長江口張網魚類群落結構特征及月相變化
2014-10-27 05:37:12史赟榮晁敏沈新強
海洋學報 2014年2期
史赟榮,晁敏,沈新強*
(1.中國水產科學研究院 東海水產研究所,上海200090)
1 引言
河口對棲息在其中的很多生物種類有著重要的作用,例如,河口不僅是很多經濟魚類的育幼場[1-3],也是洄游性魚類的洄游通道[4]。魚類群落是河口生態系統的重要組成部分[5],同時又是重要的生物資源[6]。長江口是中國最大的河口生態系統,河口魚類群落生態學研究也是國際熱點研究之一[7]。關于長江口魚類群落生態學的研究,有學者側重于魚類資源學的角度分析了其時間結構特點[8-9],也有學者從季節變動的角度分析了群落的變化[10],還有學者分析了夏秋季長江口張網魚類群落的結構特征[11]。張網是長江口漁業捕撈主要作業方式[12],目前尚未見有1周年中12個月張網連續采樣的報道,而關注魚類群落的時間格局,了解群落組成(如一些主要經濟種群)的時間變化,可以為合理管理和利用魚類資源提供依據。基于以上考慮,本研究通過對長江口2010年5月至2011年4月總共1周年12個月的張網魚類調查,分析了其群落組成特點及月度時間結構特征,以期為更好地保護和利用長江口魚類資源提供依據。
2 材料與方法
2.1 采樣方法
調查網具為定置張網,作為一種被動過濾型定置作業網具,其技術要求較低,可利用潮水漲落進行捕撈作業,因而作業在漲落潮或晝夜時均可進行。本研究中采用的網具寬度為4.2 m,高1.3 m,網身長6 m,網囊網目約為2.5 mm,每次作業時間(投網至起網)約為24 h(1晝夜)。調查水域位于30°48.50′~30°59.60′N,122°02.00′~122°07.19′E區域內(圖1),該區域也是上海市張網傳統作業水域。調查時間為2010年5月至2011年4月,共12個月,在12個月中,5月采樣2次,共4網,6月至9月均為每月4次(農歷大小潮各2次),而10月至第2年1月及3月均為每月2次(農歷大小潮各1次),而2月和4月采樣次數均為1次(農歷大潮)。一般每次隨機抽取2網全部漁獲物并測定采樣站位的鹽度和水溫,而每月徑流量以網上公布大通站流量為準[13]。起網后,現場將漁獲物定種、稱量、計數。
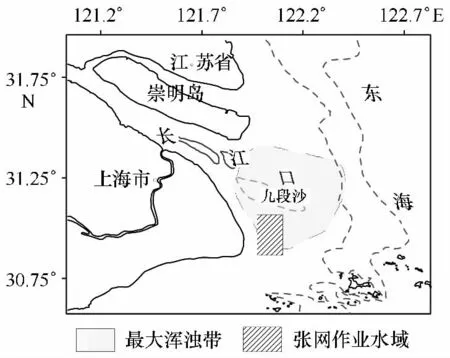
圖1 2010年5月至2011年4月張網作業水域
2.2 研究方法與數據處理
魚類群落多樣性分析采用Shannon-Wiener多樣性指數及Si mpson優勢度集中指數,相似性分析用Bray-Curtis相似性系數。
Shannon-Wiener多樣性指數:
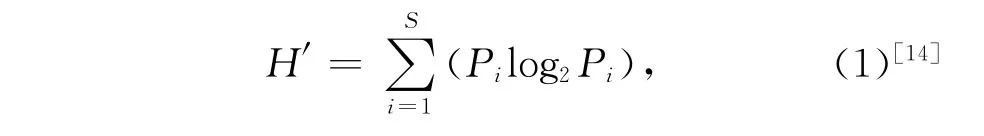
Simpson優勢度指數:
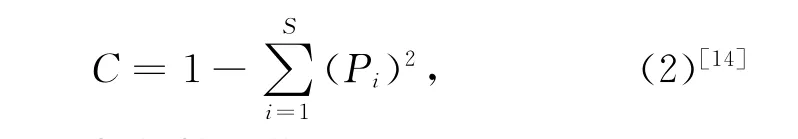
Bray-Curtis相似性系數:
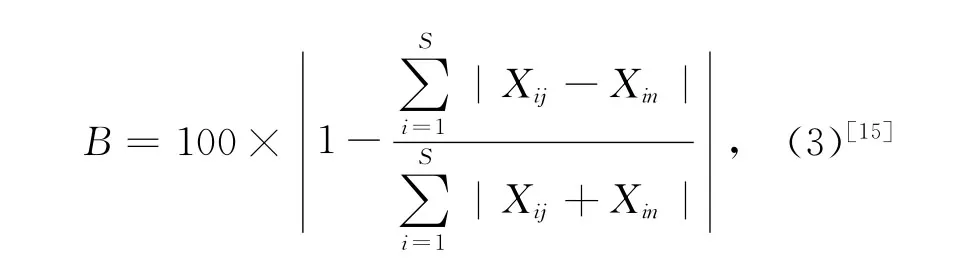
式(1)和(2)中,S表示每網中總共出現的物種數目,Pi表示第i個物種豐度在每網豐度中的比例;式(3)中,當表示各網樣品的相似性時,Xij和Xin分別為第i個種類在第j網和第n網期的豐度(經4次方轉化),S為魚類物種數目;當表示各月樣品的相似性時,Xij和Xin分別為第i個種類在第j個周期和第n個周期(月)的平均豐度(經4次方轉化),S為魚類物種數目。
