小鼠骨髓樹突狀細胞體外培養擴增與生物學特性研究
2014-12-02 04:23:52劉曉玲郭紅月宋振川
腫瘤預防與治療 2014年1期
關鍵詞:小鼠
劉曉玲,郭紅月,董 猛,宋振川
(河北省滄州中西醫結合醫院,河北滄州061001)
樹突狀細胞(dendritic cell,DC)是抗原遞呈細胞(antigen-presenting cell,APC),也是唯一能將抗原遞呈給初始T細胞激發初次免疫應答的抗原呈提細胞,被認為是機體免疫的始動者,在抗腫瘤免疫方面發揮著重要作用,基于DC的免疫治療被認為是最具前景的抗腫瘤治療[1],本研究采用簡便方法體外分離培養擴增小鼠骨髓DCs,為抗腫瘤疫苗的實驗研究奠定基礎。
1 材料與方法
1.1 材料
SPF級615小鼠,雌雄各半,鼠齡6~8周齡,體重17~21g,購于天津中國醫學科學院血液研究所實驗動物中心。胎牛血清(杭州四季青生物有限公司)、RPMI1640培養基(美國GIBICO公司)、PBS粉(北京中杉金橋生物技術有限公司)rmIl-4、rmGMCSF、rmTNF-α、FITC 標記抗鼠單克隆抗體、PE 標記抗鼠CD86單克隆抗體均購于美國BioLegend公司。CO2培養箱(美國 SHELDON)、流式細胞儀(美國Beckman Coulter公司)、倒置顯微鏡(日本OLYMPUS)。
1.2 實驗方法
1.2.1 小鼠骨髓單核細胞分離 將615小鼠拉頸處死,浸入75%酒精消毒5min,無菌狀態下取脛骨和股骨,剪掉股骨和脛骨骨兩端,用1ml一次性使用注射器抽取PBS液,刺入骨髓腔反復沖洗,骨髓細胞懸液過400目濾網去除組織碎片,收集細胞懸液離心(1 500r/min,5min),棄上清,以1∶10體積比加入37℃預溫的紅細胞裂解液重懸使紅細胞裂解,同法PBS液離心清洗2次。
1.2.2 骨髓源樹突狀細胞(BMDC)的培養擴增用全培養基懸浮細胞,細胞計數,以RMPI-1640全培養液配成2×106個/ml的細胞懸液,接種于6孔板,每孔中加入完全培養基至4ml,再加入GM-CSF(終質量濃度20ng/ml),置于37℃、5%CO2培養箱中進行培養。第3天輕輕吹打,吸去培養液和懸浮細胞,保留貼壁細胞加入新鮮的等量RMPI-1640全培養液和相同濃度的GM-CSF及終濃度為10ng/ml的IL-4,繼續培養,隔日半量換液,補充RMPI-1640全培養液和GM-CSF及IL-4。繼續培養至第8天,加入TNF-α(10ng/ml),孵育48小時收集懸浮細胞即為成熟的DCs。
1.2.3 細胞生長狀態觀察 每天相差顯微鏡下觀察DC的形態、數量、分布、貼壁情況、細胞集落生長情況并拍照。
