甜菜雄性不育系及保持系花期蛋白差異分析
2014-09-23 02:58:30程大友代翠紅
哈爾濱工業大學學報 2014年10期
程大友,崔 杰,牛 佳,代翠紅,白 晨
(1.哈爾濱工業大學食品科學與工程學院,150090哈爾濱;2.內蒙古自治區農牧業科學院甜菜研究所,010031呼和浩特)
目前利用作物的雄性不育特性生產雜交種已成為異交作物種子生產的主流.研究作物雄性不育分子機理已成為分子育種的研究熱點[1].隨著蛋白質組學技術的發展,蛋白質組分析已應用于擬南芥、水稻和西紅柿[2-4]等植物的研究中.在成熟的花粉中大量的蛋白已被鑒定出來.通過對水稻花藥小孢子時期的蛋白質組分析,揭示了富含甘氨酸蛋白、熱激蛋白、翻譯控制蛋白和一些管家蛋白以及在小孢子發育過程中的潛在因子[5].在擬南芥雄性不育突變體中,孢子發育時腺嘌呤磷酸核糖轉移酶(APRT)活性的喪失被認為是導致不育的原因[6];水稻的花藥中,低溫脅迫也能引起雄性不育,這與熱激蛋白和β-拓展蛋白的部分阻斷有關[7],在大豆的雄性不育系 NJCMS1A及其保持系中鑒定出了一些在乙烯合成、細胞程序性死亡、ATP合成、淀粉合成過程中起關鍵作用的蛋白,推測細胞質雄性不育可能是因為這些過程中某蛋白缺失或功能改變,造成這些程序紊亂,進而導致雄性不育[8];在紅蓮型水稻的CMS花藥中發現缺少與乙酰輔酶A合成有關的酶(乙酰輔酶A合成酶和二氫硫辛酸脫氫酶),導致三羧酸循環(TCA)和脂肪酸代謝不能正常進行,影響呼吸和能量代謝,最終導致細胞質雄性不育[9].此外,研究發現水稻、小麥、番茄等雄性不育還與光合作用過程中的一些酶有關,如葉綠體氧增強蛋白1(chloroplast oxygen-evolving enhancer protein 1)和 1,5-二磷酸核酮糖羧化酶(Rubisco)[10-12].因此,推測雄性不育可能與光合酶系的功能缺陷有關.甜菜在育性相關蛋白方面的報道較少,Masayuki P等在甜菜雄性不育I-12CMS(3)細胞質中找到一個12 KD的蛋白,該蛋白是只在不育系中出現的orf129的蛋白產物;Atp6的5'前導序列preSatp6只在CMS型線粒體中表達一種35 KD的蛋白[13-14],推測這兩個蛋白可能與雄性不育有關,但是并沒有進行進一步的研究.
本文利用2-DE與MALDI-TOF-MS方法對甜菜質核互作型雄性不育系及保持系花蕾發育的3個階段蛋白差異表達進行研究,以期揭示Owen型甜菜不育系與保持系花蕾不同發育時期蛋白表達的差異.
1 實 驗
1.1 實驗材料
所用材料來自哈爾濱工業大學自育的甜菜不育系DY5-CMS及其保持系DY5-O.經過春化處理的甜菜母根,于4月20日種植在哈爾濱工業大學甜菜育種試驗田,6月25日~7月15日期間進行甜菜花期取樣調查.甜菜花蕾發育分成3個時期:雄蕊原基分化期、四分體時期及單核時期.
1.2 實驗方法
1.2.1 甜菜花蕾蛋白的提取
采用TCA-丙酮沉淀法提取蛋白[15],將濃縮后的蛋白干粉置于-20℃保存備用.取13 mg濃縮后的蛋白干粉,加入150 μL蛋白裂解液反復凍融使其復融,采用Brandford法測定蛋白含量,標準蛋白為牛血清白蛋白(BSA).
1.2.2 雙向凝膠電泳
第一向等電聚焦電泳參照Bio-Rad公司等電聚焦系統說明書進行;第二向電泳參照文獻[15]方法進行.