魚類分類系統及學名分別以Nelson分類系統[16]及Fishbase為準[17]。在調查中出現的各種魚類生態類群的劃分以Elliott和Dewailly為準[3]。本研究中的生態類群包括海洋洄游性魚類(Marine migrants,MM),海洋偶見性魚類(Marine stragglers,MS),河口性魚類(Estuarine species,E),半溯河洄游性魚類(Semi-anadromous,SA),溯河洄游性魚類(Anadromous,A),降海洄游性魚類(Catadromous,CA)及淡水偶見性魚類(Freshwater stragglers,FS)等。絕大多數魚類種類隸屬的生態類群依有關文獻報道[18-22],個別尚未定義隸屬生態類群的物種以Fishbase為準[17]。魚類區系適溫類型的劃分見有關文獻[23-24],魚類棲息水層的定義及劃分則以有關文獻[23]及 Fishbase為準[17]。
用Fried man檢驗分析月度間各生態類群豐度或生物量貢獻率的顯著性差異,用非參數獨立樣本(非正態數據)或F檢驗(正態數據)分析各月物種數目、豐度、生物量、H′和C以上各統計檢驗顯著性水平為p<0.01(極顯著差異)、p<0.05(顯著差異)兩個水平。用非度量多維尺度法(Non-metricmultidimensional scaling,NMDS)分別分析每網及每月(取月平均值)樣本中魚類群落結構的差異,此外用相似性比例分析來分析組內魚類群落的相似性主要由哪些種類引起,同時用相似性程序分析檢測魚類群落不同月份組之間的顯著性差異,計算用PRIMER 5.2統計軟件包處理[25]。最后,用多元回歸分析月平均豐度、種類數目或生物量(均取對數)與月平均水溫、鹽度及徑流量的關系,計算采用 DPS 2000軟件處理[26]。
3 結果
3.1 群落組成
調查期間總共記錄1綱(硬骨魚綱)13目31科56種魚類。在目分類水平上,以鱸形目種類數目最多(27種),其次鯡形目包含的種類數目也較多(10種),而鯉形目、頜針魚目、刺魚目、仙女魚目、燈籠魚目、鯔形目等目只包含1種魚類;在科分類水平上虎魚科包含種類數目最多(8種),其次為鳀科(7種),而單種科數量較多,共有23科。按照魚類適溫類型,只出現暖溫性及暖水性2種適溫類型,其中暖溫性魚類31種,暖水性魚類25種。根據棲息水層的劃分,底棲魚類種類數目最多(26種),中上層魚類最多(21種),底層魚類最少(9種)。
圖2給出了豐度及生物量的累積曲線,由圖2可知豐度排序較高的前5種魚為鳳鱭、龍頭魚、棘頭梅童魚、尖海龍及康氏小公魚。而生物量比例最高的前5種魚類種類組成與豐度排序有所不同(圖2),在生物量排序種類中,龍頭魚、鳳鱭及棘頭梅童魚繼續保持優勢地位,但尖海龍及康氏小公魚的優勢地位由矛尾鰕虎魚及斑鰶替代。不過,無論是豐度還是生物量,均由少數魚類占據絕對優勢(圖2)。生態類群中按種類數目的多少,海洋洄游性魚類(21種)及河口性魚類(19種)比例最高;海洋洄游性魚類明顯占據豐度(58.05%)或生物量(70.09%)的絕對優勢。按體型大小,生物量及豐度均呈現出由小型魚類占據主導的格局。
3.2 種類數目、豐度及生物量的時間變化
月平均物種數目最高值出現在7月份(18.3種),此外4月份物種數目也較高(17種),而最低值則出現在1月份(7.2種)(圖3a),時間變化明顯(H=572.87,p<0.01)。豐度在月度上也有明顯的變化(H=726.34,p<0.01),變化趨勢總體上呈現先增加后急劇降低后又升高的特點,其中最高值出現在11月份(2 272尾/網),而最低值出現在冬季1月(71尾/網)(圖3b)。月平均生物量也有明顯的時間變化(F=11.08,p<0.01),其中生物量在2011年2月前變化趨勢大體與豐度變化一致,但2月后生物量總體呈現下降趨勢(圖3c),最高值(11 892 g/網)和最低值(351 g/網)分別出現在9月和1月。

圖2 2010年5月至2011年4月魚類豐度及生物量的K-優勢度曲線
