黑龍江省番茄白粉病病原鑒定
2014-08-10 12:29:50朱路路李景富許向陽姜景彬
植物保護 2014年4期
李 帥, 朱路路, 李景富, 許向陽, 姜景彬
(東北農業(yè)大學園藝學院蔬菜系,哈爾濱 150030)
研究簡報
黑龍江省番茄白粉病病原鑒定
李 帥, 朱路路, 李景富*, 許向陽, 姜景彬
(東北農業(yè)大學園藝學院蔬菜系,哈爾濱 150030)
通過收集東北農業(yè)大學日光溫室中的番茄白粉菌,并對其進行形態(tài)和分子鑒定及分析,明確了該白粉病的病原種類及其分類地位。根據(jù)分生孢子的顯微形態(tài)觀察初步確定為新番茄粉孢菌(OidiumneolycopersiciL. Kiss )。進一步根據(jù)真菌rDNA-ITS序列檢測分析后發(fā)現(xiàn)該病原菌與新番茄粉孢菌(O.neolycopersici)的相似性為100%,并對與其相似性較高的序列構建進化樹分析發(fā)現(xiàn),本試驗研究的白粉菌與其他國家和地區(qū)的白粉菌相似性不高,這可能是由于哈爾濱地區(qū)白粉菌自身小種進化的原因。
新番茄粉孢菌; 形態(tài)特征; ITS; 進化樹
1986年,在荷蘭發(fā)現(xiàn)由番茄粉孢菌(Oidiumlycopersici)引起的番茄白粉病[1],隨后科研人員相繼報道了番茄白粉病在世界范圍內的蔓延[2-5],白粉病成為嚴重威脅番茄生產的全球性病害。1990年,賈菊生報道新疆發(fā)生番茄白粉病,經鑒定病原菌無性世代為OidiopsistauricaSalmon,有性世代為韃靼內絲白粉菌[Leveillulataurica(Lev.) Arn.][6]。黑龍江[7]、遼寧[8]、河南[9]、吉林[10]等地也有番茄白粉病的相關報道。我國已經報道能夠引起番茄白粉病的病原菌主要有蓼白粉菌(ErysiphepolygoniDC)[7]、韃靼內絲白粉菌[8]和新番茄粉孢菌(OidiumneolycopersiciL. Kiss)[9-10]引發(fā)。本試驗利用形態(tài)學和分子生物學方法,對東北農業(yè)大學日光溫室中白粉菌進行鑒定分析,明確該病原菌的種類及其分類地位,同時對哈爾濱地區(qū)白粉病病原菌進行鑒定以及對不同國家和地區(qū)的白粉病病菌生理小種進行進化樹分析,為白粉病的防治提供依據(jù)。
1 材料與方法
1.1 病原菌采集、純化及保存
2011年9月在東北農業(yè)大學園藝實驗站日光溫室的番茄(‘東農706’)葉片上采集白粉病病原菌。病原菌純化采用單孢直接挑取分離法[11]。挑取的單孢接種在番茄(‘Moneymaker’)幼苗上,置于光照培養(yǎng)箱(上海博迅SPX-250B-G)中培養(yǎng),光周期L∥D=16 h∥8 h,溫度20~25 ℃,相對濕度80%。
病原菌保存方法:將已發(fā)病葉片置于健康四葉期番茄幼苗上方,輕輕拍打葉柄部位,使粉狀物落在新葉片上完成接種,被接種葉片一般在接種后1周左右開始出現(xiàn)病癥[12]。
1.2 番茄白粉病的癥狀及病原菌形態(tài)特征觀察