1.2.3 MALDI-TOF-TOF/MS質譜分析
經脫色清洗后,將凝膠掃描采集圖像利用Image Master 2D Platinum 5.0軟件(GE公司)分析蛋白質圖譜.經點探測、匹配,找出差異點.將3次重復均出現且特征明顯(含量差值大于2.0以上的)的差異蛋白點回收后委托廣州慧晶生物科技有限公司進行MALDI-TOF-TOF/MS質譜分析.
2 結果與分析
2.1 雄蕊原基分化期
對甜菜雄蕊原基分化期花蕾蛋白的差異進行分析,在不育系中檢測到蛋白點405個,保持系中檢測到325個(如圖1),檢測出的差異蛋白總數在不育系中升高的有14個點,在保持系中升高的有8個點.

圖1 不育系DY5-CMS及其保持系DY5-O的1期花蕾蛋白2DE圖譜
利用PMF分析獲得質譜圖(見圖2、3),去除假陽性點后,鑒定出6種蛋白(見表1),即NADH脫氫酶、二硫辛酰胺脫氫酶、可溶性無機焦磷酸酶、ATP合成酶的β亞基和ATP合成酶D鏈、葡萄糖調節蛋白同系物.
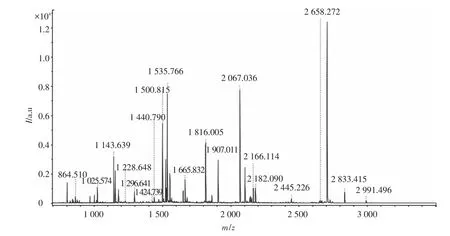
圖2 雙向電泳圖譜中蛋白點B05的肽質量指紋圖
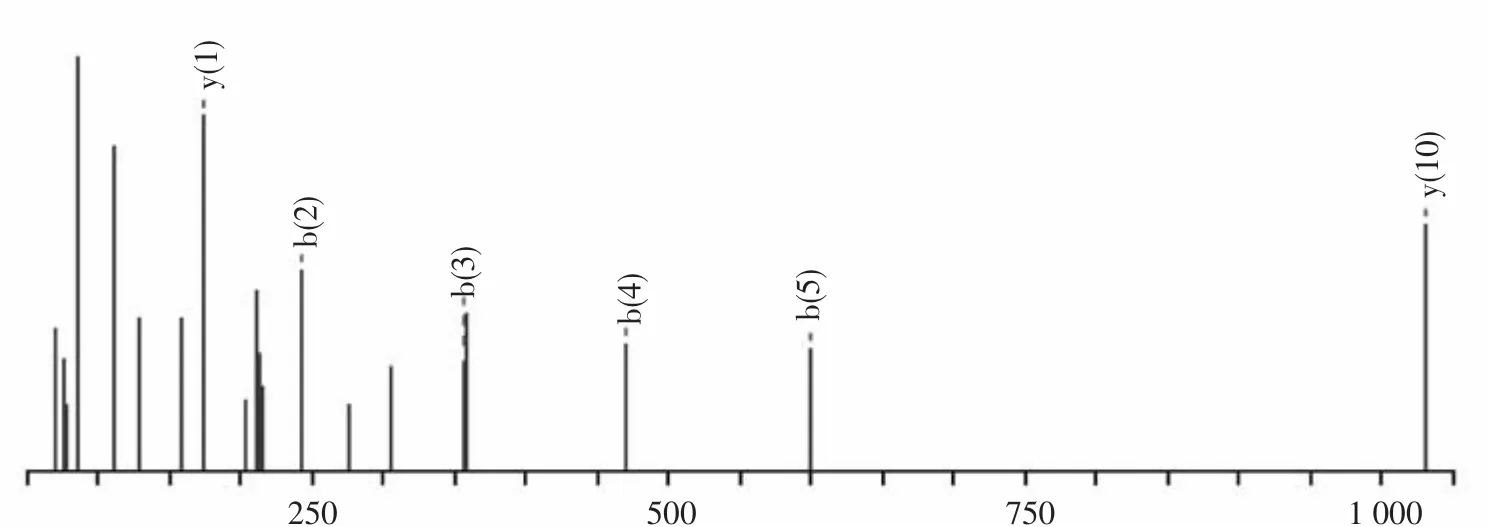
圖3 B05胰蛋白酶酶解后肽段NQⅠDEⅠVLVGGSTR的MS/MS質譜圖

表1 DY5-CMS和DY5-O 1期差異蛋白的PMF數據庫搜索鑒定結果
2.2 四分體時期
對甜菜四分體時期花蕾蛋白的差異進行分析,在不育系中檢測到蛋白點385個,保持系中檢測到306個(如圖4),檢測出2.0倍表達量以上的差異蛋白在DY5-O中有5個點表達上調,在DY5-CMS中有11個點表達上調.利用MALDITOF-MS分析去除假陽性點后,鑒定出4個蛋白.且這4個蛋白均在不育系中表達上調(見表2).
