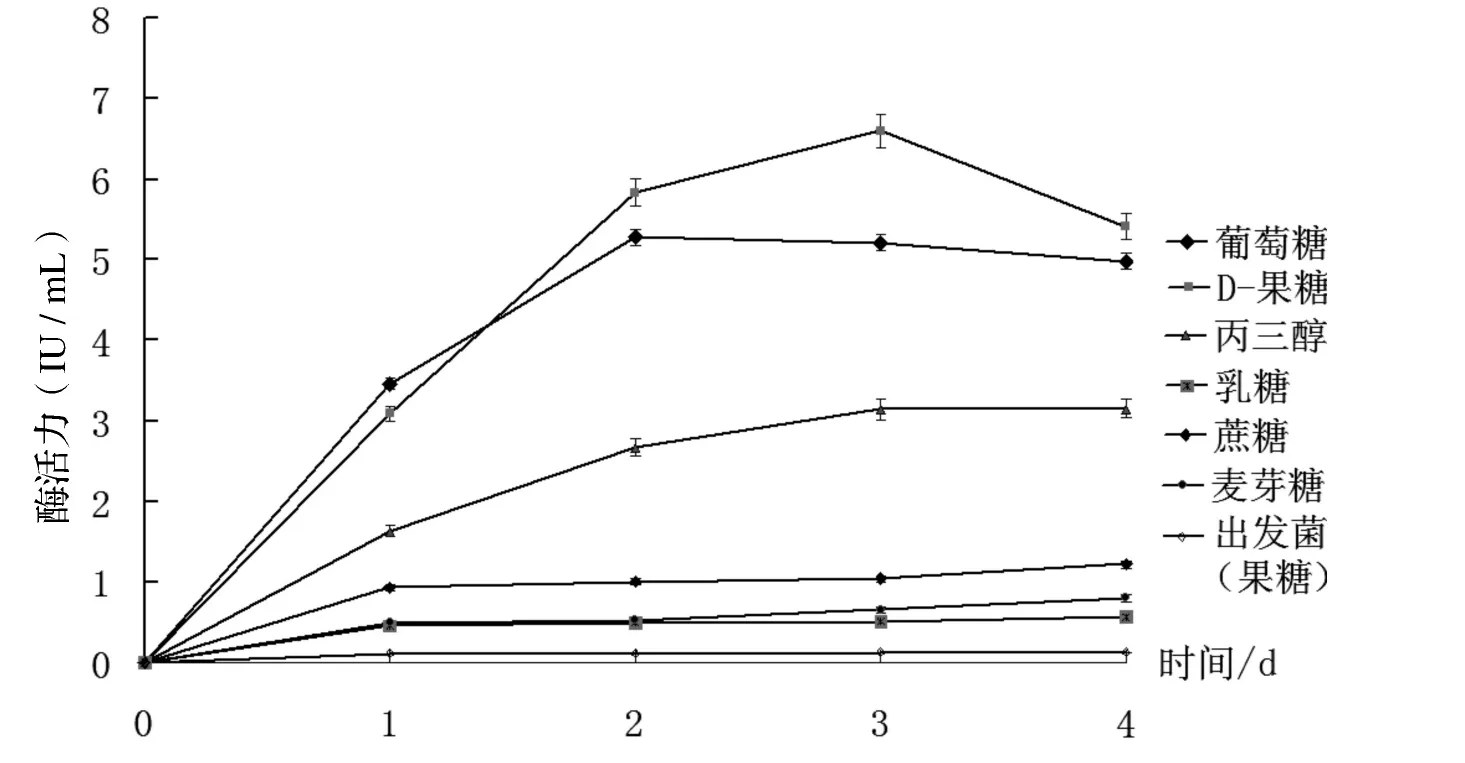
圖3 不同碳源下β-半乳糖苷酶的相對酶活力
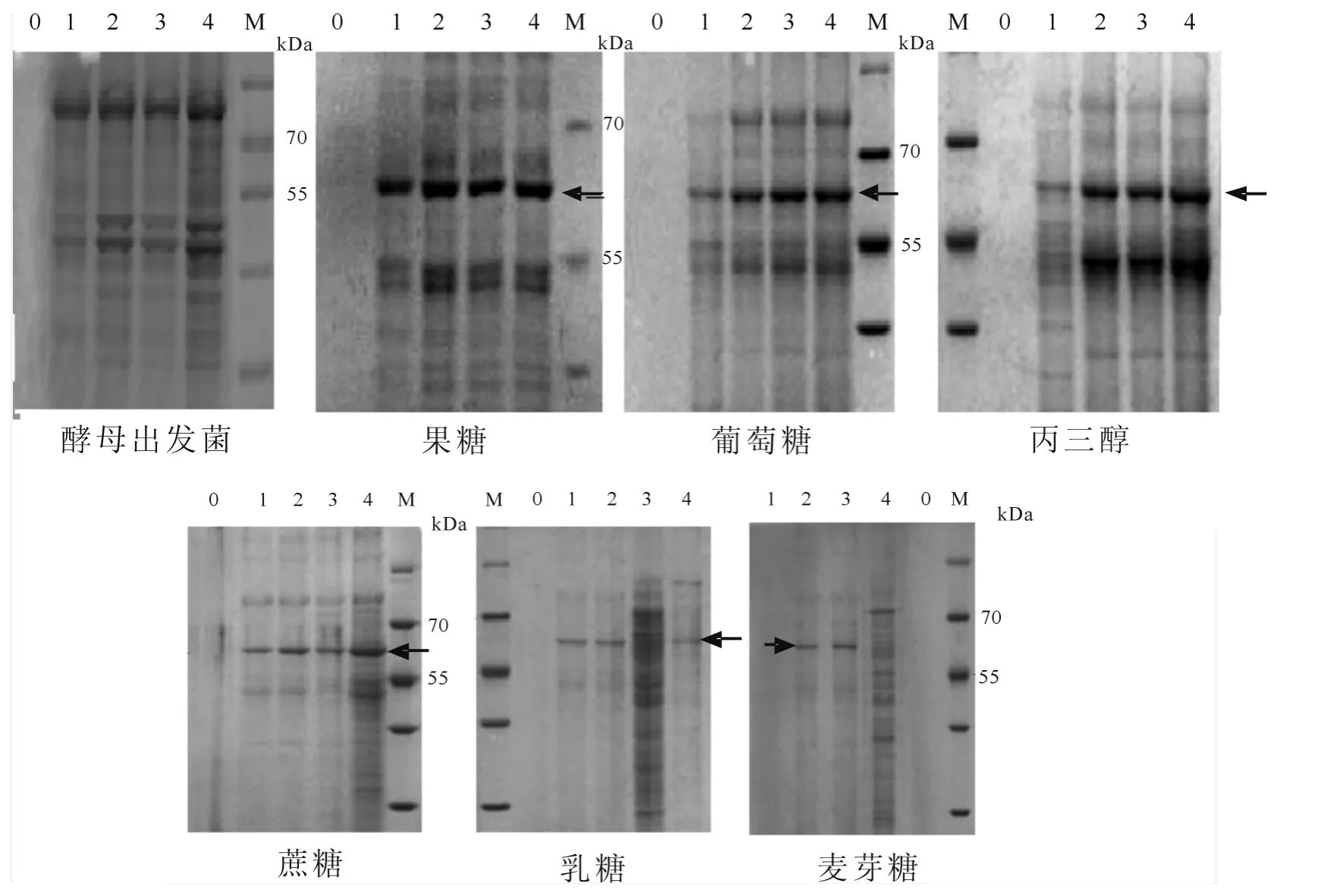
圖4 不同碳源下目標蛋白的SDS-PAGE 圖譜
2.4 不同碳源對重組酵母分泌目標蛋白的SDS-PAGE 分析
重組嗜熱菌Pyrococcus furiousβ-半乳糖苷酶的分子量大約為60 kDa,出發菌株以果糖為碳源的蛋白分泌和重組酵母菌株在6種碳源中的蛋白分泌情況的SDS-PAGE 結果如圖4所示.從SDS-PAGE 結果可以看出,出發菌株以果糖為碳源其培養基上清0、1、2、3、4 d 樣品在目標位置未出現明顯的特異蛋白條帶;當以果糖為碳源時,其培養基上清在培養前沒有特異條帶,在培養的1、2、3、4 d 中,60 kDa 處出現特異蛋白條帶,該條帶在培養的第1 d 就已經非常明顯;當以葡萄糖為碳源時,其培養基上清在培養前沒有特異條帶,在培養的1、2、3、4 d 中,60 kDa 處出現特異蛋白條帶,該條帶在培養的第1 d 就已經較明顯并持續增加到第4 d;當以丙三醇為碳源時,其培養基上清在培養前沒有特異條帶,在培養后,60 kDa 處出現特異蛋白條帶,該條帶在培養的第1 d 就已經較明顯并持續加到第4 d;當以蔗糖、麥芽糖和乳糖雙糖為碳源時,60 kDa 處出現特異蛋白條帶,但始終較弱.SDS-PAGE的檢測結果與酶活測定結果基本吻合.SDS-PAGE 結果同樣表明,以單糖和丙三醇為碳源時,β-半乳糖苷酶的分泌不平要明顯高于雙糖.