觀察番茄植株發(fā)病癥狀,包括病斑顏色、大小和形狀。對病原菌形態(tài)進行觀察,包括菌絲形態(tài)和結構、分生孢子形態(tài)和萌發(fā)方式、分生孢子梗形態(tài)以及是否產生閉囊殼等。參考柴榮耀等的方法[11]進行病原菌種類的鑒定,使用AMG顯微照相記錄病原菌特征。
1.3 病原菌基因組DNA的提取、核糖體DNA的ITS區(qū)擴增及序列分析
將采自東北農業(yè)大學日光溫室的病原菌編號為Htpm1。試驗選取3株發(fā)病植株的葉片,用蓋玻片從發(fā)病葉片上分別刮取至少100 mg病菌混合物,液氮充分研磨,使用北京康為世紀公司細菌基因組提取試劑盒(Bacteria Gen DNA Kit)提取病原菌基因組DNA,-20 ℃保存。引物ITS4、ITS5、ITS6[12],其中ITS5-ITS6組合用于初始 PCR,ITS4-ITS5組合進行半巢式PCR擴增,引物由華大基因公司合成,引物序列見表1。
表1試驗使用引物信息
Table1Primersusedinthisstudy
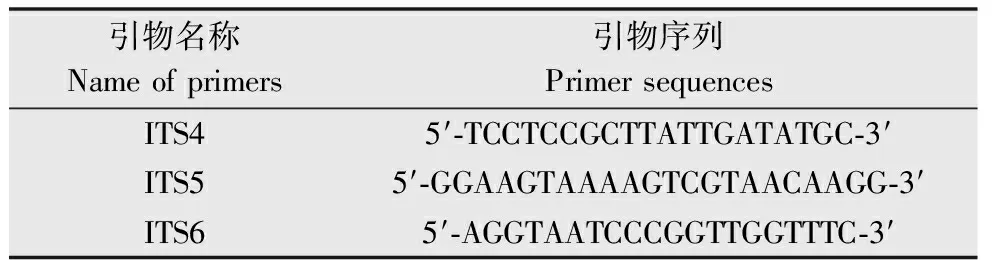
引物名稱Nameofprimers引物序列PrimersequencesITS45′?TCCTCCGCTTATTGATATGC?3′ITS55′?GGAAGTAAAAGTCGTAACAAGG?3′ITS65′?AGGTAATCCCGGTTGGTTTC?3′
PCR 反應體系(25 μL):Taq-PCR reaction mixture(15 μL)包括1.5 mmol/L MgCl2,800 μmol/L dNTPs,2.5UTaqDNA polymerase(TaqPCR Master Mix Kit; Qiagen, Tokyo, Japan),以及0.2 μmol/L 引物各1 μL,模板DNA 1 μL,無菌蒸餾水7 μL。
初始 PCR 反應程序:95 ℃ 預變性2 min;95 ℃變性1 min,55 ℃退火1 min,72 ℃延伸1 min,共30個循環(huán);最后72 ℃延伸2 min。半巢式PCR使用1 μL初始PCR產物作為模板,PCR程序同上,退火溫度調整為60 ℃。PCR產物4 ℃保存。
將3個 PCR 產物純化后送至華大基因(BGI)有限公司進行2次重復測序。將測得的序列在GenBank中進行BLAST分析,利用MEGA4.0軟件進行進化樹分析。
2 結果與分析
2.1 番茄白粉病癥狀
番茄白粉病多發(fā)于葉片、葉柄部位,病菌主要從下部葉片逐漸擴展到上部新生葉片。觀察發(fā)現(xiàn)菌斑僅在葉片上表面形成。發(fā)病初期(圖1a),葉片上表面有白色粉狀霉點,葉片發(fā)病部位出現(xiàn)褪綠點。粉狀霉點是病原菌的菌絲、分生孢子梗及分生孢子,起初分布稀疏,隨后逐漸融合在一起,增厚,形成不規(guī)則形狀并覆蓋大面積葉片。發(fā)病后期(圖1b)病葉變?yōu)楹稚饾u枯萎皺縮,但不脫落。
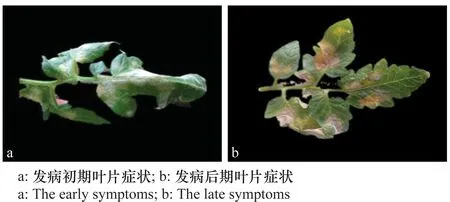
圖1 番茄白粉病發(fā)病癥狀
2.2 番茄白粉菌形態(tài)特征