Fried man統計檢驗結果顯示調查期間各月生態類群豐度或生物量比例均有極顯著差異(p<0.01),這說明不同月份魚類群落中不同生態類群對豐度或生物量的貢獻率是不一樣的。如根據不同類群對月平均豐度和生物量的貢獻率的差異可看出5—12月大體上均以海洋洄游性魚類占據優勢,而在1—4月占優勢的生態類群根據月份而呈現較大差異,如1月無論是豐度還是生物量,均以溯河洄游性魚類占據優勢,而2月均以河口性魚類占據優勢,然而3—4月豐度以半溯河洄游性魚類占據優勢,3月河口性魚類生物量仍占優勢,不過4月其優勢地位已被半溯河洄游性魚類所替代(見圖4)。

圖3 2010年5月至2011年4月長江口魚類群落種類數目(a)、豐度(b)及生物量(c)的時間變化
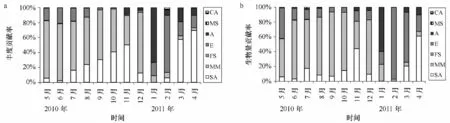
圖4 2010年5月至2011年4月生態類群對豐度(a)及生物量(b)貢獻率的時間變化
3.3 主要種類豐度的時間變化
調查期間總共有9種魚類占總豐度的比例在1%以上,其豐度在不同月份有明顯差異(圖5)。如鳳鱭豐度在秋季(9—11月)達到峰值,以11月最高;龍頭魚豐度高值區分布在夏至初冬季(8—12月),最高值出現在9月份;棘頭梅童魚豐度以5—9月較高,又以7月豐度最高;尖海龍豐度最大值出現在11—12月,而其他月份豐度很小甚至出現豐度為0的情況;康氏小公魚豐度最高值出現在9月,此外5月份豐度也較高,豐度低值主要出現在冬季月份;刀鱭豐度最高值出現在11月份,而春末至初秋月份的豐度均較低;鯔魚和帶魚豐度高值月份主要出現在夏季,而冬季豐度相對較低;紅狼牙鰕虎魚的豐度在春夏季較高(6月最高),冬季較低。
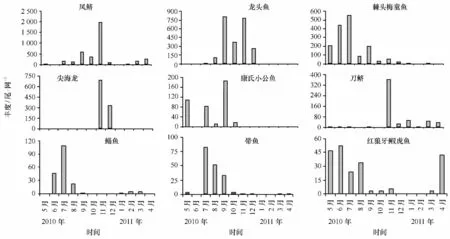
圖5 2010年5月至2011年4月主要種類豐度的時間變化
3.4 群落多樣性指數的時間變化
調查期間每網Shannon-Wiener多樣性指數(H′)與Simpson優勢度集中指數(C)有極顯著正相關性(R2=0.92,p<0.01)。H′指數在1年中不同月份有極顯著差異(H=27.731,p<0.01),其中月平均值以7月最高(2.70),同時8月也較高(2.62),而最低值分別出現在6月(1.44)和11月(1.46)(見圖6),C指數的月分布也有統計學差異(H=25.133,p<0.01),其月平均值變化趨勢與H′指數基本一致,最大值分別出現在8月(0.77)和7月(0.66),而最小值出現在6月(0.44)和11月(0.47)(圖6)。
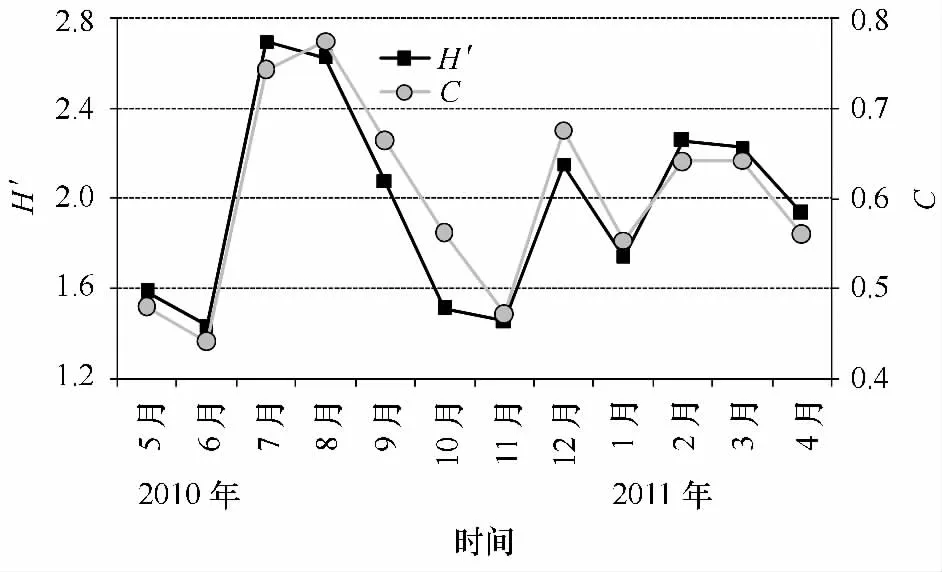
圖6 2010年5月至2011年4月Shannon-Wiener指數(H′)與Si mpson指數(C)的時間變化
3.5 群落月度動態的NMDS分析
由圖7a可知調查期間魚類群落按照每網樣品采樣日期基本可以分為4組(第Ⅰ組:1—4月;第Ⅱ組:5—8月上半月,第Ⅲ組;11月下半月—12月;第Ⅳ組:8月下半月—11月上半月)。根據圖7b還可知一般相鄰月的群落相似性較高,但是1月與12月之間魚類群落相似性距離較大,同時2010年5月與2011年4月相似性也較小。另外,每組的特征種也有較大差異,每組均出現的特征種只有棘頭梅童魚、刀鱭及鳳鱭3種魚類(表1)。表2還給出了不同組間的區別種,由該表可知不同組間的區別種也有一定差異。
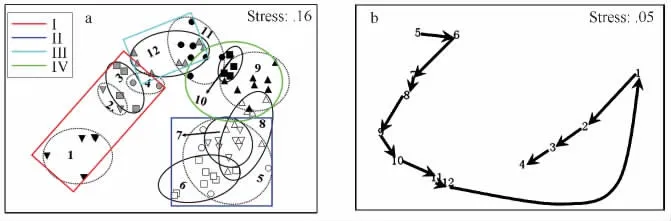
圖7 魚類群落的NMDS分析
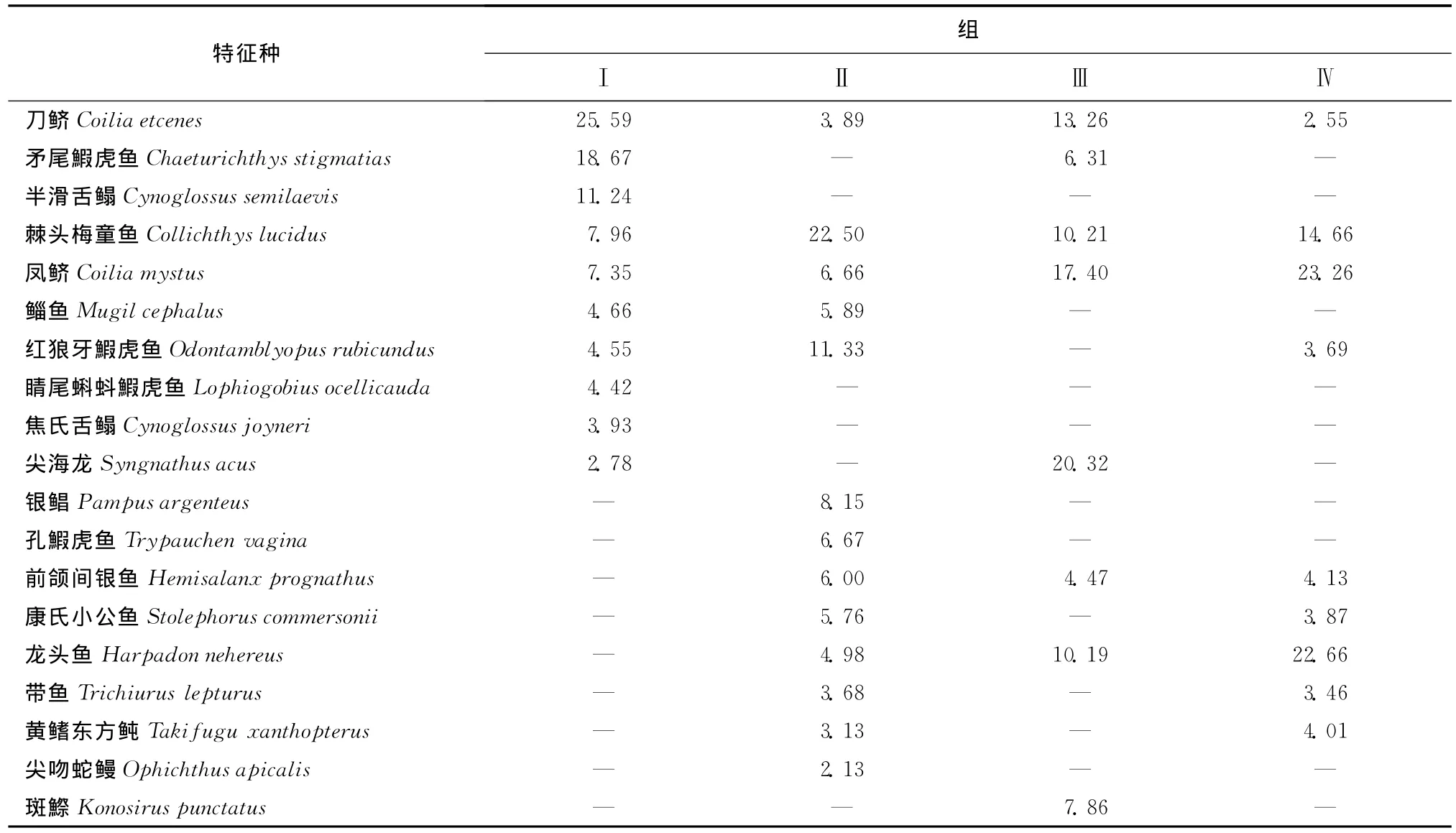
表1 各組內的特征種(對相似性的貢獻率,%)
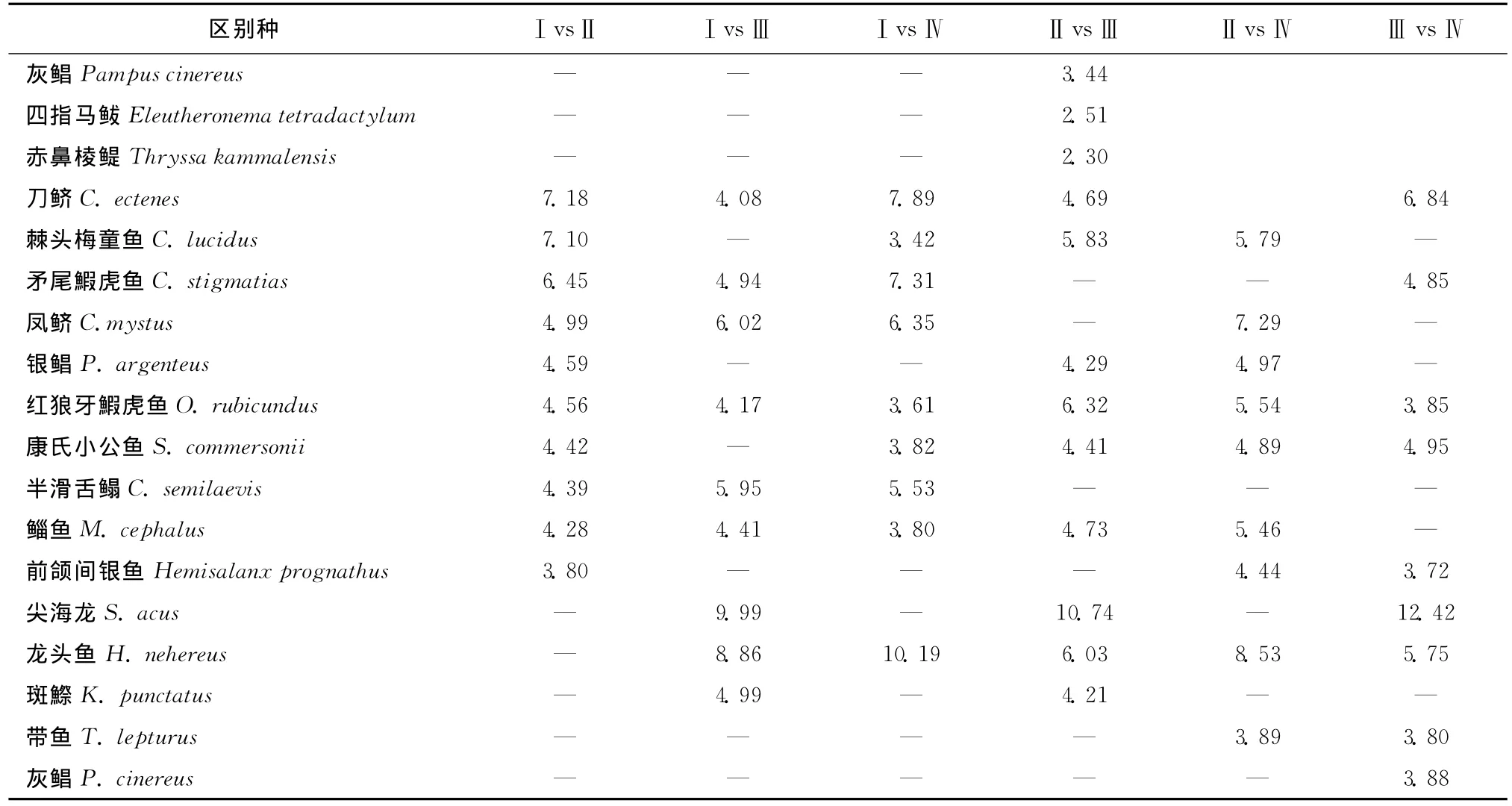
表2 各組間的區別種(對相異性的貢獻率,%)
3.6 魚類群落與環境因子的多元回歸分析
