生理活性物質對遭遇低溫逆境煙草抗性生理指標的影響
2014-09-28 03:23:40謝會雅史端甫張紅兵羅真華段伶俐周真珍楊莉穎賀利雄
湖南農業科學 2014年23期
謝會雅,史端甫,張紅兵,羅真華,段伶俐,周真珍,楊莉穎,賀利雄
(1. 湖南省煙草公司 株洲市分公司,湖南 株洲 412000;2. 湖南農業大學 資源環境學院,湖南 長沙 410128;3. 湖南農業大學 生物科學技術學院 湖南 長沙 410128)
煙草是原產于亞熱帶的喜溫作物,其生長最適溫度為25~28℃,10~13℃停止生長、1~2℃低溫可使幼苗死亡,環境溫度尤其是低溫對煙草的生育、產量和品質都有明顯影響。在湘南、湘西北和湘中煙區低溫危害(即倒春寒)頻發,導致早春移栽后煙苗受害、大田前期生長緩慢以及導致早花,嚴重引起煙葉產量和品質的波動,也一直制約優質煙葉生產的發展[1],目前增強品種的低溫耐性已成為煙草品種改良的主要目標之一。針對困擾煙草生產的“倒春寒”危害這一實際問題,通過遺傳改良育成耐低溫新品種是解決“倒春寒”危害的根本性策略,但這耗時周期長,且需要大量的人力、物力和財力。因此,通過農藝措施如噴施煙草低溫耐性調節劑提高煙草幼株的低溫耐性,不失為及時應對早春低溫危害的一種方法。
研究報道表明,多種外源化學物質(生理活性物質)如甜菜堿[2-3]、聚乙二醇[4]、水楊酸[4-5]、5-氨基乙酰丙酸[6-7]和ABA、氯化鈣、維生素C[8]等對植物的逆境抗性具有調節作用。本研究在低溫逆境脅迫條件下,對葉面噴施水楊酸、甜菜堿、聚乙二醇和5-氨基乙酰丙酸煙草植株的抗寒性相關生理指標進行了檢測分析,評價其改善煙草低溫抗性的生理效果,以期為煙草苗期低溫耐性調節劑有效組分的篩選提供初步的理論依據。
1 材料與方法
1.1 田間試驗材料
參試煙草品種為K326、由中國煙草中南試驗站提供,采用煙草育苗專用基質和漂浮育苗法,依照煙草育苗技術進行肥水管理;出苗后1~2 片真葉期間苗、3~4片真葉期移植盆栽,幼苗至6~7 片真葉時(苗齡約55 d 左右),選用生長旺盛、長勢基本一致的幼苗供試驗用。
1.2 室內實驗設計及處理方法