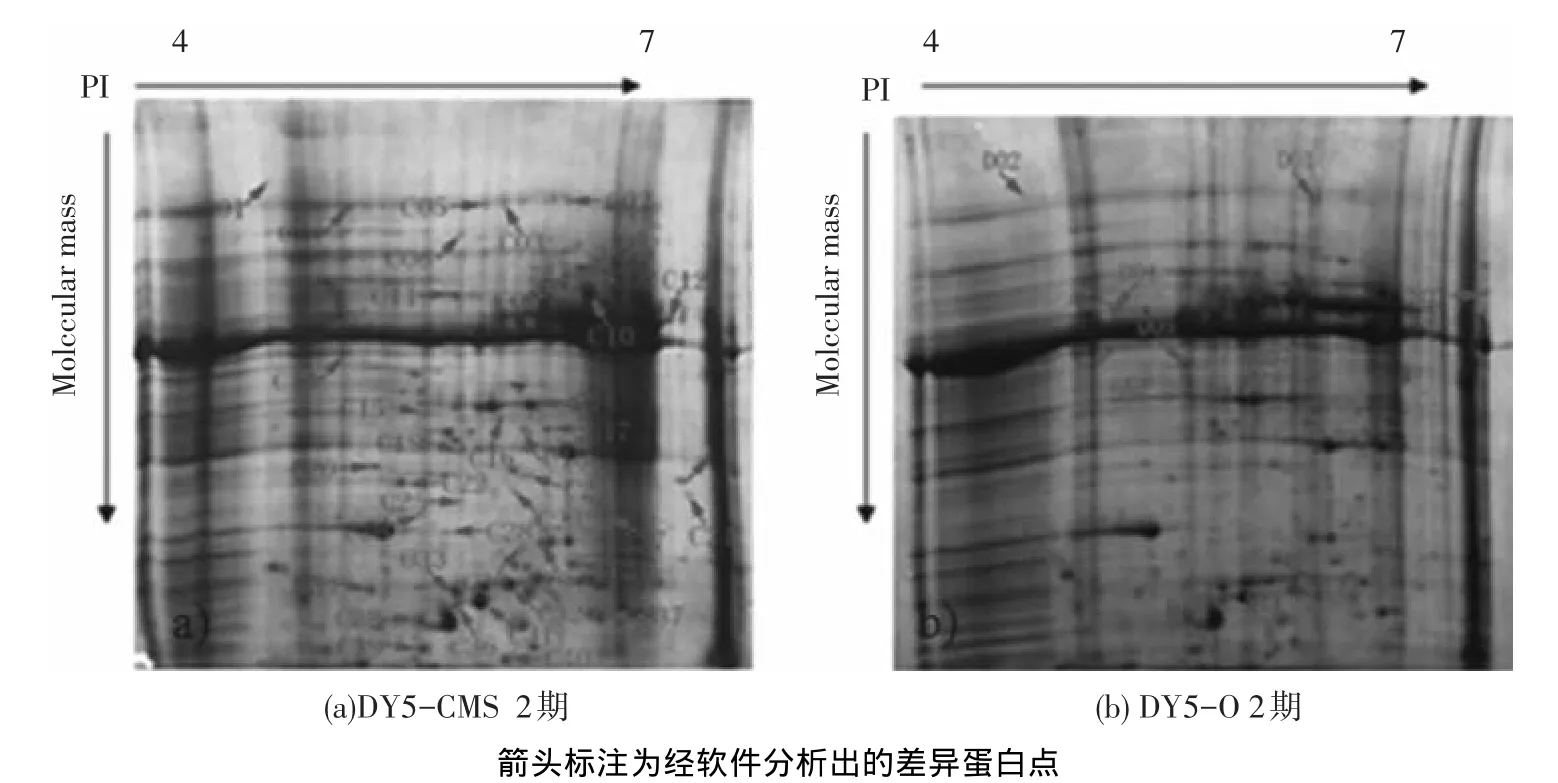
圖4 不育系DY5-CMS及其保持系DY5-O的2期花蕾蛋白2DE圖譜

表2 DY5-CMS和DY5-O 2期差異蛋白的PMF數據庫搜索鑒定結果
2.3 單核期
對甜菜單核時期花蕾蛋白的差異進行分析,在不育系中檢測到蛋白點460個,保持系中檢測到508個,檢測出2.0倍表達量以上的差異蛋白總數:從3期中找到24個差異點,其中在DY5-O中有16個點表達上調,在DY5-CMS中有8個點表達上調.利用MALDI-TOF-MS分析去除假陽性點后,鑒定出6個蛋白(見表3).