多元回歸模型顯示了所選擇的3個環境因子(鹽度、水溫、流量)與月豐度、生物量或種類數目的多元回歸關系,只有豐度與環境因子的多元回歸關系解釋得較好,說明豐度變化與所選擇的環境因子有密切的關系,且68%(R2=0.68)的信息量可以被列入分析的3個環境因子(鹽度、水溫、流量)所解釋,但其中只有水溫為顯著相關影響因子(正相關)(表3),這說明水溫越高的月份,一般魚類豐度也較高。
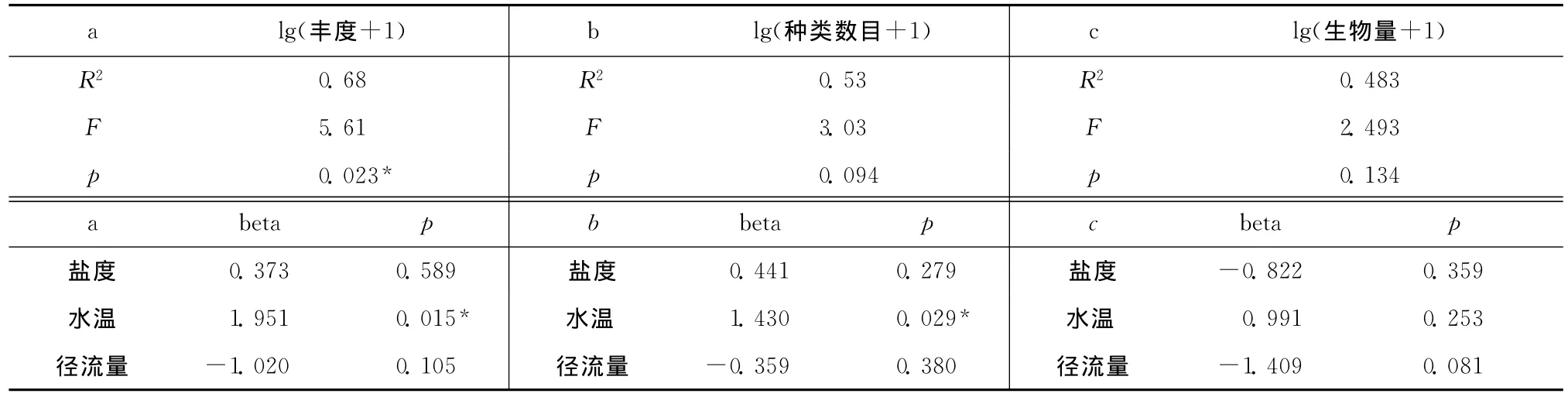
表3 月平均豐度(a)、種類數目(b)及生物量(c)與環境因素(鹽度、水溫、流量)的多元回歸關系
4 討論
張網是一種能較好地捕獲各種棲息水層魚類的作業方式[27],因而能較為客觀地反映作業水域魚類群落的狀況。如本研究中,張網對于底棲和中上層魚類均具有較高的捕獲率。此外由于長江口位于中低緯度的亞熱帶水域,其魚類群落區系類型表現出典型的暖水性(25種)及暖溫性魚類物種(31種)占據的特征。
4.1 與其他河口魚類群落生態類群的比較
長江口海洋魚類種類數目的比例與北半球溫帶河口區魚類[28]及中國其他亞熱帶河口[29-30]類似。然而,由于長江口有較高比例的河口性魚類(33.93%),這與其他河口區的狀況有所不同,如法國Canche河口[31],德國Elbe河口[32],土耳其 Koycegiz河口[33],而位于烏拉圭和阿根廷之間的Rl'o de la Pla-ta河口甚至沒有記錄河口性魚類[34],但是在葡萄牙Mondego河口[35],西非岡比亞河口[36]以及塞內加爾Sine Salou m河口[37],河口性魚類的比例均較高。當然,魚類生態類群的劃分也并不是絕對的,例如同一種魚在不同的河口中其生態類群就有可能不同,例如棘頭梅童魚在珠江口是河口性魚類[38],但在本研究中卻是海洋洄游性魚類。
強潮河口魚類群落中河口性魚類的豐度比例往往很低,例如,在英國Severn河口,河口性魚類的數量只有0.7%[39]。這很大程度是由于強潮河口強大的水動力特征(如潮汐影響)使得河口性魚類的卵和稚魚(一般其無運動能力或者運動能力很差)無法長期存在河口中,同時,如果這些魚類的魚卵、仔稚魚一旦離開了河口,其存活就會受到很大的威脅,因而也就無法在河口完成生活史循環[40]。然而,潮汐對河口性魚類分布的影響也是相對的,例如,Nicolas等[41]進一步指出即使像Loie及Seine河口這樣的強潮河口,河口性種類的豐度仍然較高,而在南非一些潮汐較弱河口中河口性魚類的豐度卻較低[7]。因而從這個角度來看潮汐并不是影響河口性魚類豐度比例的唯一因素。正如一些學者所指出的那樣,潮間帶對其所在河口的魚類豐度也有正面影響,尤其是對海洋和河口性魚類的棲息密度[41-45]。長江口是一個中等強度潮汐的河口 (2.66 m),并且有可供大量幼魚索餌和躲避捕食的較大面積的鹽沼生境 (213 km2)[45]。不過,張網采樣反映出河口性魚類的豐度比例(7.7%)并沒有底拖網那么高[46],這可能是由于張網調查水域位于長江口最大渾濁帶內[47],而一些河口性魚類如矛尾鰕虎魚卵的孵化很可能會因此受到影響,因為水體渾濁度過高可能會對魚卵的存活、孵化過程、攝食效率、生長率和種群大小產生負面影響[48],也有研究認為一些海洋性魚類往往偏好渾濁度較高的水域,會游動至河口渾濁的水體來躲避肉食魚類的捕食[49]。
4.2 魚類群落的優勢種