選用與植物低溫逆境抗性關系密切的水楊酸(SA)、甜菜堿(GB)、聚乙二醇(PEG 6000)和5-氨基乙酰丙酸(5-ALA)4種生理活性物質(以清水為對照),葉面噴施濃度分別為(輔以0.1%Tween 20):10 mg/L、15 mmol/L、2.5 mmol/L 和10%。
將供試煙苗移入人工氣候室常溫馴化3 d 后(22~25℃,5 000 lx、光照12 h),整株噴施生理活性物質溶液,以葉片上下表面濕透為度、噴施處理15 株;噴施處理24 h 后,開始低溫逆境處理。
于2013~2014年用人工氣候室(AGC-D003N,浙江求是人工環境有限公司)模擬“倒春寒”時期氣候條件實施低溫逆境脅迫處理(6~7℃,1 000 lx、光照12 h),期間正常水分管理。低溫逆境處理第0、1、3、5、7 d,光照6 h后,選取中位成熟葉片進行葉片相對電導率測定,重復5次,并採集各處理鮮葉樣品液氮冷凍、研碎,-80℃超低溫冰箱保存,供后續理化分析用;用Excel 2010 處理實驗數據。
1.3 葉片生理指標檢測方法
生理指標定量分析檢測依照《生物化學實驗教程》[9]和《植物生理學實驗技術》方法[10]、或略加改良后進行(重復3 次),比色法使用SP-752 紫外分光光度計(上海光譜)。檢測方法:(1)相對電導率測定使用DDS-307 電導儀(上海雷磁)測定葉片組織外滲液的相對電導率;(2)葉綠素含量測定采用80%丙酮提取比色法;(3)可溶性糖含量測定采用蒽酮比色法;(4)游離脯氨酸含量測定采用酸性茚三酮比色法;(5)過氧化氫酶活性測定采用高錳酸鉀滴定法;(6)丙二醛含量測定采用硫代巴比妥酸(TBA)比色法。
2 結果與分析
2.1 生理活性物質處理對煙草葉片細胞膜相對電導率的影響
當植物遭遇逆境脅迫時,半透性的細胞膜既是感受逆境脅迫的最敏感部位,也是細胞首先被“傷害”的部位,由于細胞膜的結構“受損”導致其透性增大、細胞內可溶性物質和電解質會被動地向膜外滲漏,細胞浸出液電導率的增大反映了質膜的傷害程度[11]。
從圖1 可見,與低溫處理前(0 d)相比,低溫處理第1、3、5、7 d、對照的相對電導率分別增加44.82%、75.30%、102.70%和117.72%,而4種生理活性物質處理的電導率增長幅度均低于對照,分別為11.06%~23.25%、31.51%~47.91%、50.60%~59.45%和60.43%~73.03%。說明4種生理活性物質能減輕低溫脅迫下煙草葉片受到的傷害、減少細胞溶質的外滲,對煙草細胞膜具有良好的保護作用,且4種生理活性物質中,5-ALA、SA 及GB 的效果優于PEG。