1.2.4 DC細胞表型流式細胞術分析 分別收集加入TNF-α前及加入TNF-α 48小時后的DC懸液,離心沉淀(1 000r/min,5min),去上清,加PBS 4ml懸垂,再離心2次,用PBS緩沖液調整細胞濃度至1 ×106個/ml,加入 Eppendorf管,每管 100μl,然后分別加入 FITC anti-mouseCD11c、Peanti-mouseCD86單抗各2μg,4℃避光孵育30min,PBS液洗3次后用,流式細胞儀檢測各組DC表面CD11c、CD86的表達情況,將加入TNF-α前后組作為實驗組,未加入抗體的DC組作為對照組,比較其表面標志物的表達。
1.3 統計學方法
所有數據均采用SPSS13.0統計軟件分析,DC表型表達陽性率數據分析采用χ2檢驗,以均數±標準差(±s)表示,以 P<0.05為差異有統計學意義。
2 結果
2.1 培養細胞的形態學觀察
經分離提純的小鼠骨髓細胞,培養24小時后相差顯微鏡下可見培養板底部有半貼壁細胞,少量細胞集落形成,簇狀生長,細胞體積小,圓形,細胞無明顯突起(圖1a);體外培養3天后可見貼壁細胞逐漸增多,松散粘附于培養板的細胞集落增多,散在少量DC,半貼壁,細胞體積有所增大,細胞核可見,為較小圓或橢圓形,可見貼壁的單核巨噬細胞(圖1b);第5天可見部分細胞脫壁,從細胞集落中釋放出來,呈半貼壁狀態,細胞體積變大,形態不規則,有部分突起,即為未成熟DC(圖1c~圖1e);培養第7天,細胞體積更大,培養液中有更多懸浮細胞,細胞集落散在其中,更加松散,形態趨于特征性星形,有2~5個不等的突起,懸浮細胞為擴增之骨髓樹突狀細胞(圖1f~1g);加入TNF-α 24h后,大量形態典型的DC從集落釋放,細胞生長旺盛,體積增大,有明顯樹枝狀突起,突起粗大且長,胞體飽滿,呈星形、多邊形、或梭行(圖1h~1j)。每只615小鼠平均能分離出3×107個骨髓細胞,經培養可獲得7×106個DC。
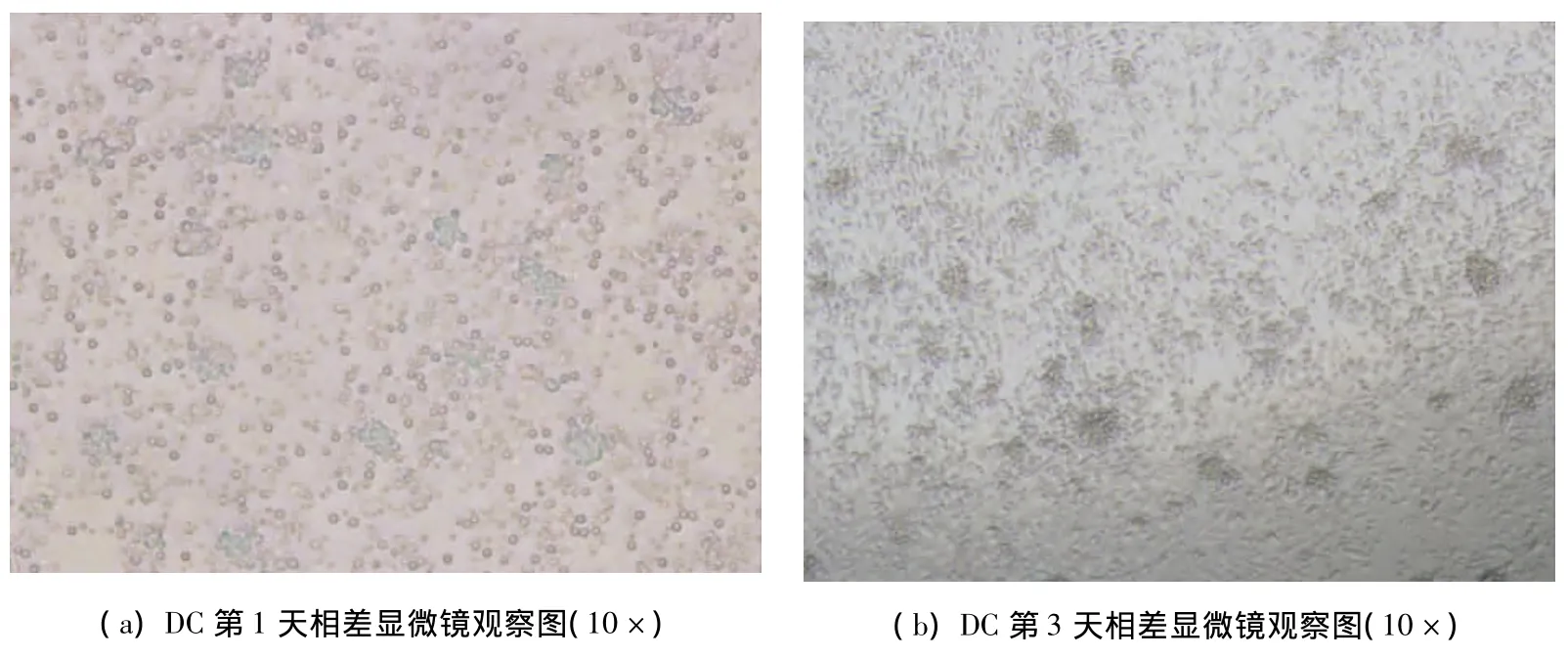
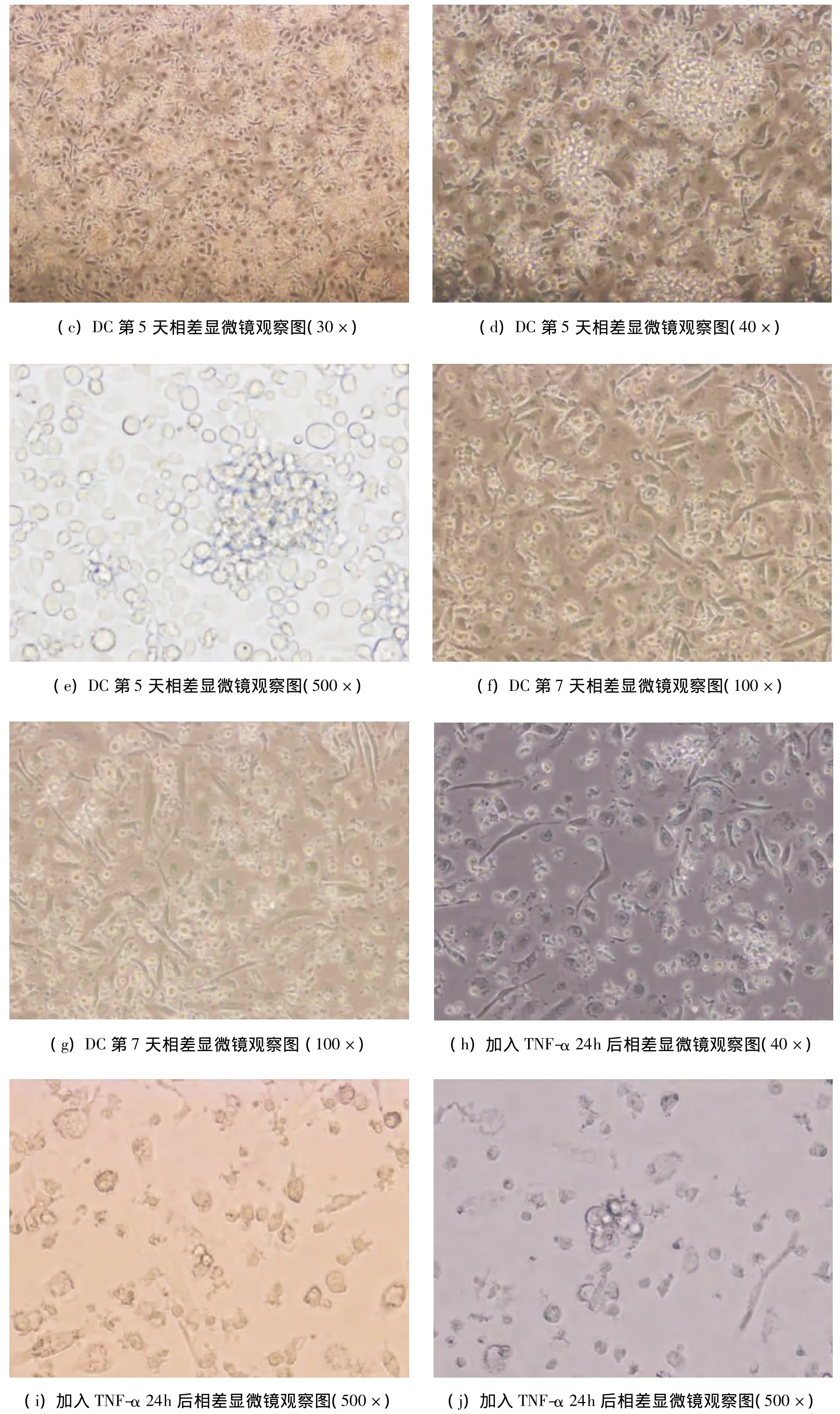
圖1 DC培養觀察圖
2.2 流式細胞儀檢測小鼠骨髓DC加入TNF-α前后CD11c及CD86的陽性表達情況
將加入TNF-α前后組作為實驗組,將未加抗體的DC作為對照組,結果顯示,CD11c陽性表達率加入 TNF-α 前為:(65.09 ±3.04)%(圖 2),加入TNF-α后為:(70.99 ± 1.44)%(圖 3)(χ2=6.625;p=0.097);CD86陽性表達率加入 TNF-α 前為(25.80±0.30)%(圖 2),加入 TNF-α 后為(55.91 ±2.41)%(圖 3)(χ2=6.856;p=0.032)。CD86 加入TNF-α前陽性表達率低于加入TNF-α后,差異有統計學意義,CD11c的陽性表達率加入TNF-α前后差異無統計學意義(p>0.05)。見表1。

圖2 加入TNF-α前CD11c、CD86陽性表達率
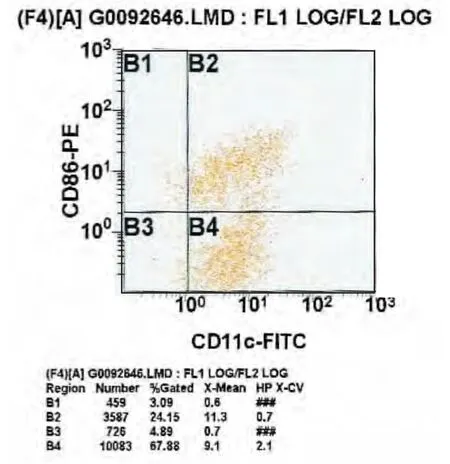
圖3 加入TNF-α后CD11c、CD86陽性表達率