河口魚類群落豐度往往由少數幾種魚類占據絕對優勢,這與河口溫鹽等環境因子分布梯度較大有關[15],本研究中魚類群落也體現出類似的特征。河口水域魚類區系這種種類數目相對較低但是個體豐度較高的特點反映出生活在河口的一些魚類對河口的環境波動有較好的適應能力[50],因而這些魚類比那些無法忍受環境劇烈變化的種類具有明顯的優勢,這是因為前者能夠更好地利用河口充足的資源(如食物),而其他潛在的競爭者卻因為不能適應這種波動的環境而無法大量出現[51]。如本研究中豐度比例較高的的棘頭梅童魚[52]、鳳鱭、刀鱭等對溫鹽就有較好的適應性。然而這些種類的豐度時間分布格局卻有較大差異,如棘頭梅童魚豐度在夏季的7月最高、刀鱭和鳳鱭等魚類在11月最高。這種魚類群落優勢種在時間上的錯峰分布現象在其他河口也有發現[53],錯峰分布反映出不同物種對生境利用的差異,同時也可能與減少優勢種之間的競爭行為有關。
4.3 魚類群落的多樣性特征
多樣性參數的計算還可以以生物量為基礎進行計算,但在本研究中,采用了基于豐度為基礎計算的多樣性參數,這是因為若從資源保護的角度考慮應以個體數為基礎估算,若從漁業經濟開發考慮則應以生物量為基礎進行分析[54]。鑒于長江口魚類資源已經處于衰退狀態的局面[55],從魚類群落的管理和保護角度出發可能以個體為基礎進行分析更為合理。
群落種類多樣性水平的高低由群落的物種數目及各物種個體比例所決定,一般物種數目較豐富且個體比例較均勻的群落,其多樣性值一般較高[56]。本研究中,魚類群落H′(1.44~2.70,平均值為1.97)和C(0.44~0.77,平均值為0.61)低于緯度更低的采用定置張網調查捕獲魚類尾數計算(24°26′46″N)的廈門東海域魚類群落多樣性參數H′(1.56~4.34,平均值為3.14)和C(0.68~0.94,平均值為0.80)[56],但與同緯度的黃龍島海域魚類群落H′(1.24~2.52,平均值為1.94)和C(0.40~0.84,平均值為0.61)接近[27]。陳國寶等[54]也發現緯度更低的南海珊瑚礁魚類群落H′指數比其他較高緯度海域魚類群落H′高。這種多樣性參數的緯度分布差異反映出緯度較低的海域魚類群落種類的個體比例一般更均勻,所以多樣性參數一般較高。而緯度越低的海域,全年生態環境(如溫度、鹽度)的變化一般也更為穩定,對大多數魚類的分布限制相對較小,因而很少有物種的個體數量能夠占據絕對優勢。
4.4 魚類群落與環境因子的關系
不同種類的魚類耐溫性有所差異[57]。長江口魚類種類數目與豐度變化趨勢基本一致,總體上溫度較高月份記錄的種類數目較多和豐度較高,這是因為氣溫較高月份大量的海洋魚類進入河口索餌、育幼所致。在萊州灣[58]、Mad Island鹽沼河口[59]、Koycegiz Lagoone河 口[33]、Acquatina[60]均有此發現。而 在Pantan和Zr manja河口[61-62],種類數目在春季最高,而豐度在冬季最高。不過在Porto-Lagos潟湖[63],種類數目和豐度的季節變化并不明顯,此外在溫帶的Canche河口[31],種類數目卻以春季(5月)最低、秋季(11月)最高。
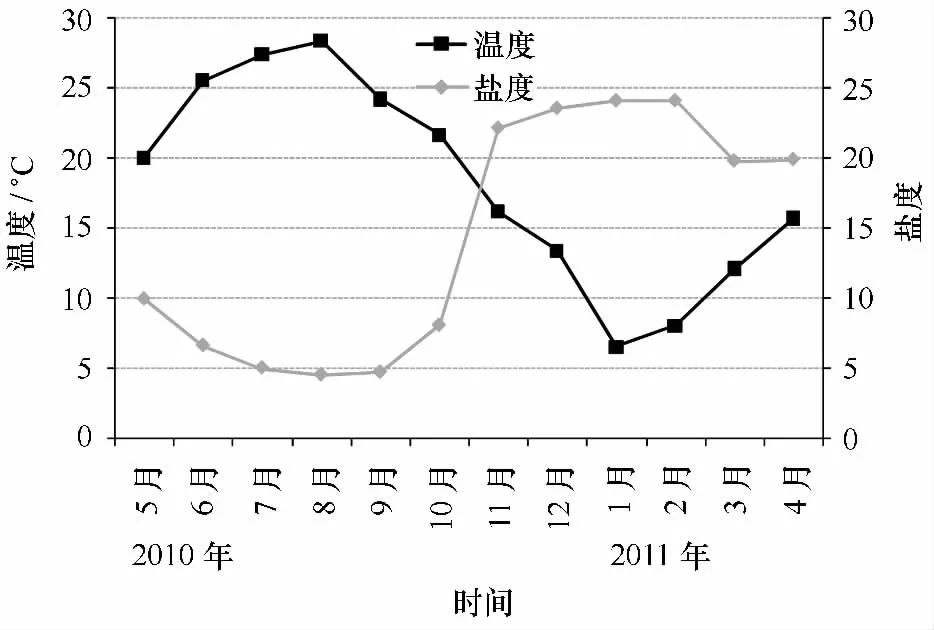
圖8 2010年5月至2011年4月水溫、鹽度的時間變化
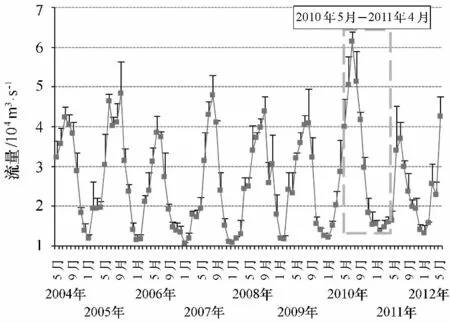
圖9 2004年5月至2012年5月長江口月徑流量分布
通過魚類群落結構月度更替可以發現,總體上相鄰月份的群落結構相似性較近。在珠江口1998年1—12月相鄰月份魚類群落相似性也較高[38]。然而,2010年12月與2011年1月群落結構有較大的差異,朱鑫華等[64]也發現黃河口魚類群落結構在1月與12月之間相差較大。1月魚類群落的急劇變化可能跟環境條件特別是水溫的下降有關系。這是因為生物對環境因子的適應都有一個耐受限度,生物只能在耐受限度規定的生態環境中生存[65]。冬季豐度的低值反映出環境因子(主要是水溫)對魚類分布的限制,低溫是導致冬季物種數目及豐度均較低的主要原因。如1月平均水溫僅為7℃左右(圖8),這個溫度已經低于長江口一些海洋魚類的最低耐受溫度,如夏秋季大量分布的龍頭魚(最低耐受溫度為8℃),其分布明顯受冬季的低溫限制[17]。