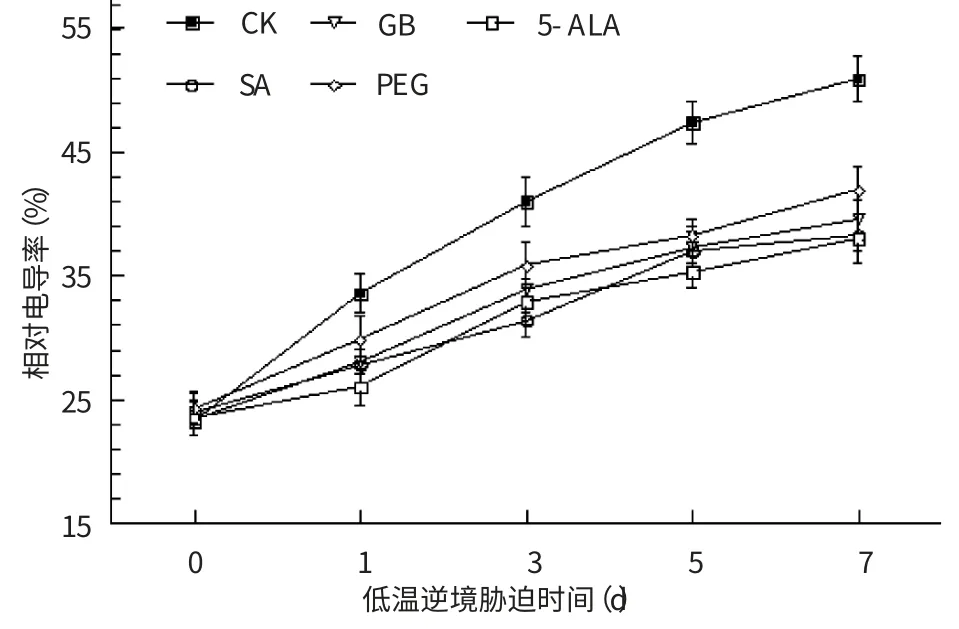
圖1 生理活性物質處理煙草葉片細胞膜相對電導率的影響
2.2 生理活性物質處理對煙草植株可溶性糖、游離脯氨酸含量的影響
當遭遇逆境脅迫時,植物會主動調整自身代謝、強化專一的生理代謝,積累可溶性糖、脯氨酸和甜菜堿等相溶性溶質來提高細胞液濃度、降低細胞水勢和冰點,保護細胞膜結構的完整性[12];防止蛋白質發生凝固變形,在細胞蛋白質(酶)高級結構的維持、翻譯后的修飾和輸運過程中發揮“小分子伴侶”的生理作用;此外,脯氨酸還直接參與逆境脅迫中O2-、OH-等活性氧清除過程[13]。因此,可溶性糖、脯氨酸等相溶性溶質的含量是植物逆境抗性的重要生理指標。
可溶性糖和游離脯氨酸含量的測定結果如圖2 所示,低溫脅迫引起了煙草體內可溶性糖和脯氨酸的積累。低溫處理第1、3、5、7 d,對照的可溶性糖含量分別較低溫處理前增加了8.30%、33.60%、83.40%和98.81%,SA、GB、PEG 和5-ALA 處理的可溶性糖含量的增長均高于對照,分別為17.98%~33.84%、53.18%~73.56%、96.25%~119.39%、117.98%~134.98%,4種生理活性物質的處理效果以5-ALA 略優、但差異不顯著(圖2a)。隨著低溫脅迫時間的延長,脯氨酸含量也表現出了與可溶性糖含量類似的變化,如低溫脅迫第7 天,對照脯氨酸含量較低溫脅迫前增加了84.51%,4種生理活性物質處理的煙苗脯氨酸含量增加幅度為93.90%~115.55%,顯著高于對照(圖2b)。
綜合來看,4種生理活性物質處理均較對照進一步促進了低溫逆境脅迫處理期間可溶性糖和脯氨酸的積累,對煙苗低溫脅迫的響應能力具有明顯的調節作用,可增強煙草植株的低溫抗性。
2.3 生理活性物質處理對煙草葉片過氧化氫酶活性和丙二醛含量的影響
