Rock可能通過抑制PI3K/Akt通路促進(jìn)缺氧誘導(dǎo)的心肌細(xì)胞凋亡*
2014-08-13 12:39:58董家龍李菊香孫國(guó)芳
中國(guó)病理生理雜志 2014年11期
關(guān)鍵詞:信號(hào)
董家龍, 李菊香, 孫國(guó)芳, 洪 葵
(1南昌大學(xué)第二附屬醫(yī)院心內(nèi)科,江西省分子醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,江西 南昌 330006; 2贛州市南康區(qū)第一人民醫(yī)院心內(nèi)科,江西 贛州 341400)
Rho相關(guān)的卷曲螺旋蛋白激酶(Rho-associated coiled-coil protein kinase,ROCK)是新近發(fā)現(xiàn)的與細(xì)胞凋亡相關(guān)的激酶,是激活的caspase-3的直接裂解產(chǎn)物,ROCK有2個(gè)亞單位,即ROCK1和ROCK2。研究表明ROCK與心肌損傷、心肌細(xì)胞凋亡及心力衰竭有關(guān)聯(lián)[1]。然而,ROCK在缺血性心肌損傷中與磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)通路有何關(guān)系?目前尚不清楚。本研究旨在觀察缺氧誘導(dǎo)心肌細(xì)胞凋亡過程中,ROCK1和ROCK2的變化及其與心肌PI3K之間的關(guān)系。
材 料 和 方 法
1 材料
Sprague-Dawley乳鼠(出生1~3 d) 雌雄不限,由南昌大學(xué)醫(yī)學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物科學(xué)部提供(動(dòng)物合格證:醫(yī)動(dòng)字 021-9602)。DMEM高糖培養(yǎng)基及胎牛血清(FBS)購(gòu)自HyClone;胰蛋白酶購(gòu)自Sigma;山羊抗鼠ROCK-1Ⅰ抗、山羊抗鼠ROCK-2Ⅰ抗、兔抗鼠caspase-3Ⅰ抗、兔抗鼠p-PI3KⅠ抗、兔抗鼠PI3KⅠ抗均購(gòu)自Santa Cruz;抗α-橫紋肌肌動(dòng)蛋白免疫組化試劑盒購(gòu)自武漢博士德;MTS溶液購(gòu)自Promega;Annexin V-FITC細(xì)胞凋亡檢測(cè)試劑盒購(gòu)自南京凱基生物有限公司
2 方法
2.1原代心肌細(xì)胞的培養(yǎng)及鑒定 取健康乳鼠,利用0.08%胰酶消化3次后,離心棄胰酶,接著用1%II型膠原酶消化心肌組織1~2 h,至組織消化完全,以含15% FBS的DMEM培養(yǎng)基(1∶1)終止消化,室溫下離心5 min,收集細(xì)胞沉淀以15% FBS的DMEM培養(yǎng)基充分重懸,200目濾網(wǎng)過濾細(xì)胞懸液至培養(yǎng)皿,置CO2培養(yǎng)箱培養(yǎng),經(jīng)差速貼壁2 h后,加入0.1 mmol/L 5’-BrdU抑制成纖維細(xì)胞的生長(zhǎng),以1×109/L的細(xì)胞密度接種于35 mm的培養(yǎng)皿中,待心肌細(xì)胞生長(zhǎng)接近匯和狀態(tài)及呈現(xiàn)同步搏動(dòng)時(shí)開始實(shí)驗(yàn)。48 h后,取有心肌細(xì)胞生長(zhǎng)的玻片,用37 ℃ PBS漂洗,4%多聚甲醛固定,加大鼠抗α-橫紋肌肌動(dòng)蛋白單克隆Ⅰ抗(1∶50),于4 ℃濕盒過夜,加FITC羊抗小鼠IgG (1∶50),孵育30 min,PBS洗2次,DAPI染核,封片劑封片,顯微鏡下觀察。