除了龍頭魚,鳀魚的分布也明顯受到低溫的限制,研究發現鳀魚最適生存溫度為8~30℃[17],因而當冬季長江口水溫在8℃以下時鳀魚一般會離開河口水域洄游到水溫較高(一般大于10℃[66])的鄰近海域進行越冬。Laffaille等[67]也有類似發現。當然,有時高溫和低溫均會對魚類的分布產生影響,如海洋洄游性魚類尖海龍,在水溫較高月份(6—8月)及水溫較低的1—3月份,其豐度基本為0。除了水溫,鹽度也是一個影響河口魚類群落的重要影響因子(主要影響空間結構[31]),但由于本研究中為定點調查,調查水域沒有包括淡水生境(鹽度小于0.5)及海洋生境(鹽度大于30),1年中水體鹽度梯度相對較小(4.5~24.1)(圖8),并且占群落豐度絕對優勢的鳳鱭、刀鱭及棘頭梅童魚由于對鹽度的適應范圍很廣(能適應鹽度小于0.5的水體),因而鹽度對總豐度的影響并不如預期那樣大,這可能是溫度對魚類群落的影響權重要高于鹽度的主要原因,同時也有學者發現即使在長江口鄰近水域(采樣區域沒有包括淡水及低鹽水體(鹽度小于5)),鹽度也不是一個影響魚類群落結構的主要因子[68]。不過,在熱帶水域,由于一年中水溫變化較小,水溫對魚類群落的影響有限[36]。此外,雖然2010年5月與2011年4月水溫值較為接近,然而在NMDS排序圖上魚類群落距離卻相差較大,這反映出魚類群落很可能存在著年際間的變動,而這種變動是由于年際間環境因子的差異引起的。長江口環境是一個受長江徑流和海洋環境影響的水域,徑流的改變會通過鹽度、懸浮物甚至營養鹽的變化從而影響到魚類群落結構,2011年春季長江流域由于極端氣候影響導致長江流量急劇下降,這進一步影響到了長江口的生境條件(如5月與第2年4月鹽度及徑流量差異較大)(圖9),因而2010年5月與2011年4月群落結構差異較大很可能是由于干旱引起的。有研究發現年際間由于氣候條件的變化,魚類群落結構會發生明顯的變化[69-70]。為了更好地研究這些氣象極端事件對長江口區魚類群落時間變動的影響,有必要進一步開展時間跨度更大的調查。
4.5 長江口魚類群落保護的建議
在捕獲的56種魚類中,有經濟價值的魚類有30種,這說明長江河口仍是很多經濟魚類重要的索餌場和育幼場。其中的棘頭梅童魚、刀鱭、龍頭魚、鳳鱭等重要經濟魚類[18]以刀鱭經濟價值最高,一些較大體型(>100 g/尾)刀鱭價格甚至在8 000元/kg以上(2012年價格,與漁民交流)。雖然,目前漁政部門對刀鱭捕撈船只數量進行了嚴格控制,但刀鱭資源卻一直處于衰退中[71],本研究發現11月為長江口刀鱭當年淡水孵化幼魚下海索餌的主要時間,為更好地保護刀鱭,有必要在11月份單獨設立張網作業禁漁期,以最大限度地維持其補充群體的數量。此外,長江口各種經濟魚類產量以鳳鱭最高[12],11月份鳳鱭的數量達到一年中的最大值,這種保護方法同時還能較好地保護好鳳鱭群體。此外除休漁期外,一年中長江口各個時期均允許張網作業,其中每年1—4月的鰻苗張網作業對漁業危害極大。據報道2005年前后每年超過3 000條的作業船只在長江口捕撈鰻苗,由于鰻苗體型過小,使得漁民一般選擇網目僅為1 mm左右的網具作業,過小的網目和過高的捕撈強度致使大量幼魚死亡[72]。據漁民反映,目前在長江口捕撈的張網船只有越來越多的趨勢,為了保護好長江口魚類群落,必須嚴格控制鰻苗捕撈努力量。
[1]Shenker J M,Dean J M.The utilization of an intertidal salt marsh creek by larval and juvenile fishes:abundance,diversity and temporal variation[J].Estuaries,1997,2:154-163.