3 討論
目前,商用乳糖酶以耐熱性較差的胞內酶為主,這些胞內酶的產率低并且分離純化過程比較復雜,使得商業用乳糖酶的市場價格較昂貴.從食品安全性考慮,在乳業生產上加入低溫酶容易引起污染,存在食品質量安全隱患.耐高溫乳糖酶在100℃經幾十分鐘甚至數小時熱處理仍保持很高的活性,并且在高溫條件下,乳糖更易水解.牛乳消毒主要采用巴氏消毒法或高溫瞬時消毒法,耐高溫酶在上述消毒甚至滅菌條件下仍能滿足去除乳糖的要求,耐高溫乳糖酶的產業化前景廣闊[10-13].本研究主要分析碳源對組成型重組酵母分泌源自嗜熱菌Pyrococcus furiosuβ-半乳糖苷酶的影響,結果發現重組畢赤酵母在不同碳源下分泌表達重組β-半乳糖苷酶的能力存在明顯差異,并得出6種碳源中果糖為酵母產β-半乳糖苷酶的最佳碳源,葡萄糖、丙三醇次之,且單糖碳源要優于雙糖,畢赤酵母生長與β-半乳糖苷酶合成同步,在生長的對數期酶活力最高.
[1]高秀榮,馬力,葉華.β-半乳糖苷酶的研究進展[J].生物技術通報,2005,30 (3):18-20.
[2]張莉,李慶章,田雷.β-半乳糖苷酶研究進展[J].東北農業大學學報,2009,40 (7):128-131.
[3]卜建斌,云戰友,刑黎明.β-半乳糖苷酶的應用及固定化[J].農產品加工學刊,2006,10 (2):4-6.
[4]Sieber R,Stransky M,de Vrese M.Lactose intolerance and consumption of milk and milk products[J].Ernahrungswiss,1997,36 (4):375-393.
[5]董藝凝,陳海琴,劉小鳴,等.耐熱β-半乳糖苷酶的研究進展[J].食品工業科技,2012,33 (1):384-387.
[6]Panesar PS,Kumari S,Panesar R.Potential Applications of Immobilized β-Galactosidase in Food Processing Industries[J].Enzyme Research.2010,27 (1):1-16.
[7]田小梅,任建洪,房聰.不同碳源下畢赤酵母GS115 蛋白組學分析[J].中國生物工程雜志,2012,32 (1):21-29.
[8]劉建福,王璋.許時嬰.碳源對K.fragilis LFS-8611 β-半乳糖苷酶合成的影響[J].工業微生物,2004,34 (3):24-26.
[9]Frederick FM,Brent R,Kingston RE,et al.精編分子生物學實驗指南(第五版)[M].金由辛,包慧中,趙麗云,等譯.北京:科學出版社,2008.
[10]高兆建,侯進慧,孫會剛,等.耐高溫β-半乳糖苷酶的分離純化與酶學性質分析[J].食品科學,2003,31 (23):151-156.
[11]蔣燕靈,朱儉,俞一萍.耐壬乳糖酶的純化及理化性質研究[J].中國食品學報,2005,5 (1):43-46.
[12]Koning S,Elferink M,Konings W,et al.Cellobiose Uptake in the Hyperthermophilic Archaeon Pyrococcus furiosus Is Mediated by an Inducible,High-Affinity ABC Transporter[J].Journal of bacteriology.2001,183(17):4979-4984.
[13]Flores MV,Ertola RJ,Voget CE.Effect of monovalentcations on the stability and activity of kluyveromyces lactis β-galactosidase[J].Lebensm-Wiss u-Technol,1996,29 (5):503-506.