表1 流式細胞儀檢測樹突狀細胞CD11c CD86陽性表達率 [(±s)%]
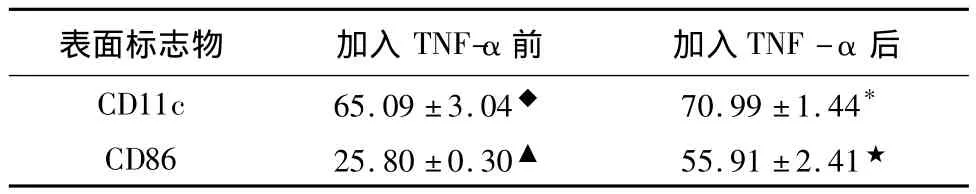
表1 流式細胞儀檢測樹突狀細胞CD11c CD86陽性表達率 [(±s)%]
◆組與* 組比較,P >0.05(χ2=6.625;p=0.097)▲組與★組比較,P <0.05(χ2=6.856;p=0.032)
表面標志物 加入TNF-α前 加入TNF-α后CD11c 65.09 ±3.04◆ 70.99 ±1.44*CD86 25.80 ±0.30▲ 55.91 ±2.41★
3 討論
樹突狀細胞是由Steinman和Cohn首先由小鼠脾臟組織中分離發現的,因其具有典型的樹突狀突起而命名[2]。在接受抗原刺激成熟后,DC可以有效地攝取抗原,并通過抗原肽/MHC分子復合物的形式,將抗原有效提呈給初始T細胞,刺激生成針對該抗原的特異性細胞毒T細胞,從而有效啟動機體免疫反應[3]。目前,已能夠通過體外抗原沖擊、基因轉染、細胞融合等方法制備出相應的DCs腫瘤疫苗,并且在動物實驗中獲得很好的效果[4-6]。
DC主要來源于骨髓的CD34+造血干細胞,故可以從骨髓細胞中分離誘導DC,獲得一定量有功能的DC是深入研究其生物學功能的關鍵,然而DC在體內分布分散,含量極低,在動物和人外周血中尚不足白細胞總量的1%,在實質器官中低于局部細胞1%,在血液中低于有核細胞的0.1%,故如何體外大量擴增DCs成為研究者目前急需解決的問題[7]。
在小鼠DC的體外分離培養擴增中添加細胞因子是最重要的環節,GM-CSF是維持DC發育及分化最根本的細胞因子,但是單一使用GM-CSF只能產生少量的DC集落,并且還會刺激骨髓細胞中的巨噬細胞、中性粒細胞大量增殖,影響DC純度,而IL-4能抑制培養物中巨噬細胞和中性粒細胞的產生,并能使DC具有典型形態和很強的處理外源性抗原能力[8]。通過聯合應用GM-CSF和IL-4進行體外誘導培養至第7天時,加入脂多糖(LPS)繼續培養24h,能獲得大量成熟 DCs[9]。
在本研究開始,GM-CSF及IL-4濃度配比較低,骨髓細胞生長緩慢,調整GM-CSF濃度至20ng/ml,IL-4濃度至10ng/ml,細胞數量增多,體積增大。本實驗在第三天開始加入IL-4,而不是在本實驗開始前加入,是考慮IL-4與GM-CSF作用相抵觸之處。TNF-α能抑制粒細胞的產生,同時誘導 DC的成熟[10],可以將成熟的DC數量提高10%~15%。
由于DC來源復雜,到目前為止,還沒有發現一種表面分子能夠代表不同組織及不同分化時期的所有DC的特異性標記,目前通常采用細胞形態、表型及異基因淋巴細胞刺激能力三者相結合的方法。
據報道33D1、凝集素類似受體 NLDC-k145、DEC-205及整合素CD11c[11]被認為是小鼠DC較特異性的表面標記,在這3種表面分子中,CD11c是相對分子質量為150 000的β2整合素家族成員,髓樣樹突狀細胞表達髓源性抗原CD11c,CD11c是確定DCs特異性表面標志物之一,與CD86相比,包括DC前體、未成熟及活化成熟的DC均有較高的表達率[12]。CD86是細胞表面黏附因子,也是激活T細胞的第2信號分子,可作為DC的活性指標[13]。