[2]Rakocinski C F,Lyczkowski-Shultz J,Richardson S L.Ichthyoplankton assemblage structure in Mississippi sound as revealed by canonical correspondence analysis[J].Estuarine,Coastal and Shelf Science,1996,43:237-257.
[3]Elliott M,Hemingway K L.Fishes in Estuaries[M].Oxford:Black well Science Ltd,2002.
[4]Mc Lusky D S,Elliott M.The Estuarine Ecosystem:Ecology,Threats and Management[M].Oxford:Oxford University Press,2004.
[5]Martino E J,Able K W.Fish assemblages across the marine to low salinity transition zone of a temperate estuary[J].Estuarine,Coastal and Shelf Science,2003,56:969-987.
[6]Chícharo M A,Chícharo L,Morais P.Inter-annual differences of icht hyofauna structure of the Guadiana estuary and adjacent coastal area(SE Portugal/SW Spain):Before and after Alqueva dam construction[J].Estuarine,Coastal and Shelf Science,2006,70:39-51.
[7]Elliott M,Whitfield A K,Potter I C,et al.The guild approach to categorizing estuarine fish assemblages:a global review[J].Fish and Fisheries,2007(8):241-268.
[8]張國祥,張雪生.長江口定置張網漁業調查[J].水產學報,1985,9(2):186-198.
[9]楊偉祥,羅秉征,盧繼武,等.長江口魚類資源調查與研究[J].海洋科學集刊,1992,33:281-300.
[10]金斌松.長江口鹽沼潮溝魚類多樣性時空分布格局[D].上海:復旦大學,2010.
[11]沈新強,史赟榮,晁敏,等.夏、秋季長江口魚類群落結構[J].水產學報,2011,35(5):700-710.
[12]長江漁業資源管理委員會.長江漁業資源管理回顧與展望[M].1997.
[13]水位管理系統.http://yu-zhu.vicp.net/yzhshwgl.aspx.
[14]戈峰.現代生態學[M].北京:科學出版社,2002.
[15]金顯仕,單秀娟,郭學武,等.長江口及其鄰近海域漁業生物的群落結構特征[J].生態學報,2009,29(9):4761-4772.
[16]Nelson J S.Fishes of the World[M].4th ed.New Yor k:John Wiley &Sons,2006.
[17]Froese R,Pauly D.Fishbase.World Wide Web electronic publication[R/OL].www.fishbase.org,version(07/2010).2010.
[18]莊平,王幼槐,李圣法,等.長江口魚類[M].上海:上海科學技術出版社,2006.
[19]楊東萊,吳光宗,孫繼仁.長江口及其鄰近海區的浮性魚卵和仔稚魚的生態研究[J].海洋與湖沼,1990,21(4):346-355.
[20]劉淑德,線薇微,劉棟.春季長江口及其鄰近海域魚類浮游生物群落特征[J].應用生態學報,2008 19(10):2284-2292.
[21]張衡,何文珊,童春富,等.崇西濕地冬季潮灘魚類種類組成及多樣性分析[J].長江流域資源與環境,2007,16(3):308-313.
[22]張衡,何文珊,童春富,等.長江口低鹽淡水區潮間帶魚類群落結構季節及半月相變化[J].應用生態學報,2008,19(5):1110-1116.
[23]鄭元甲,陳雪忠,程家驊,等.東海大陸架剩余資源與環境[M].上海:上海科學技術出版社,2003.
[24]黃良敏,張雅芝,姚舒栓.廈門東海域定置網漁獲魚類種類組成及其季節變化[J].臺灣海峽,2006,25(4):509-520.
[25]Clarke K R,War wick R M.Change in marine communities:An Approach to statistical analysis and interpretation[M].2nd ed.Ply mouth:PRI MPER-E,2001.
[26]唐啟義.DPS數據處理系統[M].第2版.北京:科學出版社,2009.
[27]郁堯山,張慶生,陳衛民,等.浙江北部島礁周圍海域魚類優勢種及其種間關系的初步研究[J].水產學報,1986,10(2):145-149.
[28]Potter I C,Bird D J,Claridge P N,et al.Fish fauna of the Severn Estuary.Are there long-term changes in abundance and species composition and are the recruit ment patterns of the main marine species correlated?[J].Journal of Experi mental Marine Biology and Ecology,2001,258:15-37.
[29]沈盎綠,徐兆禮.甌江口海域夏秋季魚類初步調查[J].海洋漁業,2008,30(3):285-290.
[30]詹海剛.珠江口及鄰近水域魚類群落結構研究[J].海洋學報,1998,20:91-97.
[31]Selleslagh J,Amara R.Environ mental factors structuring fish composition and assemblages in a small macrotidal estuary(eastern English Channel)[J].Estuarine,Coastal and Shelf Science,2008,79:507-517.
[32]Thiel R,Potter I C.The ichthyofaunal composition of the Elbe estuary:an analysis in space and time[J].Marine Biology,2001,138:603-616.
[33]Akin S,Buhan E,Winemiller K O,et al.Fish assemblage structure of Koycegiz lagoon-estuary,Turkey:spatial and temporal distribution patterns in relation to environmental variation[J].Estuarine,Coastal and Shelf Science,2005,64:671-684.
[34]Jaureguizar A J,Menni R C,Guerrero R,et al.Environmental factors structuring fish communities of the Río de la Plata estuary[J].Fisheries Research,2004,66:195-211.
[35]Leit?o R,Martino F,Cabral H N,et al.The fish assemblage of the Mondego Estuary:composition,structure and trends over the past two decades[J].Hydrobiologia,2007,587:269-279.
[36]Si mier M,Laurent C,Ecoutin J,et al.The Gambia River estuary:A reference point for estuarine fish assemblages studies in West Africa[J].Estuarine,Coastal and Shelf Science,2006,69:615-628.
[37]Simier M,Blanc L,Aliau me C,et al.Spatial and temporal structure of fish assemblages in an“inverse estuary”,the Sine Saloum system[J].Estuarine,Coastal and Shelf Science,2004,59:69-86.
[38]李永振,陳國寶,孫典榮.珠江口魚類組成分析[J].水產學報,2000,24(4):312-317.
[39]Potter I C,Claridge P N,Hyndes G A,et al.Seasonal,annual and regional variations in ichthyofaunal composition in the Inner Sever n Estuary and Inner Bristol Channel[J].Journal of the Marine Biological Association,1997,77:507-525.