2.2心肌細(xì)胞缺氧模型制備及各種條件培養(yǎng) 心肌細(xì)胞生長(zhǎng)接近匯合狀態(tài),呈現(xiàn)同步搏動(dòng)時(shí)開始實(shí)驗(yàn)。模擬缺血溶液(NaH2PO40.9 mmol/L, NaHCO36.0 mmol/L, CaCl21.8 mmol/L, MgSO41.2 mmol/L, HEPES 20 mmol/L,NaCl 98.5 mmol/L, KCl 10.0 mmol/L,乳酸鈉40 mmol/L, pH 6.5)以95% N2~5% CO2預(yù)飽和1 h,將培養(yǎng)的細(xì)胞以模擬缺血液置換正常培養(yǎng)基,放入密閉氣體盒中缺氧;以DMEM培養(yǎng)基于CO2培養(yǎng)箱中培養(yǎng),為有氧培養(yǎng)。實(shí)驗(yàn)分為5組:常規(guī)培養(yǎng)組、缺氧1 h組、缺氧3 h組、缺氧6 h組和缺氧9 h組。細(xì)胞經(jīng)實(shí)驗(yàn)干預(yù)后,檢測(cè)各時(shí)點(diǎn)相關(guān)指標(biāo):細(xì)胞凋亡率、細(xì)胞存活率及ROCK-1、ROCK-2、活化caspase-3、p-PI3K和PI3K的表達(dá)情況。
2.3MTT法檢測(cè)心肌細(xì)胞成活率 將細(xì)胞種植在96孔板中,空白調(diào)零孔只加不含細(xì)胞的培養(yǎng)基。實(shí)驗(yàn)處理后,每孔加入5 g/L MTT溶液20 μL,繼續(xù)培養(yǎng)4 h,棄上清,加入150 μL DMSO,振蕩10 min,酶標(biāo)儀(Labststems)測(cè)定492 nm處各孔吸光度(A)值。計(jì)算細(xì)胞存活率,細(xì)胞存活率=(實(shí)驗(yàn)組A/對(duì)照組A)×100%。
2.4流式細(xì)胞儀檢測(cè)細(xì)胞凋亡率 用胰酶消化收集上述各組細(xì)胞, PBS洗滌細(xì)胞2次,再用500 μL的Binding Buffer懸浮細(xì)胞,分別加入Annexin V-FITC和PI各5 μL,最后用FACS Calibar流式細(xì)胞儀(Beckton Dickinson,)檢測(cè)。
2.5全自動(dòng)生化分析儀檢測(cè)乳酸脫氫酶(lactate dehydrogenase,LDH) 各組實(shí)驗(yàn)干預(yù)后取心肌細(xì)胞培養(yǎng)液 400 μL,利用全自動(dòng)生化分析儀(Beckman)測(cè)定LDH活性。
2.6Western blotting 用RIPA法取總蛋白,BCA法測(cè)定蛋白質(zhì)濃度,取20 μg進(jìn)行SDS-PAGE分離蛋白。蛋白轉(zhuǎn)印到硝酸纖維素膜上,3% BSA液4 ℃封閉過夜。膜分別用ROCK1、ROCK2、caspase-3活化片段、p-PI3K和PI3K的Ⅰ抗 (1∶200) 孵育,4 ℃ 過夜, Ⅱ抗孵育2 h,DAB顯色照相。結(jié)果用LabWork 3.0 UVP軟件, 以目的條帶與β-actin的灰度值進(jìn)行分析。
3 統(tǒng)計(jì)學(xué)處理
實(shí)驗(yàn)均重復(fù)6次。用統(tǒng)計(jì)學(xué)軟件SPSS 12.0進(jìn)行分析,數(shù)據(jù)采用均數(shù)±標(biāo)準(zhǔn)差(mean±SD)表示,兩兩比較采用LSD-t檢驗(yàn)。以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
結(jié) 果
1 心肌細(xì)胞的鑒定
倒置顯微鏡下觀察:細(xì)胞呈片狀有節(jié)律的搏動(dòng),細(xì)胞致密,高度折光,細(xì)胞核小,常有1~2個(gè)核仁,有的細(xì)胞呈不規(guī)則的星形,并伸出偽足。免疫組化結(jié)果顯示,抗體陽(yáng)性心肌細(xì)胞胞漿內(nèi)有綠色熒光,見圖1。
2 心肌細(xì)胞存活率、凋亡率及上清液LDH的變化
缺氧1 h、3 h、6 h、9 h后,心肌細(xì)胞凋亡率和上清液LDH活性均顯著高于對(duì)照組(P<0.01),細(xì)胞存活率均低于對(duì)照組(P<0.01),見圖2、表1。
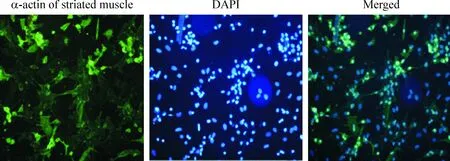
Figure 1. Cardiomyocyte identification observed by fluorescence microscopy(×100).

Figure 2. Cardiomyocyte apoptosis induced by hypoxia at different time points.