不成熟DC具有很強的吞噬、加工處理抗原的能力,但是對T細胞的刺激功能比較低,其表面低表達甚至不表達MHC-Ⅱ類分子和共刺激分子如CD40、CD80和CD86。未成熟DCs捕獲抗原后,并在一些活化因子(如 LPS、IL-1β、TNF-α 等)作用下可分化為成熟DCs,其表面MHC-Ⅱ類分子及共刺激分子的表達顯著上調,其介導的免疫反應增強,對T細胞的刺激功能增加,但抗原攝取能力下降[14]。
本實驗流式細胞儀檢測表面標志物,經統計學分析,CD11c添加TNF-α前后的陽性表達率無差別,CD86加入TNF-α后陽性表達率較加入 TNF-α前明顯增加。說明未成熟DC和成熟DC,CD11c均有較高表達,成熟DC較未成熟DC,CD11c表達無明顯增高,說明CD11c是DC的特異性表面標志物,CD86在成熟DC表達較未成熟DC高。說明TNF-α能提高CD34+干細胞表面GM-CSF受體數量,促進細胞表達CD86,阻斷粒細胞分化途徑,促進DC分化、成熟[15]。
本實驗經rmGM-CSF和rmIL-4聯合誘導,可以從小鼠骨髓獲得大量具有典型DC形態學特征及生物學特征的樹突狀細胞,這為DC疫苗的研究及臨床應用奠定了基礎。
[1] 徐 榕,江虹虹,宋海峰.樹突狀細胞疫苗研究的新策略[J].中國腫瘤,2011,20(2):92-97.
[2] Steinman RM,Chon ZA.Identification of a novel cell type in peripheral lymphoid organs of mice.Imorphology,quantitation,tissue distribution[J].Exp Med,1973,137(5):1142-1162.
[3] Chauvin C,Josien R.Dendritic cells as killers:mechanistic aspects and potential roles[J].J Immunol,2008,139(5):671-676.
[4] Brok MH,Nierkens S,Figdor CG,et al.Dendritic cells:tools and targets for antitumor vaccination[J].Expert Rev Vaccines,2005,5(4):699-710.
[5] Banchereau J,Palucka AK.Dendritic cells as therapeutic vaccines against cancer[J].Nat Rev Immunol,2005,5(4):296-306.
