日本佳盾蜾蠃的生物學特性觀察
2014-11-25 02:58:38黃敦元余江帆許曉風
環境昆蟲學報 2014年6期
黃敦元 ,何 波,谷 平,余江帆,彭 飛,許曉風
(1.江西環境工程職業學院,江西贛州 341000;2.南京師范大學生命科學學院動物學國家重點學科,江蘇生物多樣性重點實驗室,南京 210046;3.江西省林業科技培訓中心,南昌 330038)
蜾蠃亞科Eumeninae 是胡蜂科中最大的一個類群(Carpenter,1982;Kim et al.,2005),目前研究發現,全世界共有3585 種,我國現有記錄161 種(Yamane 1990;Kim and Yamane,2001;周鑫等,2011,2012,2013;黃敦元等,2013;You et al.,2013)。蜾蠃亞科多為獨棲物種,無固定居住巢穴,一般交配后開始營巢產卵繁育后代,其巢穴有兩種類型:一種是呈中空壺狀,上部有細頸,開口于頸端部的泥質巢穴 (李鐵生,1982);另一種巢是利用開口于竹管、蘆葦管等中空材料,銜泥分隔成多個蟲室,直接將卵產于竹管或者葦管內壁上,產卵后外出捕捉蛾類幼蟲帶回蟲室中以供其卵孵化出的幼蟲食用,最后用泥塊密封蟲室和巢穴口 (Cowan,1991;Fateryga,2013)。
日本佳盾蜾蠃Euodynerus nipanicus (Schulthess)屬膜翅目Hymenoptera 胡蜂科Vespidae 蜾蠃亞科Eumeninae 佳盾蜾蠃屬Euodynerus Dalla Torre 1904(李鐵生,1982),該蜂主要分布在中國的江蘇、浙江、江西和臺灣等地(黃敦元等,2013),以及日本和韓國等地(Kim,2012),是一種重要的獨棲性天敵昆蟲,其捕食兇猛,飛翔迅速,對林區鱗翅目蛾類害蟲的控制作用明顯 (李鐵生,1985)。本研究利用人工巢管收集并室內觀察該蜂,旨在進一步明確其生物學習性,以期為日本佳盾蜾蠃的保護與利用提供生物學依據。
1 材料與方法
1.1 研究地區概況
調查的時間從2011年2月到2013年11月。調查地點為江西省贛州市贛縣沙地鎮 (N:26°06';E:114°46')、江西省贛州市南康市譚東鎮(N:25°76';E:114°87')和江西省贛州市湖邊鎮 (N:25° 53';E:114°54')共3個典型樣地。
江西省贛州市氣候特點:氣候溫暖、光照充足、雨量充沛、四季分明,春季天氣多變且雨量充沛,回暖較早;夏季高溫;秋季涼爽干燥;冬季陰雨天氣較多,年平均氣溫在19.1℃至20.4℃,年平均無霜期287 天,年平均降雨量1605.6 mm,4-6月為雨季,占全年雨量的47%,年平均日照時數為1774.4 小時。