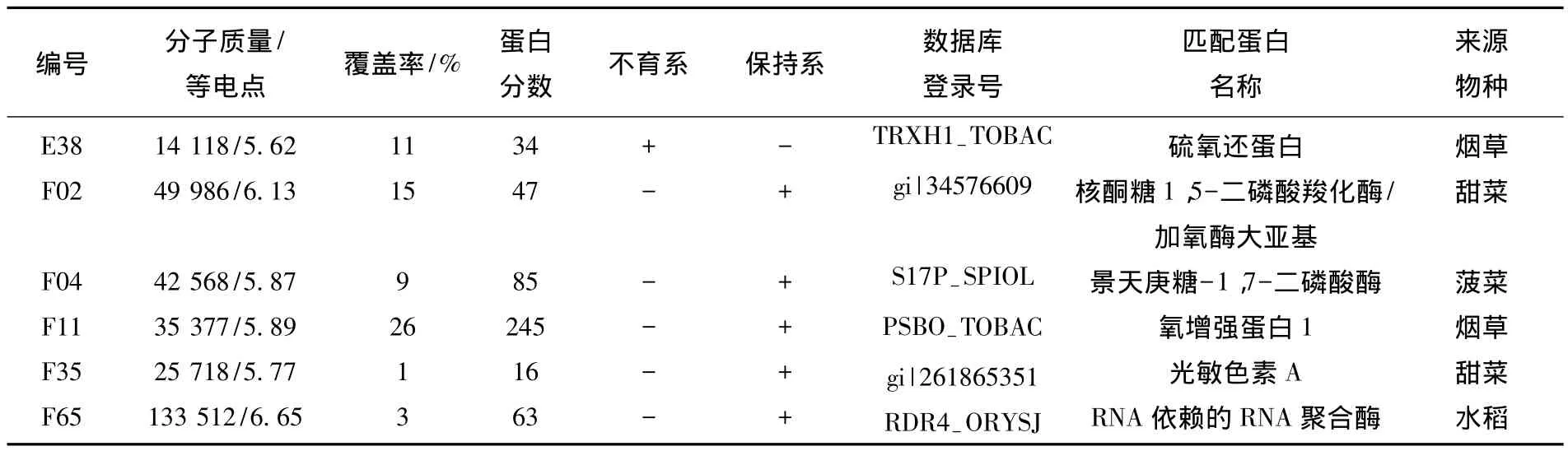
表3 DY5-CMS和DY5-O 3期差異蛋白的PMF數據庫搜索鑒定結果
3 討論
3.1 雄蕊原基分化期花蕾育性蛋白差異分析
從甜菜不育系及保持系1期花蕾蛋白中鑒定出6個差異蛋白,其中不育系中有NADH脫氫酶、二硫辛酰胺脫氫酶、可溶性無機焦磷酸酶、ATP合成酶的β亞基及ATP合成酶的D鏈5個蛋白表達上調,這5個蛋白均與呼吸和能量代謝有關.在保持系中只檢測出葡萄糖調節蛋白78表達上調.