[6] Gong J,Koido S,Calderwood SK.Cell fusion:from hybridoma to dendritic cell-based vaccine[J].Expert Rev Immunol,2008,7(7):1055-1068.
[7] 易輝君,盧光琇.不同來源血清或血漿培養條件下的樹突狀細胞表型和功能的比較[J].細胞與分子免疫學雜志,2012,28(6):561-563.
[8] 郭慈仁,歐啟水,李良慶.小鼠骨髓來源樹突狀細胞體外擴增和鑒定[J].福建醫科大學學報,2004,38(3):261-263
[9] 劉 錚,代繼宏,符 州,等.小鼠骨髓源樹突狀細胞體外誘導培養及初步鑒定[J]。南京醫科大學學報,2011,28(2):25-31.
[10]孫國龍,王凱忠,周 莉,等.小鼠骨髓及脾臟來源的樹突狀細胞培養及鑒定[J].中國實驗診斷學,2012,16(3):408-411
[11] Makala LH,Nishikawa Y,Mishima M,et al.Phenotype and unction of murine peritoneal cavity macrophage derived dendriticells[J].J Vet Med Sci,2002,64(9):813-820
[12] Reschner A,Hubert P,Delvenne P,et al.Innate lymphocyte and dendritic cell cross-talk:a key factor in the regulation of the immune response[J].Clin Exp Immunol,2008,152:219-226.
[13] Guermonp rez P,Valladeau J,Zitvogel L,et al.Antigen p resentation and T cell stimulation by dendritic cells[J].Immunol,2002,20:621-667.
[14] Liu Q,Shu X,Sun A,et al.Plant-derived small molecule albaconol suppresses LPS triggered proinflammatory cytokine production and antigen presentation of dendritic cells by impairing NF-kB activation [J].Int Immunopharmacol,2008,8:1103-1111.
[15]熊 菲.影響樹突狀細胞分化成熟因素的研究進展[J].國外醫學免疫學分冊,2005,28(2):107-111.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