1.2 研究方法
1.2.1 幼蟲的收集與觀察
利用人工巢穴(trap-nesting) (圖版Ⅰ-1)野外收集日本佳盾蜾蠃,實驗室解剖觀察并記錄各蟲態的發育歷期;記錄日本佳盾蜾蠃的生活史,同時對其寄生性天敵的寄生行為進行觀察。用照相機(Canon Power Shot S3IS)拍攝各個蟲態,自蛹期起每12 h 拍照一次,直至羽化,并用游標卡尺測量各蟲態的長度大小。
1.2.2 羽化及性比的觀察
野外每15 天采集一次筑巢巢管,解剖后統計羽化期每個巢穴中羽化出的雌雄個體數量,據此計算不同世代的性比。
1.2.3 成蟲交配行為的觀察
制作規格為3 m×3 m×3 m的網罩安置在蜂巢附近,將羽化出巢的雌雄個體人為放置在網罩內觀察其交配行為。
1.2.4 成蟲活動習性與壽命觀察
用白色涂改液標記剛筑巢的巢管,對筑巢雌蜂的活動規律進行跟蹤觀察,同時通過野外觀察和室內解剖雌蜂觀察卵巢有無產卵斑相結合的方法(賀春玲等,2011)來確定成蟲的壽命。
1.2.5 雌蜂訪花行為觀察
在日本佳盾蜾蠃成蟲活躍期(2013年4、7、10月),每月隨機選擇4個晴好天氣,巢區周邊800 m 范圍內觀察該蜂到訪的植物種類,輔以拍照和攝像的方法記錄該蜂的訪花行為。
1.3 數據統計與分析
實驗數據存儲和圖形制作采用SPSS 13.0 和Excel 2003 軟件進行處理。
2 結果與分析
2.1 形態特征(成蟲)
日本佳盾蜾蠃雌性個體體長在9.68-12.35 mm 之間,平均11.17 ±0.25 mm (n=15)(圖版Ⅰ-8)。頭部寬略窄于胸部。額部除額溝處有1 黃斑外,全呈黑色,密布粗糙刻點,覆以淺色短毛。褐色單眼呈倒三角形排列于兩復眼頂部之間,觸角支角突黑色,柄節前緣黃色,余黑色,梗節及鞭節均黑色。顱頂部及頰部均黑色,僅復眼后緣上部有1 黃色斑,呈點狀,刻點較淺,覆短毛。唇基半部黃色,端半部黑色,密布刻點及短毛,上唇棕色,上顎楔狀,基部黑色,向端部漸呈暗棕色,近基部邊緣有1 黃斑,齒尖銳。前胸背板前緣截狀,肩角明顯,前緣兩側各有1 黃色橫帶狀斑,余均黑色,密布粗糙刻點,覆淺色短毛。并胸腹節全呈黑色,中部凹陷,有橫皺褶,中央有縱溝,兩側刻點粗糙,覆短毛。中胸側板全星黑色,中部有粗糙刻點,后胸側板全呈黑色,布有橫皺褶,均覆短毛。翅基片黃色,中央有1棕色斑,光滑無毛。腹部第1-4 節背板黑色,僅沿端部邊緣有1 黃色橫帶,密布刻點及短毛。5-6 節背板及各節腹板全呈黑色,僅第2 節腹板兩側常有1 黃色斑,各節有略淺的刻點及短毛。