植物遭遇逆境脅迫時,富集的活性氧(ROS)直接氧化損傷細胞內生物大分子,引起一系列生理生化紊亂;同時,加劇膜脂質的氧化程度、破壞生物膜的完整性,增加膜脂質氧化產物丙二醛(MDA)的量。另一方面,植物體內過氧化氫酶(CAT)廣泛存在,是植物組織細胞清除H2O2的主要保護酶,逆境脅迫下維持有較高的CAT 活性,可減輕活性氧的毒害作用、穩定膜結構,提高植物的抗逆性[14]。
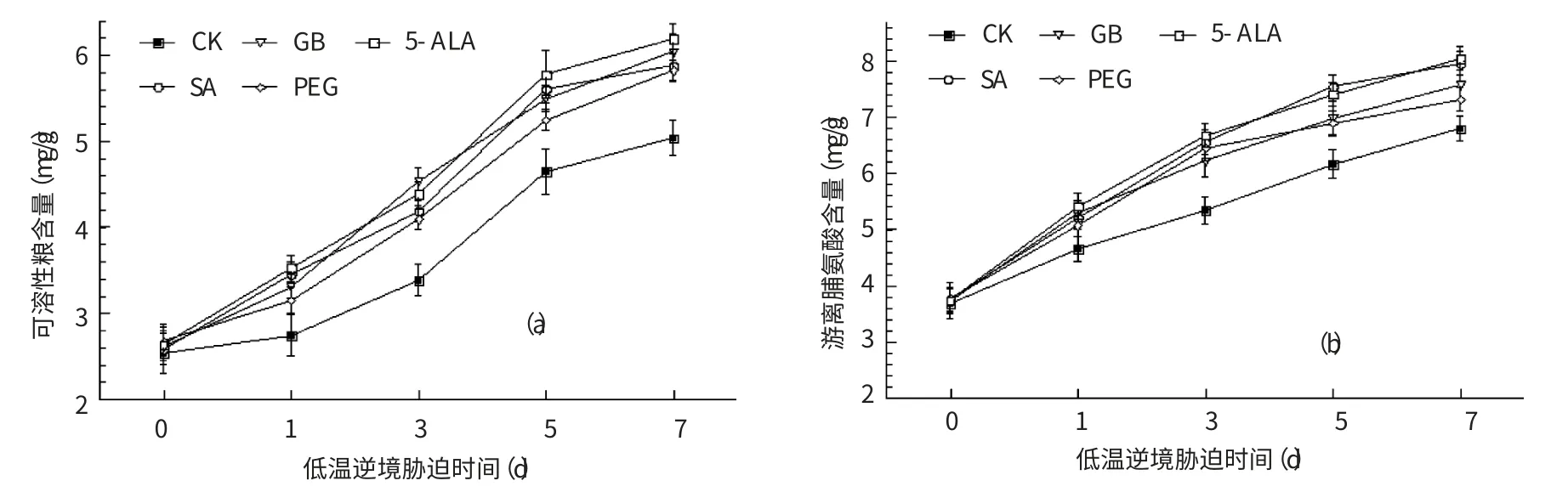
圖2 生理活性物質處理對煙草可溶性糖(a)、游離脯氨酸(b)含量的影響
從圖3a 可看出,對照及各處理的煙苗CAT 活性隨低溫脅迫的延長持續下降,低溫處理第1、3、5、7 d,對照較低溫處理前分別降低18.16%、46.78%、70.17%和76.23%,4個生理活性物質處理的降低幅度分別為7.54% ~9.57% 、13.48% ~19.73% 、22.69% ~35.41% 、33.21%~45.61%,均低于對照組,顯示出4種生理活性物質對CAT 具有較好的保護作用、CAT 維持有較高的活性。
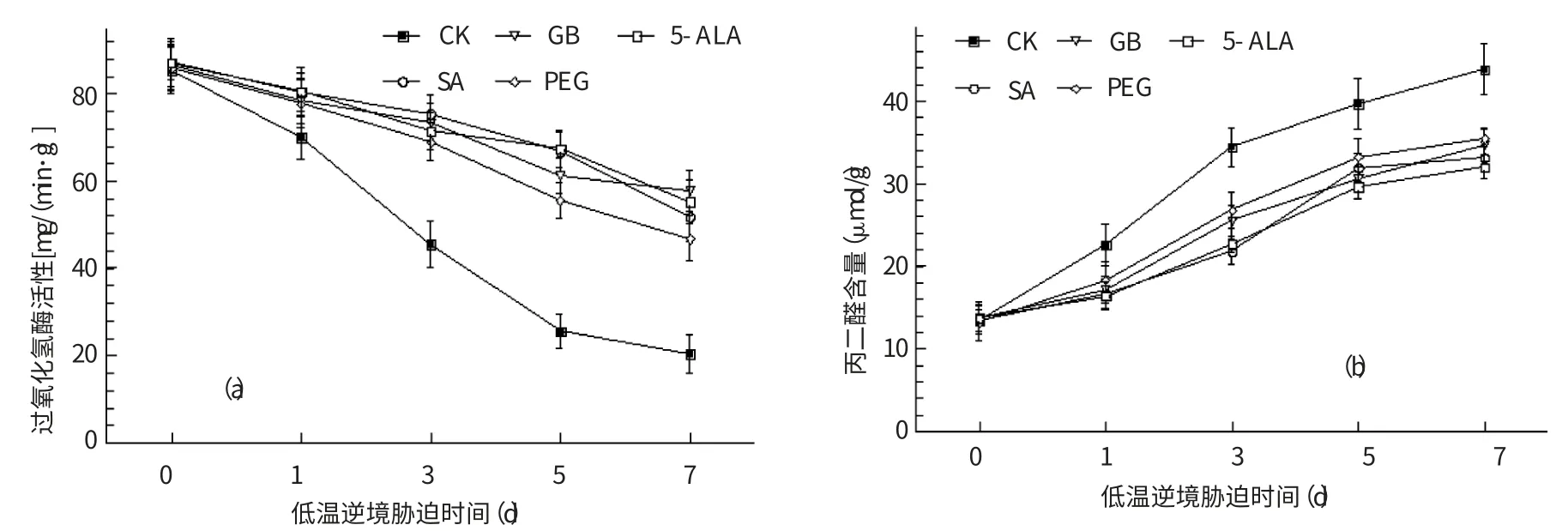
圖3 生理活性物質處理對煙草過氧化氫酶活性(a)、丙二醛含量(b)的影響
低溫逆境脅迫可導致煙苗MDA 的產生,并隨低溫脅迫時間的增加而上升(圖3b),如低溫處理第3 天,對照較處理前增加了156.8%,而4種生理活性物質處理分別增加了62.21%、85.43%、103.42%、65.64%,明顯低于對照,4種生理活性物質處理可減輕低溫逆境脅迫下煙苗MDA 的增加,其中SA、GB 和5-ALA 的處理效果優于PEG 處理。
從CAT 活性和MDA 含量變化的結果來看,生理活性物質處理可減輕低溫脅迫下煙草CAT 活性的下降,減緩生物膜脂質氧化損害、對細胞質膜系統具有良好的保護作用;另外,SA、GB 和5-ALA 這3種生理活性物質對煙苗低溫耐性指標改善的生理效果比PEG 處理更為明顯。
2.4 生理活性物質處理對煙草葉片葉綠素含量的影響
低溫逆境脅迫將導致植物光合系統的“低溫光抑制”,葉綠體色素被氧化“漂白”分解是光合系統的低溫傷害之一[15]。此外,煙草生產中“倒春寒”發生時期的光照較弱,而在弱光照下植物的光合速率與葉綠素含量存在良好的線性關系,葉綠素含量直接影響著植物光合作用的水平[16]。