NADH脫氫酶是線粒體呼吸鏈的重要組成部分,在糖酵解、糖異生、三羧酸循環中起著重要作用.ATP合成酶是高等植物細胞內最大的酶系之一,可以合成ATP為代謝活動提供能量.本實驗發現的5種與能量代謝有關的蛋白均在不育系中表達上調,推測ATP合成酶被水解導致能量供應不足,促使雄性不育系植物中與能量代謝有關酶的表達量上調.因此,推測這些與呼吸及能量代謝有關的蛋白與甜菜的細胞質雄性不育有關.
葡萄糖調節蛋白78(GRP78)是內質網上一種重要的分子伴侶,能阻止內質網內新生肽聚集,并輔助內質網中新生肽形成正確構象;GRP78還是一種鈣結合蛋白,維持內質網鈣穩態及內環境的穩態.本實驗只在甜菜的保持系DY5-O中發現了該蛋白,而在不育系DY5-CMS中沒有發現,推測在不育系中該酶的缺失導致花粉發育過程中的代謝紊亂,最終導致不育現象的產生.
3.2 四分體時期花蕾育性蛋白差異分析
從甜菜DY5-CMS及DY5-O的四分體時期花蕾蛋白中找到4個差異蛋白,分別為順烏頭酸酶、抗壞血酸過氧化物酶、延伸因子2及半胱氨酸合成酶.他們與甜菜的雄性不育有關.
順烏頭酸酶(aconitase,ACO)是三羧酸循環中能量轉化的一個重要控制酶.在介導植物抗病、氧化脅迫和調控細胞死亡方面具有重要作用.張明珠等[16]發現順烏頭酸酶基因在殺雄劑SQ-1誘導小麥雄性不育花藥中的表達,在生理型不育花藥發育的單核早期cACO基因表達水平與同期可育花藥相比顯著升高,這與本實驗的研究結果恰好吻合.
抗壞血酸過氧化物酶是葉綠體中清除H2O2的主要酶,在鹽漬、熱休克等逆境條件下均易導致轉錄水平和活性提高,但在本實驗中其對育性生理生化的影響有待進一步研究.
延伸因子(elongation factor)是參與蛋白翻譯延伸的重要蛋白質.研究發現,延伸因子在原核生物以及植物線粒體和質體等細胞的蛋白質合成延伸階段發揮著重要的作用,在生長信號傳導、抗旱、抗病等方面也有重要作用[17].
3.3 單核期花蕾育性蛋白差異表達分析
在甜菜的3期花蕾中鑒定出的7個蛋白中,有5個是與植物光合作用有關的蛋白,分別是H型硫氧還蛋白 1、核酮糖 1,5-二磷酸羧化酶(Rubisco)大亞基、景天庚糖-1,7-二磷酸酶、氧增強蛋白1和光敏色素A,這些蛋白均在植物的光合作用中起著重要作用.另外2個分別是脂肪氧化酶(LOX)和RNA依賴的RNA聚合酶.
硫氧還蛋白可以通過還原靶蛋白中的二硫鍵參與細胞的一系列生化反應.在光合器官內,葉綠體鐵氧還蛋白/硫氧還蛋白(Fd/Trx)參與葉綠體中還原磷酸戊糖途徑酶的激活、氧化磷酸戊糖途徑酶的暗激活等酶活性的調節[18].本實驗表明硫氧還蛋白在甜菜不育系3期花蕾中表達上調,說明在甜菜不育系的花藥發育至此期該酶起到一定的作用.