雄性個體體長在7.98-10.74 mm 之間,平均9.13 ±0.33 mm (n=15) (圖版Ⅰ-8)。體色近似于雌蜂,唇基全呈黃色,中、后足基節前緣黃色,觸角末端節呈鉤狀,腹部共7 節。
2.2 生物學特性
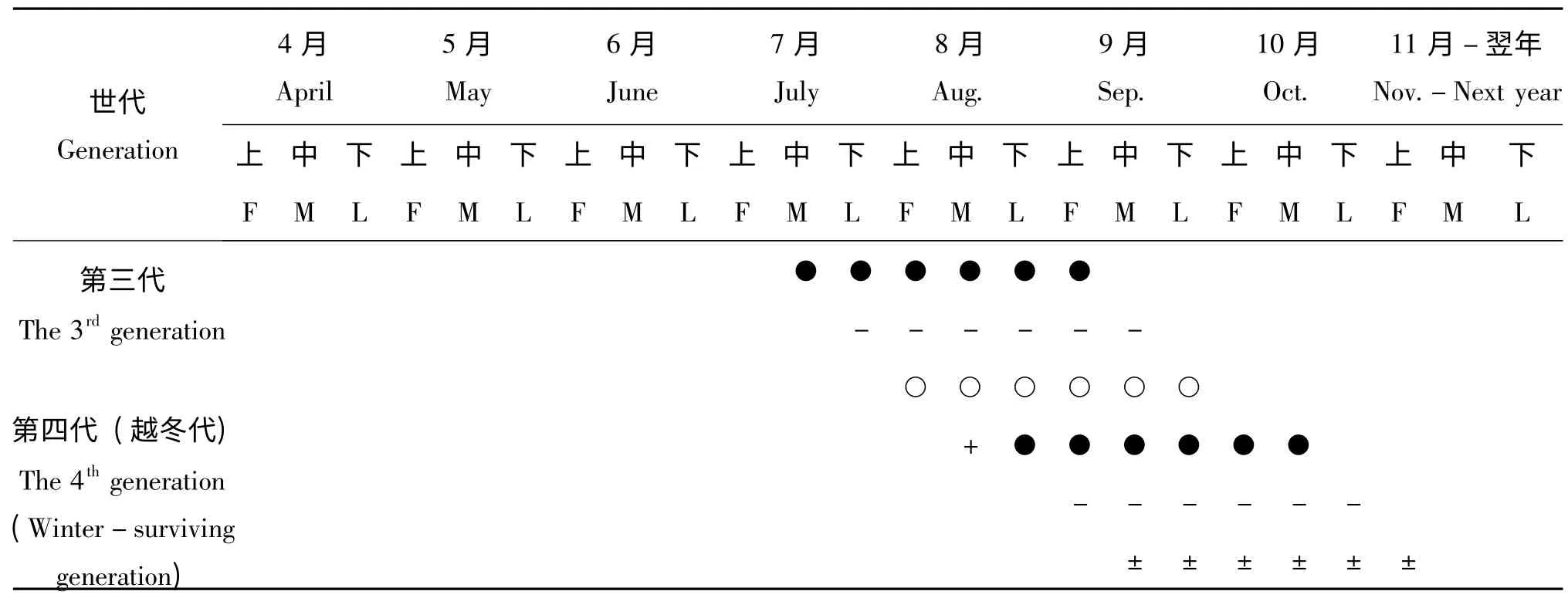
早在2009年8月,我們在江西宜春、贛州等地野外采集過程中發現有日本佳盾蜾蠃的分布,營巢地點多為枯萎的樹枝和竹竿。為了便于研究該蜂的生物學及營巢生物學規律,我們嘗試使用人工巢管(蘆葦管)來收集該蜂,在2010年野外放置的巢管中發現該蜂有大量筑巢現象,2011年開始系統研究該蜂的生物學和營巢規律。日本佳盾蜾蠃的各蟲態形態圖版Ⅰ
2.2.1 生活史
日本佳盾蜾蠃在江西贛州地區1年4 代,并有世代重疊現象,最后一代成蟲10月下旬數量開始減少,11月上旬基本不見其活動,以滯育狀態下的老熟幼蟲在蟲室內越冬,越冬成蟲4月底開始羽化并陸續出巢活動,最后一代的成蟲10月下旬數量開始減少,11月上旬基本不見其活動。自然條件下,越冬幼蟲翌年4月中旬至5月初開始陸續化蛹,蛹期6-9 d。單個巢管中所有成蟲的出巢歷期約3-5 d,一般雄蜂先于雌蜂1-2 d 出巢,雌蜂羽化出巢后即可交配,交配后雌蜂多數在羽化地點附近選址筑巢,雌蜂壽命約25 d。雄蜂壽命約15 d,1 頭雄蜂可與多頭雌蜂交尾。該蜂卵期2-4 d,幼蟲期6-10 d,幼蟲取食完所在蟲室中的食物后開始吐絲做繭,進入老熟幼蟲期,約4-5 d,越冬代滯育期約190 d。日本佳盾蜾蠃在江西贛州地區的年生活史見表1。

表1 日本佳盾蜾蠃的生活史(江西,贛州)Table1 The life cycle of Euodynerus nipanicus (Schulthess)(Ganzhou,Jiangxi)
(續上表)
2.2.1.1 卵
卵近似微彎的長圓柱形(臘腸形),長度一般在2.74-3.06 mm 之間,平均2.91 ±0.13 mm (n=10),直徑在0.78-0.92 mm 之間,平均0.84±0.04 mm (n=10),卵期2-4 d,平均2.5 ±0.99 d (n=10)。剛產的卵無色透明略帶乳白色,表面光滑,24 h 后漸變成淺黃色(圖版Ⅰ-2),以一根1-3 mm 長的白色絲狀物懸掛在蟲室最內端的上壁。大約1-2 d 后,卵的中部會有明顯的膨大,2 d 后卵殼內幼蟲的形態發育完成。體視鏡(40×)下可觀察到蟲體在緩慢地蠕動,然后卵殼從背中線逐漸開裂,以頭部破卵而出,掉落于食物上,幼蟲不取食卵殼。
2.2.1.2 幼蟲