表1 不同時(shí)點(diǎn)心肌細(xì)胞存活率、凋亡率和上清液LDH活性的變化
3 心肌細(xì)胞中ROCK-1、ROCK-2蛋白表達(dá)
ROCK-1在缺氧1 h 即開始升高,在6 h 表達(dá)達(dá)高峰,在9 h開始回落,均明顯高于對(duì)照組(P<0.05)。ROCK-2與在缺氧1 h即開始升高,在3~6小時(shí)表達(dá)達(dá)高峰,在9 h開始降低,均明顯高于對(duì)照組(P<0.05),與ROCK-1的表達(dá)有平行對(duì)應(yīng)關(guān)系,見圖3。
4 心肌細(xì)胞中PI3K和p-PI3K的表達(dá)
PI3K在不同的缺氧時(shí)點(diǎn)表達(dá)無明顯變化,差別無統(tǒng)計(jì)學(xué)意義。p-PI3K蛋白的表達(dá)在缺氧1 h開始升高,在3 h左右達(dá)高峰,缺氧9 h后回落,差別有統(tǒng)計(jì)學(xué)意義,見圖4。

Figure 3. Expression of ROCK-1 and ROCK-2 in cardiomyocytes subjected to hypoxia for different time.1: control; 2: hypoxia 1 h; 3: hypoxia 3 h; 4: hypoxia 6 h;5: hypoxia 9 h. Mean±SD.n=6. *P<0.05 vs control.

Figure 4. Expression of PI3K and p-PI3K in cardiomyocytes subjected to hypoxia for different time. 1: control; 2: hypoxia 1 h; 3: hypoxia 3 h; 4: hypoxia 6 h; 5: hypo-xia 9 h. Mean±SD.n=6. *P<0.05 vs control.
5 缺氧后心肌細(xì)胞中caspase-3蛋白表達(dá)
Caspase-3活化片段在缺氧1 h開始逐漸升高,在隨后觀察的時(shí)點(diǎn)內(nèi)持續(xù)處于高表達(dá)狀態(tài),與對(duì)照組比較,差別有統(tǒng)計(jì)學(xué)意義(P<0.01),這與心肌細(xì)胞的凋亡率和培養(yǎng)液中的LDH的變化是一致的,見圖5。
討 論
研究表明,ROCK 是一種絲氨酸/蘇氨酸蛋白激酶,至少有2種亞型,即ROCK1和ROCK2。ROCK參與調(diào)節(jié)細(xì)胞的一些基本功能,如收縮、游走黏附、增殖和凋亡等[2],ROCK-1和ROCK-2幾乎在全身各組織都有表達(dá),兩者在心臟和血管平滑肌表達(dá)均較多,ROCK-2 mRNA在大腦和骨骼肌表達(dá)較多。靜息狀態(tài)下的ROCK沒有酶活性,因?yàn)镽OCK的激酶活性可能存在一種自我抑制的機(jī)制,即通過自身的折返使其催化中心(激酶域)覆蓋,這樣使得該激酶的激酶域與ATP的親和力受到抑制或使得其下游區(qū)域不能與底物結(jié)合而不被激活。ROCK的活性受細(xì)胞外信號(hào)和一些胞漿蛋白的調(diào)節(jié),ROCK接受Rho傳遞的活化信號(hào),發(fā)生多個(gè)氨基酸位點(diǎn)的磷酸化而激活,并介導(dǎo)其下游一系列磷酸化/脫磷酸化反應(yīng)。心力衰竭和冠心病發(fā)病過程中伴隨著心肌細(xì)胞凋亡[3],近年來,認(rèn)為ROCK與心肌細(xì)胞凋亡有關(guān)系[4]。研究發(fā)現(xiàn)在心肌細(xì)胞中,ROCK1可以被caspase-3激活,ROCK1是活化的caspase-3的直接裂解底物,而激活的ROCK1又可以反饋激活caspase-3,如此形成惡性循環(huán)[5]。

Figure 5. The expression of cleaved caspase-3 in cardiomyocytes subjected to hypoxia for different time. 1: control; 2: hypoxia 1 h; 3: hypoxia 3 h; 4: hypoxia 6 h; 5: hypoxia 9 h. Mean±SD.n=6.**P<0.01 vs control.