核酮糖 1,5-二磷酸羧化酶/加氧酶(Rubisco)能催化光合作用的二氧化碳固定反應.該類蛋白的差異表達普遍存在于植物雄性不育系和保持系或不育系和轉換系之間,在水稻、蘿卜、小麥、甘藍、番茄、煙草等植物雄性不育中均檢測到Rubisco的功能缺陷或下調[19-20].本實驗在甜菜保持系的花蕾中也檢測到這種蛋白,而在不育系中缺失.推測Rubisco在不育系中缺失影響發育過程中糖和蛋白質含量,而糖和蛋白質代謝的失調與不育系花藥中小孢子的敗育有關.花粉敗育的同時營養代謝紊亂,合成受阻,分解加劇,影響雄性器官的發育,最終導致不育.
景天庚酮糖-1,7-二磷酸酶(SBPase)在植物光合作用卡爾文循環過程中控制著碳的流入和再生,對核酮糖-1,5-二磷酸的再生也起著重要作用.SBPase活性的改變會影響植物的生殖生長,如表現出開花延遲,開花數減少的現象[21].本實驗發現在甜菜的保持系中SBPase的表達上調,推測不育系中SBPase缺失影響了核酮糖-1,5-二磷酸的再生,進而影響花粉的敗育.
氧增強蛋白1(oxygen-evolving enhancer protein 1,OEE1)是由核基因錳簇穩定外周蛋白PsbO基因編碼的定位于葉綠體上的蛋白.在甜菜不育系的單核時期花蕾中該蛋白表達下調,其作用機理尚不明確.
光敏色素是一種色素蛋白復合體,可以調節開花,還能控制性別表達和育性的轉變.光敏核不育水稻在長日照下雄性不育,在短日照下可育.光敏色素通過調節光周期參與光敏水稻的雄性器官發育和育性轉變,本實驗在保持系中檢測到的光敏色素A在不育系中表達下調或缺失,推測光敏色素A是與雄性不育有關的蛋白之一.
RNA依賴的RNA聚合酶(RDR)通過基因沉默途徑參與植物的生長發育調節、逆境應答以及表觀遺傳修飾等生物學過程.本實驗發現RDR在不育系甜菜的3期花蕾中表達下調或缺失,推測不育系中RDR的表達下調或缺失可能對甜菜不育系花粉敗育后期有影響.
4 結 論
1)雄蕊原基分化期,找到6個與雄性不育有關的蛋白,其中5個蛋白還與能量和呼吸代謝有關,即NADH脫氫酶、二硫辛酰胺脫氫酶、可溶性無機焦磷酸酶、ATP合成酶的β亞基和ATP合成酶D鏈.這5個蛋白均在不育系中表達上調,只有一個在保持系中表達上調的蛋白是葡萄糖調節蛋白78.
2)四分體時期,分別是順烏頭酸酶、抗壞血酸過氧化物酶、延伸因子2及半胱氨酸合成酶可能與雄性不育有關.這些蛋白均在不育系中表達上調.
3)單核時期,篩選出6個可能與育性有關的蛋白.其中只有硫氧還蛋白在不育系中表達上調,其余5個蛋白均在保持系中表達上調.
[1]CHENG D Y,KITAZAKI K,XU D C,et,al.The distribution of normal and male-sterile cytoplasms in Chinese sugar-beet germplasm[J].Euphytica,2009,165:343-351.
[2]CARRETERO-PAULETL,GALSTYANA,ROIGVILLANOVA I,et al.Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis,poplar,rice,moss and algae[J].Plant Physiol,2010,153:1398-1412.
[3]DAI S,CHEN T,CHONG K,et al.Proteomics identification of differentially expressed proteins associated with pollen germination and tube growth revealscharacteristics of germinated Oryza sativa pollen[J].Mol Cell Proteomics,2007,6:207-230.