初孵幼蟲呈淺乳白色,體長一般在2.08-3.51 mm 之間,平均2.75 ±0.15 mm (n=10),無足,光滑無毛,體軀柔軟彎曲呈“C”型,頭部小,呈乳白色(圖版Ⅰ-3)。幼蟲吸附在蟲室中的鱗翅目蛾類幼蟲上,先用大顎咬開鱗翅目幼蟲的蟲體,通過身體的蠕動來吸食幼蟲的體液至只剩軀殼。取食過程中能明顯的觀察到低齡幼蟲體液的流動情況,食物在中腸后端開始積累,消化道的顏色也因此逐漸由后向前加深,逐漸能透過身體看到一條淺黃色的消化道。低齡幼蟲(1-3 齡)只取食不排便,幼蟲的體色與被取食幼蟲的體色相似(圖版Ⅰ-4)。4-5 齡幼蟲節間多皺褶,粗肥,啃食鱗翅目幼蟲的軀體,此時幼蟲體色均轉為淺綠或黃白色(圖版Ⅰ-5),取食量明顯增大。5-7 天幼蟲進入吐絲階段,繭的大小一般與蟲室的大小一致,也可能略小于蟲室。幼蟲做完繭后,進行一次排便,排泄物為棕黑色粘稠。之后體色漸為淺黃色,體形由“C”型轉為扁平且直,進入老熟期(圖版Ⅰ-6),幼蟲期6-10 d,平均8.1±0.99 d (n=10)。
老熟幼蟲體粗胖、兩端略尖呈梭形,體表光滑無毛,體節明顯可見且呈深黃色(圖版Ⅰ-7),體長1.209 ±0.261 mm,寬0.412 ±0.098 mm。頭部寬圓,骨化弱,上顎具簡單齒,下顎須及下唇須明顯可見。日本佳盾蜾蠃老熟幼蟲期一般為4-5 d (越冬代老熟幼蟲期及滯育期較長,約190 d)(圖版Ⅰ-7)。
2.2.1.3 蛹
日本佳盾蜾蠃的蛹型為被蛹(圖版Ⅰ-10)。雌雄個體蛹的大小差異較大,雌蛹體長在10.64-13.92 mm 之間,平均12.31 ±1.08 mm (n=15);雄蛹體長在8.84-10.82 mm 之間,平均10.07 ±0.56 mm (n=15)。體色初期乳白色,逐漸由乳白色-淡黃色-土黃色-褐色-黑色轉變(圖版Ⅰ-10);頭胸部的顏色先于腹部變暗;腹部節間處隨頭胸部同時變黑,其余部位由褐色逐漸至黑色。復眼1 對,單眼3 只,位于額頂兩復眼中間,呈倒三角形排列;初期單、復眼顏色與蟲體相近,約2 d 后變為淺粉色,并逐漸由淺紅色轉紅色,再由紅色轉暗紅色后轉黑色。整個蛹期約6-9 d,平均7.4 ±0.84 d (n=10),羽化完成后變成成蟲,新成蟲打通蟲室隔離層及前廳堵塞物,爬出巢口。

圖版Ⅰ 日本佳盾蜾蠃的形態特征PlateⅠ The morphology of Euodynerus nipanicus
2.2.2 成蟲的羽化及性比
利用人工巢管筑巢的日本佳盾蜾蠃在江西贛州地區每年4月底至5月初開始陸續羽化出巢,雄蜂先于雌蜂1-2 天羽化出巢,出巢后在巢區附近抖動雙翅清理蟲體等待其它雌蜂羽化出巢。越冬代成蟲從4月底開始羽化,5月初羽化數量逐漸上升并達到高峰,至5月中旬開始數量逐漸降低,其他世代的羽化時間見表1。成蜂羽化時間主要集中在晴好天氣下的上午7∶00-9∶00,其次是下午15∶00-17∶00 時。