分生孢子橢圓或圓柱形,無色,透明,平均32.2 μm×17.3 μm(n=150) (圖2a)。分生孢子單生于分生孢子梗頂端,分生孢子梗直立(圖2b),無分支,透明,平均71.5 μm×6.21 μm(n=100)。腳胞通常有1~2個細胞。菌絲上形成淺裂葉片形狀的附著胞(圖2c),芽管在分生孢子兩端或近端處萌發(fā),末端形成附著胞,附著胞多為乳突形 (圖2d)。未發(fā)現(xiàn)有閉囊殼。觀察結果與Kiss所述相吻合[3],因此鑒定該病原菌為新番茄粉孢菌(OidiumneolycopersiciL. Kiss)。

圖2 番茄白粉菌的分生孢子梗和分生孢子
2.3 番茄白粉菌核糖體DNA的ITS區(qū)擴增及序列分析
對提取的番茄白粉菌DNA的ITS區(qū)進行PCR擴增,所得片段長度為687 bp,GenBank登錄號為JQ972700。BLAST分析表明與新番茄粉孢菌(O.neolycopersici)相似性為100%,進一步證明該菌為新番茄粉孢菌。將得到的ITS序列與NCBI上相似性較高的序列進行比對,采用MEGA4.0軟件進行聚類分析,檢測方法用自展法(Bootstrap)。分析結果如圖3所示,所有序列分為幾個大支,我國臺灣地區(qū)、西班牙和我國哈爾濱地區(qū)的白粉病菌分別單獨為一支,可能是其自身小種進化的原因。匈牙利、美國和英國番茄白粉菌的相似性極高,法國、韓國、中國河南地區(qū)、荷蘭、日本和中國遼寧地區(qū)的番茄白粉菌相似性也極高。這種差異說明地域的不同并非是白粉菌生理小種變異的直接原因。
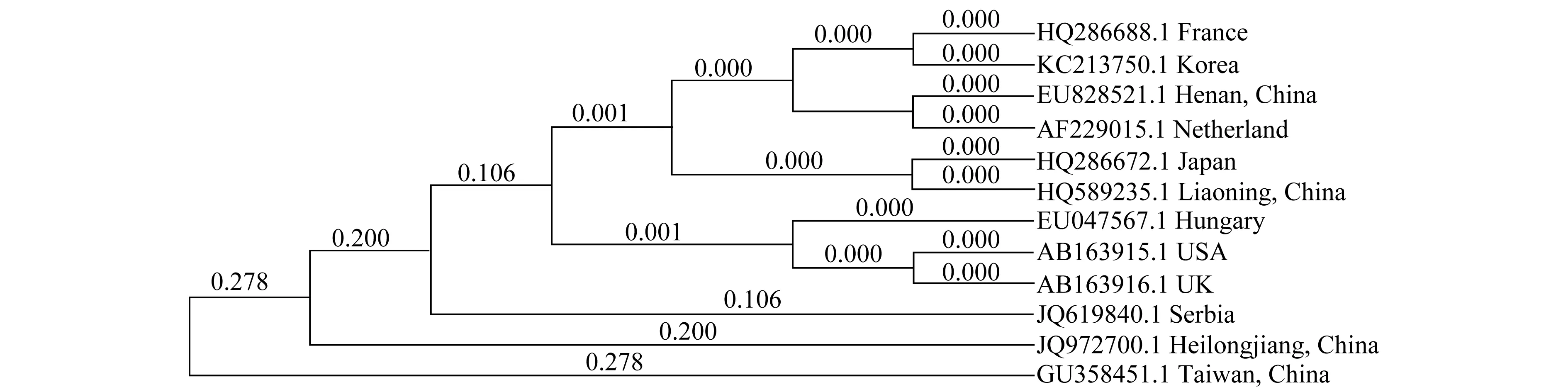
圖3 根據(jù)ITS序列構建的番茄白粉菌系統(tǒng)進化樹
3 討論
白粉病一直是番茄育種和生產中的主要病害之一,高溫高濕是適宜白粉菌生長的條件,不同的生態(tài)地理環(huán)境容易引發(fā)白粉菌生理小種的變異[13-15]。Kiss報道引起番茄白粉病的病原菌有兩個種,即番茄粉孢菌(O.lycopersici)和新番茄粉孢菌(O.neolycopersici),但至今未發(fā)現(xiàn)該菌的有性世代[3]。國內有關番茄白粉病的研究多見于發(fā)生情況和病原鑒定[10],黑龍江省未有新番茄粉孢菌引起番茄白粉病的相關報道,本研究首先對東北農業(yè)大學日光溫室的白粉菌進行形態(tài)觀察,初步鑒定結果顯示引起哈爾濱地區(qū)番茄白粉病的病原菌為新番茄粉孢菌,進一步利用真菌rDNA-ITS序列的保守性對采自哈爾濱的番茄白粉病病菌進行PCR擴增檢測分析,從分子水平上驗證了此菌為新番茄粉孢菌。通過構建進化樹發(fā)現(xiàn),哈爾濱與國內其他地區(qū)以及國外的白粉菌均有差異,但差異很小,法國、荷蘭、匈牙利、日本、韓國、法國等國家番茄白粉菌的相似性極高,種間差異很小,這說明地域差異并不是白粉菌生理小種產生差異的直接原因,可能與其自身分化有關。
本試驗從形態(tài)和分子兩方面對番茄白粉菌進行觀察鑒定,并對相似性高的序列進行系統(tǒng)進化分析,以期對番茄白粉病菌有更深入的了解,并為番茄白粉病的防治提供重要的理論依據(jù)。
[1]Paternotte S J. Echte meeldauw in tomaat geen echte bedreiging[J]. Groenten en Fruit,1988, 43:30-31.