本研究發(fā)現(xiàn),缺氧損傷能引起心肌細(xì)胞凋亡增加及引起caspase-3活化,缺氧早期可以引起磷酸化的PI3K蛋白表達(dá)增加,這可能是心肌細(xì)胞抗凋亡的的保護(hù)機(jī)制,在繼續(xù)缺氧過程中磷酸化PI3K開始下降至基本消失。與此同時(shí),缺氧也可以引起ROCK-1和ROCK-2蛋白增加,并且在缺氧6 h左右表達(dá)達(dá)高峰,心肌細(xì)胞凋亡率也隨著缺氧時(shí)間而逐漸增加,p-PI3K在ROCK表達(dá)達(dá)高峰時(shí)開始回落, 提示ROCKs的激活可能抑制了p-PI3K的表達(dá),進(jìn)而促進(jìn)了缺氧心肌細(xì)胞的凋亡。
PI3K/Akt信號(hào)通路是近年來發(fā)現(xiàn)參與細(xì)胞增殖調(diào)控的重要信號(hào)通路,近來研究資料表明PI3K/Akt通路是許多生命活動(dòng)中關(guān)鍵的信號(hào)分子,PI3K/Akt介導(dǎo)的信號(hào)通路調(diào)節(jié)細(xì)胞的分裂、分化和凋亡等活動(dòng)。研究發(fā)現(xiàn)PI3K/Akt信號(hào)途徑在缺血預(yù)適應(yīng)、缺血再灌注損傷中被激活,進(jìn)而抑制心肌細(xì)胞凋亡,發(fā)揮心肌保護(hù)作用。研究表明PTEN是一種磷酸(酯)酶,是新近證實(shí)的ROCK的一種底物,其可以使磷酸肌醇底物去磷酸化,在細(xì)胞內(nèi)信號(hào)轉(zhuǎn)導(dǎo)中起了很重要的作用,尤其在PI3K/Akt信號(hào)通路中,它是PI3K上游區(qū)域的一個(gè)負(fù)性調(diào)節(jié)蛋白,與細(xì)胞生長(zhǎng)的調(diào)節(jié),蛋白質(zhì)的合成,轉(zhuǎn)錄子的調(diào)節(jié)及細(xì)胞存活有關(guān)。PTEN可以通過ROCK而磷酸化,進(jìn)而被激活。有文獻(xiàn)表明ROCK抑制劑保護(hù)心臟是通過抗細(xì)胞凋亡的作用,這是ROCK抑制劑減弱了缺血再灌注引起的Bcl-2下調(diào)及通過激活PI3K/Akt/eNOS途徑來保護(hù)心臟,這種保護(hù)作用可以被PI3K抑制劑LY294002完全阻斷;在心肌的局部缺血/再灌注損傷中,心肌中RhoA的表達(dá)及其后ROCK活性均有增加,在缺血再灌注早期給予ROCK抑制劑可以顯著減少SD大鼠心肌梗塞的面積及心肌細(xì)胞凋亡率[6]。綜上所述, ROCKs可能參與了缺氧致心肌細(xì)胞凋亡的過程,其機(jī)制可能是通過抑制p-PI3K信號(hào)途徑進(jìn)而促進(jìn)缺氧心肌細(xì)胞凋亡;ROCK可能通過激活caspase-3引起缺氧心肌細(xì)胞凋亡。
[參 考 文 獻(xiàn)]
[1] Fukui S, Fukumoto Y, Suzuki J, et al. Long-term inhibition of Rho-kinase ameliorates diastolic heart failure in hypertensive rats[J]. J Cardiovasc Pharmacol, 2008, 51(3): 317-326.
[2] Okamoto R, Li Y, Noma K, et al. FHL2 prevents cardiac hypertrophy in mice with cardiac-specific deletion of ROCK2[J]. FASEB J, 2013, 27(4):1439-1449.
[3] 米亞非,李小鷹,江 健,等. rAAV-1SERCA2a轉(zhuǎn)染改善心力衰竭犬心功能及其機(jī)制探討[J]. 中國(guó)病理生理雜志, 2014, 30(2): 208-213.
[4] Surma M, Wei L, Shi J. Rho kinase as a therapeutic target in cardiovascular disease[J]. Future Cardiol, 2011,7(5):657-671.
[5] Chang J, Xie M, Shah VR, et al. Activation of Rho-associated coiled-coil protein kinase 1 (ROCK-1) by caspase-3 cleavage plays an essential role in cardiac myocyte apoptosis[J]. Proc Natl Acad Sci U S A, 2006, 103(39):14495-14500.
[6] Jiang ZH, Zhang TT, Zhang JF. Protective effects of fasudil hydrochloride post-conditioning on acute myocardial ischemia/reperfusion injury in rats[J]. Cardiol J, 2013, 20(2): 197-202.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(hào)(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(hào)(2018年2期)2018-04-18 12:18:10
鐵道通信信號(hào)(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
中國(guó)病理生理雜志(2015年8期)2015-12-21 12:38:06

- 中國(guó)病理生理雜志的其它文章
- 艾塞那肽對(duì)新診斷2型糖尿病患者骨代謝的影響*
- 用阻塞性睡眠呼吸暫停低通氣綜合征動(dòng)物模型研究vaspin與胰島素信號(hào)通路的相關(guān)性*
- 白細(xì)胞介素4調(diào)節(jié)人臍帶間充質(zhì)干細(xì)胞的增殖和造血支持能力*
- 扶腎降濁方含藥血清對(duì)腎小管間質(zhì)損害大鼠成纖維細(xì)胞TGF-β1/Smads信號(hào)轉(zhuǎn)導(dǎo)通路的影響*
- AT1受體自身抗體對(duì)糖尿病腎病大鼠腎臟細(xì)胞凋亡及JNK表達(dá)的作用*
- 人肺動(dòng)脈平滑肌細(xì)胞中微小RNA-210通過MKP-1負(fù)性調(diào)節(jié)低氧下細(xì)胞的增殖*