日本佳盾蜾蠃不同世代的性比略有不同,全年共收集巢管200 枝,累計羽化出巢1020 頭,其中雌蜂762 頭,雄蜂258 頭。通過表2 可以看出,不同時期收集的巢管,其出蜂的雌雄比有一定的差異,其中5月15 收集的巢管,其出蜂的雌雄比最大,為3.67∶1;雌雄比最小的是在8月15 日收集的巢管,僅為2.31∶1;10 次收集的巢管成蟲羽化后的平均性比為2.95∶1。
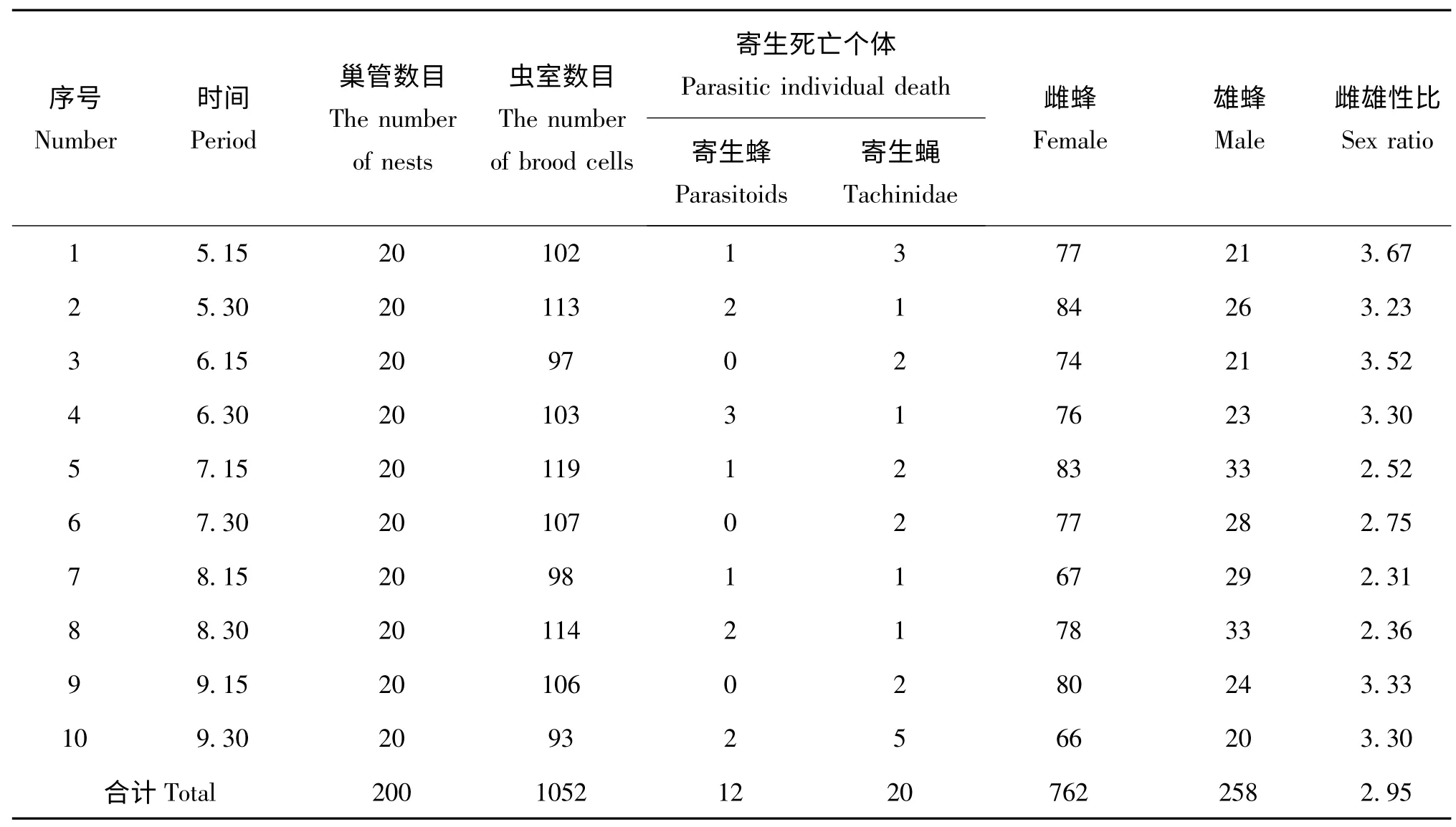
表2 日本佳盾蜾蠃的性比及寄生情況(江西,贛州)Table 2 The sex ratio and parasitism of Euodynerus nipanicus (Schulthess)(Ganzhou,Jiangxi)
2.2.3 成蟲的交配行為
成蟲在羽化出巢當天即可進行交配,交配率以羽化后次日上午最高,4 d 后顯著下降。晴好天氣下,該蜂全天均可進行交配,交配高峰一般在上午的8∶00-10∶00 之間和下午的16∶00-18∶00之間。雄蜂羽化較雌蜂早且主要集中在巢口附近的雜草灌木周圍或是蜜源植物附近等待羽化出巢的雌蜂,尋找交配機會,一旦發現雌蜂,雄蜂立即撲上去與之交尾。交尾時雄蜂爬到雌蜂體背用上顎和足抓住雌蜂,不斷用前足撥動雌蜂的前胸背部,待雌蜂腹部末端上翹接受交尾時,雄蜂伸長并向下彎曲腹部,翻出陽莖,插入雌蜂的生殖孔。交配時間在60-250 s 之間,平均141.1 ±46.2 s (n=10)。一頭雄蜂可先后可與多頭雌蜂交配,而雌蜂一生只接受一次交配。
2.2.4 成蟲的營巢活動
雌蜂羽化出巢后尋找蜜源植物補充營養,期間完成交配。交配后的雌蜂在羽化巢區附近選擇合適的巢管并清理好巢管后,開始采集濕潤的泥土筑巢。筑巢時從巢管的末端開始,一般按照構筑末端預封塞物(deep plug)、構建蟲室(brood cell)、產卵、準備蜂糧、構建室間隔離層 (cell partition)、構建空室 (deep cell)構建下一個蟲室、最后用泥塊密封管口意味前庭 (vestibular cell)構筑完成。雌蜂在晴好天氣下每天出巢次數在45-66 之間,平均55 ±5.33 次(n=10),其中捕食次數約26 次,采集泥塊次數約21 次,其他為外出采集花蜜補充自身營養或是采集不成功。雌蜂晚上棲息于距巢口1-2 cm 主道內。