[2]Neshev G. Powdery mildew (Oidiumsp.) on tomatoes in Bulgaria[J]. Phytoparasitica,1993, 21:339-343.
[3]Kiss L. Occurrence of a new powdery mildew fungus (Erysiphesp.) on tomatoes in Hungary[J]. Plant Disease,1996, 80:224.
[4]Olalla L,Tores J A.First report of powdery mildew of tomato caused by anErysiphesp. in Spain[J]. Plant Disease,1998, 82:592.
[5]Marois J J, Momol M T, Kimbrough J W. First report of powdery mildew on greenhouse tomatoes caused byOidiumneolycopersiciin Florida[J]. Plant Disease,2001, 85:1292.
[6]賈菊生.新疆番茄病害一新記錄-番茄白粉病[J].植物保護,1990,16(4): 5.
[7]程志明.黑龍江番茄新病害番茄白粉病[J].北方園藝,1992(6):40.
[8]王媛媛,陳立杰,段玉璽,等.沈陽地區(qū)溫室番茄發(fā)生白粉病[J].植物保護,2004,30(5):91.
[9]Li C W, Pei D L, Wang W J, et al. First report of powdery mildew caused byOidiumneolycopersicion tomato in China[J]. Plant Disease, 2008, 92:1370-1370.
[10]劉微,劉淑艷,李玉,等.番茄白粉病的病原菌鑒定[J].植物病理學報,2009,39(1): 11-15.
[11]柴榮耀,金敏忠.介紹一種單孢直接挑取獲得單孢菌株的方法[J].植物保護,1991,17(5):53.
[12]Whipps J M, Budge S P, Fenlon J S. Characteristics and host range of tomato powdery mildew[J]. Plant Pathology,1998, 47:36-48.
[13]Yoshinori M, Takeshi S, Nobuyuki M, et al. Identification of individual powdery mildew fungi infecting leaves and direct detection of gene expression by single conidium polymerase chain reaction[J]. Phytopathology,2005, 95:1137-1143.
[14]Kusaba M, Tsuge T. Phylogeny of alternaria fungi known to produce host specific toxins on the basis of variation in internal transcribed spacers of ribosomal DNA[J].Current Genetics,1995, 28:491-498.
[15]Kiss L, Cook R T A, Saenz G S, et al. Identification of two powdery mildew fungi,Oidiumneolycopersicisp. nov. andO.lycopersici, infecting tomato in different parts of the world[J]. Mycological Research,2001, 105 (6): 684-697.
IdentificationofthepathogencausingtomatopowderymildewinHeilongjiangProvince
Li Shuai, Zhu Lulu, Li Jingfu, Xu Xiangyang, Jiang Jingbin
(DepartmentofVegetables,CollegeofHorticulture,NortheastAgriculturalUniversity,Harbin150030,China)
Powdery mildew was observed on tomato (SolanumlycopersicumL.) in the greenhouse in Northeast Agricultural University. In order to clear the pathogenic species and its classification status, the pathogen was identified by morphological and molecular identification. The conidia morphological observation demonstrated that the pathogen might beOidiumneolycopersiciL.Kiss. Moreover, Sequence analysis of the rDNA-ITS region showed that the identity was 100% withOidiumneolycopersici. Phylogenetic analysis showed that the pathogen of tomato powdery mildew in Harbin has low homology with that in other regions and countries, indicating it may be due to the evolution of the pathogen race.
Oidiumneolycopersici; morphological characteristics; ITS; phylogenetic tree
2013-09-07
:2014-01-25
“十二五”國家科技支撐計劃(2012BAD02B02-7)
S 482.2
:ADOI:10.3969/j.issn.0529-1542.2014.04.022
* 通信作者 E-mail: lijf_2005@126.com