表1 所示結果表明,低溫脅迫使煙草葉片葉綠素含量逐漸減少。低溫處理第1、3、5、7 d,對照葉綠素含量分別較低溫處理前減少了11.02%、22.83%、33.86%和39.37%,4種生理活性物質處理的減少幅度分別為3.97% ~6.35% 、8.73% ~13.60% 、16.67% ~23.26% 、21.43%~28.00%,均顯著低于對照,表明這4種生理活性物質處理減緩了低溫脅迫所導致的葉綠素分解速度,對葉綠素有一定的保護作用。低溫脅迫下煙草的葉綠素保有量較高,可促進煙苗光合能力的維持,為保持煙苗的生長和營養吸收、增強煙苗的抗逆性提供基本保障。
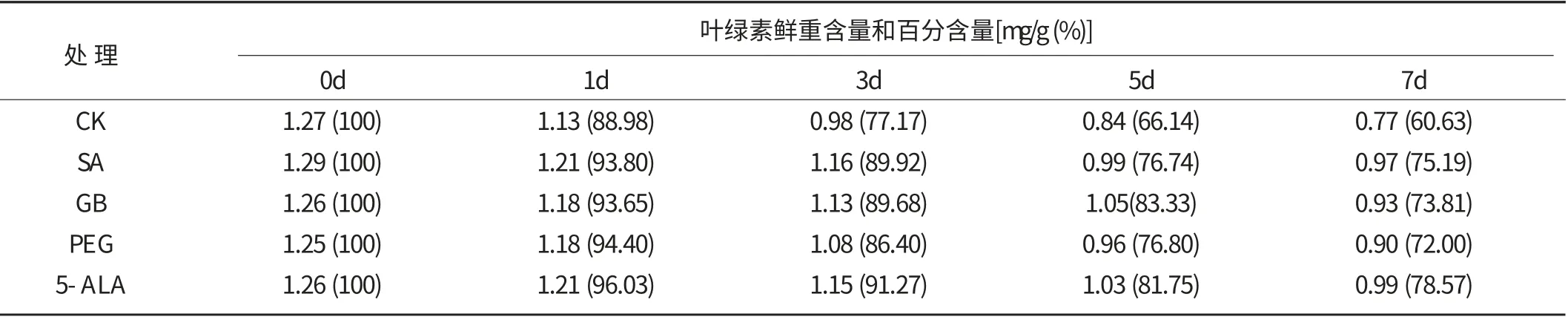
表1 低溫脅迫下生理活性物質處理煙草葉片葉綠素含量的變化
3 小結與討論
本研究表明,噴施適宜濃度的水楊酸(SA)、甜菜堿(GB)、聚乙二醇(PEG)和5-氨基乙酰丙酸(5-ALA),能促進低溫脅迫下煙草可溶性糖和游離脯氨酸的積累、減輕過氧化氫酶活性和葉綠素含量的下降、降低煙草葉片的相對電導率和MDA 含量,從而提高了煙草植株的抗寒能力。另外,綜合比較這4種生理活性物質的處理效果,初步確定SA、5-ALA 和GB 優于PEG,為煙草苗期低溫耐性調節劑有效組分篩選提供了參考。
為了確保研究結果的準確性和實用性,后續研究中:①本實驗使用的生理活性物質對植物抗寒性的其他機制如煙苗的根系活力、光合活性、水勢、抗氧化系統等因素的影響有待進一步驗證;②植物激素或植物生長調節物質對煙草生長發育、煙葉產量和品質等方面有較大的影響,本研究使用的生理活性物質在此方面、尤其是對煙草品質方面的影響如何有待檢測分析。
[1]岳彩鵬,韓錦峰,陳 衛.煙草開花研究進展[J].煙草科技,2001,170(9):36-40.
[2]韓冬芳,李雪萍,李 軍,等.甜菜堿提高植物抗寒性的機理及其應用[J].熱帶亞熱帶植物學報,2010,18(2):211-216.
[3]楊 雷,畢相東,王關林.3種抗寒保護劑對大葉黃楊葉片抗凍性的影響[J].遼寧師范大學學報(自然科學版),2005,28(3):342-34
[4]劉自剛,張 雁.水楊酸在農業生產中的利用[J].作物雜志,2007,(2):16-19.