雄蜂羽化出巢后尋找蜜源植物補充營養,白天潛伏在巢口附近的雜草灌木周圍或是蜜源植物附近尋求交配機會,不參與新巢的構建和蜂糧的采集。在同一個巢管雌蜂沒有羽化之前,雄蜂夜宿巢管并幫助打通蟲室通向巢口的隔離層;在雌蜂筑巢期間,雄蜂一般不夜宿自己羽化的巢管和雌蜂新構建的巢管,而是尋找廢棄的老巢或是雜草灌木的葉面背部夜宿。雄蜂壽命短于雌蜂,一般只有15 d 左右。
2.2.5 蜜源植物
日本佳盾蜾蠃為捕食性天敵昆蟲,雌蜂捕食鱗翅目昆蟲幼蟲作為繁育后代的食物。同時,該蜂也為雜食性昆蟲,觀察表明,該蜂訪問紫藤Wisteria sinensis Sweet、黃 荊Vitex negundo Linn.、紅蓼Polygonum orientale Linn.、野棉花Anemone hupehensis Lemoine.(黃敦元等,2013)、鹽膚木Rhus chinensis Mill.、空心菜 Ipomoea aquatica Forsk、野棉花Anemone hupehensis Lemoine.等植物的花朵,取食花蜜補充自身營養。
日本佳盾蜾蠃雌性個體的額、顱頂、胸部等部位覆有短毛,其訪花主要目的是取食花蜜維持個體生存需要,在訪問蜜源植物花時,前足、中足攀附在花萼或花枝上,將頭伸入花柱底部取食花蜜,在取食花蜜過程中常會對花部結構造成一定的破壞,但同時體壁上會粘附少量花粉,對到訪植物可能具有一定的傳粉功能。
2.2.6 寄生性天敵
通過對定期收集回實驗室的巢管進行室內飼養和觀察,在200個巢管中,累計有1052個蟲室,每個蟲室有1 頭日本佳盾蜾蠃幼蟲,因寄生而死亡的幼蟲有32 頭,其中有12 頭幼蟲被寄生蜂寄生,20 頭幼蟲被寄生蠅寄生(表2)。本研究中涉及的寄生蜂和寄生蠅具體物種需要進一步鑒定。
3 結論與討論
3.1 雌雄比在不同世代之間的差異
巢管營巢的獨棲野生蜜蜂和胡蜂類群,其不同世代雌雄比的差異主要受以下因素的影響:首先是巢區附近食物資源的豐富度是影響雌雄比的主要因素之一 (Jason et al.,2006;Peterson and Roitberg,2006),Jason 等 (2006)研究顯示:Megachile rotundata (F.)巢區附近食物資源的豐富度通過影響其后代個體幼蟲期的食物多少而決定其后代的雌雄比;其次是受到雌蜂所選擇巢管的規格及制作蟲室的大小影響 (Carlo et al.,2011;Fateryga,2013),Fateryga (2013)通過野外觀察和解剖Euodynerus quadrifasciatus (F.)