[4]XU K,CAO M J,ZHU Y G,et al.Differential expression of mitochondrial proteins between C-type cytoplasmic male sterility line C48-2 and its maintainer line in maize[J].Acta Agron Sin,2008,34(2):232-237.
[5]NIJAT I, TURSUN K, WEINMAN J J, et al.Characterization of rice anther proteins expressed at the young microspore stage[J].Proteomics,2001,1:1149-1161.
[6]WU R,DEPHOURE N,HAAS W,et al.Correct interpretation of comprehensive phosphorylation dynamics requires normalization by protein expression changes[J].Mol Cell Proteomics,2011,10:M111 009654.
[7]NIJAT I,TURSUN K,ROLFE B G,et al.Effect of early cold stress on the maturation of rice anthers[J].Proteomics,2004,4:1873-1882.
[8]曾維英,楊守萍,蓋鈞鎰,等.大豆質核互作雄性不育系NJCMS1A及其保持系的花藥差異蛋白質組學研究[J].中國農業科學,2007,40(12):2679-2687.
[9]文李,劉蓋,張再君,等.紅蓮型水稻細胞質雄性不育花粉總蛋白質初步比較分析[J].遺傳,2007,25(2):112-117.
[10]謝錦云,李小蘭,陳平,等.溫敏核不育水稻花藥蛋白質組的初步分析[J].中國生物化學與分子生物學報,2003,19(2):215-221.
[11]陳蕊紅,葉景秀,張改生,等.小麥質核互作型雄性不育系及其保持系花藥差異蛋白質組學分析[J].生物化學與生物物理進展,2009,36(4):431-440.
[12]SHEORANA I S,ANDREW R S R,DOUGLAS J H O,et al.Differential expression of proteins in the wild type and 7B-1 male-sterile mutant anthers of tomato(Solanum lycopersicum):a proteomic analysis[J].Proteomics,2009,71:624-636.
[13]YAMAMOTO M P,SHINADA H,ONODERA Y,et al.A male sterility-associated mitochondrial protein in wild beets causes pollen disruption in transgenic plants[J].Plant J,2008(6):1027-1036.
[14]YAMAMOTO M P,KUBO T,MIKAMI T.The 5'-leader sequence ofsugarbeetmitochondrialatp6 encodes a novel polypeptide that is characteristic of Owen cytoplasmic male sterility[J].MolGen Genomics,2005,273:342-349.
[15]YI F Y,SHI F L,LONG R C,et al.Comparative proteomic analysis of bud proteins of alfalfa male sterile and fertile plants at bud emergence stage[J].Acta Bot Boreal-Occident Sin,2013,33(10):1964-1971.
[16]張明珠,袁蕾,王俊生,等.順烏頭酸酶基因在殺雄劑SQ-1誘導小麥雄性不育花藥中的表達分析[J].中國生物化學與與分子生物學報,2010,26(8):740-748.
[17]FU J M,IVANA M I,VARA P P V.Roles of protein synthesis elongation factor EF-Tu in heat tolerance in plants[J].Journal of Botany,2012(10):835-836.
[18]YU Y,ZHANG X,ZHANG Y,et al.Isolation of the cDNA fragment of watermelon genic malesterility related gene s using DDRT2PCR[J].Journal of Northwest A&F University(Nat Sci Ed),2008,36(11):163-175.
[19]廖英明.紅麻蛋白質組2-DE建立及不育系和保持系差異蛋白分析[D].福建:福建農林大學,2010.
[20]劉祚昌,李繼耕,羅會馨,等.二磷酸核酮糖羧化酶與細胞質雄性不育性的研究[J].遺傳學報,1983,10(1):36-42.
[21]YUAN C Z,HAN X J,LI Y Q,et al.Genetic transform ation and biologicalfunction analysisofthe sedoheptulose-1,7-bisphosphatase gene from mulberry[J].Science of Sericulture,2013,39(3):413-0419.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
海峽科技與產業(2016年3期)2016-05-17 04:32:12