[5]康國章,歐志英,王正詢,等.水楊酸誘導提高香蕉幼苗耐寒性的機制研究[J].園藝學報,2003,30(2):141-146.
[6]汪良駒,姜衛兵,章 鎮,等.5-氨基乙酰丙酸的生物合成和生理活性及其在農業中的潛在應用[J].植物生理學通訊,2003,39(3):185-192.
[7]宋士清,郭世榮.5-氨基乙酰丙酸的生理作用及其在農業生產中的應用(綜述)[J].河北科技師范學院學報,2004,18(2):54-57,72.
[8]李艷軍.外源化學物質誘導對番茄苗期抗冷性的影響[D].黑龍江:東北農業大學園藝學院,2005.
[9]王金亭,方俊.生物化學實驗教程[M].武漢:華中科技大學出版社,2010.
[10]蕭浪濤,王三根.植物生理學實驗技術[M].北京:中國農業出版社,2005.
[11]Warren G J,Thorlby G J,Knight M R.Themolecu larbiologicala pproach tounder standing freezing to lerance in the model plant,Arabi dopsis thaliana[J].Env Stress or sand Gene Re,2000,(1):245-258.
[12]陳新建,陳占寬,劉國順,等.植物對水分脅迫響應的分子機制與抗逆基因工程的研究進展[J].熱帶亞熱帶植物學報,2000,8(1):81-90.[13]謝 虹,楊 蘭,李忠光.脯氨酸在植物非生物脅迫耐性形成中的作用[J].生物技術通報,2011,(2):23-26,60.
[14]晉 艷,楊宇虹,華水金,等.低溫脅迫對煙草保護性酶類及氮和碳化合物的影響[J].西南師范大學學報,2007,32(3):74-79.
[15]段 偉,李新國,孟慶偉,等.低溫下的植物光抑制機理[J].西北植物學報,2003,23(6):1017-1023.
[16]許大全.葉綠素含量的測定及其應用中的幾個問題[J].植物生理學通訊,2009,45(9):896-898.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
聲屏世界(2014年6期)2014-02-28 15:18:09