的巢管提出:巢管中單個蟲室大小是影響不同蟲室中卵和幼蟲的發育情況進而影響到個體大小和雌雄性比,因為蟲室的大小制約蟲室中幼蟲發育所需的食物數量;再次是季節的更替和溫度的變化也是導致雌雄個體差異及性比不同的因素之一(Fye,1965;Longair,1981),Fye (1965)通過巢管誘集大量Rygchium leucomelas (Sauss.)并對其生物學進行研究發現,該蜂一年2 代,越冬代雄性個體的數目偏多,夏季一代的個體雌性個體偏多。本研究中的日本佳盾蜾蠃在每年的第1 代和最后1 代的雌雄比比較大,7-8月份羽化出巢的第3 代雌雄比明顯偏小,我們初步認為該時期的食物資源緊缺和巢區內的溫度過高是導致這種現象的主要因素,有待于進一步的驗證。
3.2 巢室內成蜂羽化順序
雌蜂在營巢構建蟲室時,發育成雄性成蟲的蟲室一般在最外層的幾個蟲室中。雄蜂先于雌蜂1-2 d 羽化,羽化后雄蜂打通蟲室通向巢口的室間隔離層外出取食,雌蜂沒有羽化之前回巢管過夜,等待后續羽化的雌蜂并幫助及時打通巢管末端的室間隔離層。雄蜂是如何確定雌蜂的羽化我們目前還尚不清楚,初步推測是雄蜂聽到巢管蟲室內雌蜂羽化后翅膀產生的振動以便確定雌蜂的羽化。
References)
Bruun HH.Patterns of species richness in dry grassland pat ches in an agricultural landscape[J].Ecography,2000,23,641-650.
Carlo P,Roberto B,Walter B.Few,small,and male:multiple effects of reduced nest space on the offspring of the solitary wasp,Euodynerus (Pareuodynerus)posticus (Hymenoptera:Vespidae)[J].Comptes Rendus Biologies,2011,334 (1):50-60.
Carpenter JM.The phylogenetic relationships and natural classification of the Vespoidea (Hymenoptera)[J].Systematic Entomology,1982,7 (1):11-38.
Cowan DP.The solitary and presocial Vespidae.In Ross KC,Matthews RW,eds.The Social Biology of Wasps[C].Ithaca and London:Comstock Publishing Associates,1991,678.