[40]Potter I C,Hyndes G A.Characteristics of the ichthyofaunas of southwestern Australian estuaries,including comparisons with Holarctic estuaries and estuaries elsewhere in temperate Australia:a review[J].Australian Jour nal of Ecology,1999,24:395-421.
[41]Nicolas D,Lobry J,Le P O,et al.Functional diversity in European estuaries:Relating the composition of fish assemblages to the abiotic environment[J].Estuarine,Coastal and Shelf Science,2010,88:329-338.
[42]Elliott M,Taylor C J L.The str ucture and f unctioning of an estuarine/marine fish community in the Fort h estuary,Scotland[C]//Klekowski R Z,Styczynska E,Falkowski L.Proceedings of the 21st European Marine Biology Sy mposium.Gdansk:Polish Academy of Sciences,1989:227-240.
[43]Costa M J,Elliott M.Fish usage and feeding in two industrialised estuaries-the Tagus,Portugal,and the Forth,Scotland[C]//Elliott M,Ducrotoy J P.Estuaries and Coasts:Spatial and Temporal Inter Comparisons.Fredensborg:Olsen and Olsen,1991:289-297.
[44]Mc Lusky D S,Bryant D M,Elliott M.The impact of land-clai m on the invertebrates,fish and birds of the Forth estuary[J].Aquatic Conservation:Marine and Freshwater Ecosystems.1992,2:211-222.
[45]Jin B S,Fu C Z,Zhong J S,et al.Fish utilization of a salt marsh intertidal creek in the Yangtze River estuary,China[J].Estuarine,Coastal and Shelf Science,2007,73:844-852.
[46]史赟榮,晁敏,全為民,等.2010春季長江口魚類群落空間分布特征[J].中國水產科學,2011,18(5):1141-1151.
[47]沈煥庭,潘安定.長江河口最大渾濁帶[M].北京:海洋出版社,2001.
[48]Whitfield A K.Ichthyofaunal assemblages in estuaries:a South African case study[J].Reviews in Fish Biology and Fisheries,1999,9:151-186.
[49]Beckm W,Heck K L,Able K W.A better understanding of the habitats that serve as nurseries for marine species and the factors that create site-specific variability in nursery quality will improve conservation and management of these areas[J].Bio Science,2001,51:633-641.
[50]Maes J,Stevens M,Ollevier F.The composition and community structure of the ichthyofauna of the upper Scheldt estuary:synthesis of a 10-year data collection(1991—2001)[J].Journal of Applied Ichthyology,2005,21:86-93.
[51]Whitfield A K.Biology and ecology of fishes in souther n African estuaries[M].2nd ed.Icht hyological Monogro:Smith Institute of Ichthyology.version,1998.
[52]單樂州,邵鑫斌,閆茂倉.棘頭梅童魚幼魚生物學特性的初步觀察[J].水產養殖,2007,28(6):4-6.
[53]Hagan S M,Able K W.Seasonal changes of the pelagic fish assemblage in a southern New Jersey estuary[J].Estuarine,Coastal and Shelf Science,2003,56:15-29.
[54]陳國寶,李永振,陳新軍.南海主要珊瑚礁水域的魚類物種多樣性研究[J].生物多樣性,2007,15(4):373-381.
[55]李美玲,黃碩琳 .關于長江口漁業資源管理的探討[J].安徽農業科學,2009,37(13):6196-6198.
[56]張雅芝,黃良敏.廈門東海域魚類的群落結構及種類多樣性研究[J].熱帶海洋學報.28(2):66-76.
[57]Marshall S,Elliott M.Environ mental Influences on the Fish Assemblage of the Hu mber Estuary,U.K[J].Estuarine,Coastal and Shelf Science,1998,46:175-184.
[58]金顯仕,鄧景耀.萊州灣漁業資源群落結構和生物多樣性的變化[J].生物多樣性,2000,1(8):65-72.
[59]Akin S,Winemiller K O,Gel wick F P.Seasonal and spatial variations in fish and macrocrustacean assemblage structure in Mad Island Marsh estuary,Texas[J].Estuarine,Coastal and Shelf Science,2003,57:269-282.
[60]Maci S,Basset A.Composition,structural characteristics and temporal patterns of fish assemblages in non-tidal Mediterranean lagoons:A casestudy[J].Estuarine,Coastal and Shelf Science,2009,83:602-612.
[61]M'atic-Skoko S,Peharda M,Pallaoro A,et al.Species composition,seasonal fluctuations,and residency of inshore FIsh assemblages in the Pantan estuary of the eastern middle Adriatic[J].Acta Adriatica,2005,46:201-212.
[62]M'ati-Skoko S,Peharda M,Pallaoro A,et al.Infralittoral fish assemblages in the Zr manja estuary,Adriatic Sea[J].Acta Adriatica,2007,48:45-55.
[63]Koutrakis E T,Tsikliras A C,Sinis A I.Temporal variability of the ichthyofauna in a northern Aegean coastal lagoon(Greece).Influence of environ mental factors[J].Hydrobiologia,2005,543:245-257.
[64]朱鑫華,繆鋒,劉棟,等.黃河口及鄰近海域魚類群落時空格局與優勢種特征研究[J].海洋科學集刊,2001,43:142-151.
[65]孫儒泳.動物生態學原理[M].第2版.北京:北京師范大學,1992.
[66]趙保仁.長江沖淡水鋒面變動及其與徑流量的關系[J].海洋科學集刊,1992,33:27-36.
[67]Laffaille P,Feunteun E,Lefeuvre J C,et al.Composition of fish communities in a European macrotidal salt marsh(the Moint Saint-Michel bay,France)[J].Estuarine,Coastal and Shelf Science,2000,51:429-438.
[68]于海成.長江口及鄰近海域魚類群落結構分析[D].青島:中國科學院海洋研究所,2008.
[69]Garcia A M,Vieira J P,Winemiller K O.Effects of 1997-1998 El Niňo on the dynamics of the shallow-water fish assemblage of the Patos Lagoon Estuary(Brazil)[J].Estuarine,Coastal and Shelf Science,2003,57:489-500.
[70]Martinho F,Leit?o R,Viegas I,et al.The influence of an extreme drought event in the fish community of a southern Europe temperate estuary[J].Estuarine,Coastal and Shelf Science,2007,75:537-546.
[71]黎雨軒,何文平,劉家壽,等.長江口刀鱭耳石年輪確證和年齡與生長研究[J].水生生物學報,2010,34(4):787-793.
[72]Zhang H,He W S,Tong C F,et al.The effect of fishing the anguillid elver(Anguilla japonica)on the fishery of the Yangtze Estuary[J].Estuarine,Coastal and Shelf Science,2008,76:902-908.