Fateryga AV.The nest structure in four wasp species of the genus Euodynerus Dalla Torre (Hymenoptera,Vespidae:Eumeninae)[J].Entomological Review,2013,99 (4):412-421.
Fye RE.The biology of the Vespidae,Pompilidae and Sphecidae(Hymenoptera)from trap nests in northwestern Ontario [J].Canadian Entomol.,1965,97 (7):716-744.
He CL,Ji BZ,Liu SW.Morphology and biology of Xylocopa tranquebarorum[J].Chinese Journal of Applied Entomology,2011,48 (6):1751-1758.[賀春玲,嵇保中,劉曙雯.長木蜂的形態和生物學觀察[J].應用昆蟲學報,2011,48 (6):1751-1758]
Huang DY,He B,Yu JF,et al.Nesting biology of Euodynerus nipanicus Schulthess (Hymenoptera:Vespidae)[J].Journal of Environmental Entomology,2013,(35)6:778-787.[黃敦元,何 波,余江帆,等.日本佳盾蜾蠃營巢生物學研究[J].環境昆蟲學報,2013,35 (6):778-787]
Jason HP,Bernard DR,Peterson JH.Impacts of flight distance on sex ratio and resource allocation to offspring in the leafcutter bee,Megachile rotundata [J].Behavioral Ecology and Sociobiology,2006,59 (5):589-596.
Ju Y,Chen B,Li TJ.Two new species of the genus Ancistrocerus Wesmael (Hymenoptera:Vespidae:Eumeninae) from China,with a key to the Oriental species[J].Zookeys,2013,303:77-86.
Kim JG,Choi YC,Choi JY,et al.Environmental evaluation by using Hymenoptera induced by Bamboo Pipe Traps indicated by Eumenid wasp (Hymenoptera:Eumenidae)[J].Korean Journal of Applied Entomology,2005,44:307-315.
Kim JK,Yamane S.A revision of Eumenes Latreille (Hymenoptera:Vespidae)from the Far East Asia,with descriptions of one new species and one new subspecies[J].Entomological Science,2001,4 (2):139-155.
Kim JK.Taxonomic Review of the genus Euodynerus (Hymenoptera:Vespidae:Eumeninae)in the Korean Peninsula[J].Anim.Syst.Evol.Divers.,2012,28 (3):161-167.
Peterson JH,Roitberg BD.Impact of resource levels on sex ratio and resource allocation in the solitary bee,Megachile rotundata [J].Environmental Entomology,2006,35 (5):1404-1410.
Lee TS.Wasps in Rural Region of China[M].Beijing:Agricultural Publishing House,1982,64-115.[李鐵生.中國農區胡蜂.北京:農業出版社.1982,64-115]
Lee TS.Economic Insect Fauna of China,Fasc.30,Hymenoptera:Vespoidea[M].Beijing:Science Press,1985,1-159.[李鐵生.中國經濟昆蟲志·第三十冊· 胡蜂總科[M].北京:科學出版社,1985,1-159]
Longair RW.Sex-ratio variations in xylophilous aculeate Hymenoptera[J].Evolution,1981,35:597-600.
Zhou X,Chen B,Li TJ.Two new species of the genus Discoelius Latreille(Hymenoptera:Vespidae:Eumeninae)from China,with a key to the Chinese species[J].Journal of Hymenoptera Research,2013,32:45-54.
Yamane S.A revision of the Japanese Eumenidae (Hymenoptera,Vespoidea)[J].Insecta Matsumurana,New Series,1990,43:1-189.
Zhou X,Chen B,Li TJ.Two new species and a key to species of the genus Eumenes Latreille (Hymenoptera:Vespidae:Eumeninae)from southwestern China[J].Entomotaxonomia,2012,34 (2):467-474.
Zhou X,Li TJ,Chen B.The Taxonomic research progress of Eumeninae(Hymenoptera:Vespidae)[J].Journal of Chongqing Normal University (Natural Science),2011,28 (6):22-29.[周鑫,李廷景,陳斌.蜾蠃亞科(膜翅目:胡蜂科)分類研究進展[J].重慶師范大學學報(自然科學版),2011,28 (6):22-29]
