柴達木盆地諾木洪地區5種優勢荒漠植物水分來源
2014-08-08 02:15:12朱建佳陳同同
生態學報 2014年21期
邢 星,陳 輝,*,朱建佳,2,陳同同
(1. 河北師范大學資源與環境科學學院,河北省環境演化與生態建設實驗室, 石家莊 050024;2. 中國科學院地理科學與資源研究所, 北京 100101)
柴達木盆地諾木洪地區5種優勢荒漠植物水分來源
邢 星1,陳 輝1,*,朱建佳1,2,陳同同1
(1. 河北師范大學資源與環境科學學院,河北省環境演化與生態建設實驗室, 石家莊 050024;2. 中國科學院地理科學與資源研究所, 北京 100101)
通過測定柴達木盆地諾木洪地區5種荒漠植物木質部水分及其不同潛在水源的穩定性氫氧同位素值,利用多源線性混合模型(IsoSource)分析了不同水分來源對荒漠植物的貢獻率。結果表明:當地大氣降水線為y=7.019x-3.217 (R2=0.970,P<0.001),很好地反映了該地區氣溫高、濕度低的氣候特點。諾木洪地區5種優勢植物整個生長季使用土壤水比例最大,其次為地下水。駝絨藜使用10—50 cm土壤層水分,白刺、檉柳利用50—70 cm土壤層水分比例最大,這兩種植物存在對50—70 cm土壤層水分的競爭;麻黃和沙拐棗對各層土壤水分的利用比例較為平均,因此存在對各層土壤水的競爭現象。4種灌木白刺、麻黃、檉柳、沙拐棗在生長季對不同水源的利用存在轉換,但生長季末期都對地下水利用比例逐漸增大。地下水是荒漠植被的重要水源,因此維持干旱半干旱地區地下水水位對荒漠植物的生長具有重要意義。植物根系貫穿于整個土壤剖面,但是根系分布與其吸水位置不完全對應,過去利用根系結構進行植物水分來源判斷的方法存在一定的局限性。
柴達木盆地; 諾木洪;荒漠植物;水分來源;氫氧同位素
水分是維持植物生命系統的重要物質,水分來源的選擇便成為植物生命周期中的重要選擇過程。植物水分來源包括地下水、地表水、土壤水等等,但是大氣降水卻是這些水分的最終來源[1]。雖然植物所吸收利用的水分均來自于降水,但是由于蒸發因素、海拔高度差異、蓄水層地質特征、土壤對水分吸收溶解濃度的差異等因素,都會造成水中δ18O、δD的不同[2- 3]。
有研究認為,水分在從根系到葉片的蒸騰作用下會在葉片中產生同位素的富集[4],少部分排鹽的鹽生植物根系會在從土壤吸收水分的過程中發生同位素的分餾[5],但大多數植物在這一過程中水分的氫氧同位素不會發生分餾[6- 8]。而且,水分在植物莖中向上傳遞的過程中,利用木質部的運輸,不存在穩定同位素分餾的現象[9]。因此,通過對比植物莖木質部與各個來源水分的δ18O、δD值,便可以找出該種植物是運用何種來源的水分進行生長[10],為利用穩定性同位素來分析植物水分來源提供了理論基礎。
穩定同位素技術到20世紀70年代才應用到生態學領域,該項技術在確定植物群落水分來源方面得到了廣泛的應用,近年來許多學者致力于對干旱區植物水分來源研究。新的研究發現:干旱區不同海拔梯度條件下喬木、灌木都有不同程度的利用夏季降水的狀況[11]。研究發現:灌溉前后幼齡檉柳表現出對水源利用的轉換,而成年檉柳則對灌溉反映不敏感[12]。降水較多的樣地,植物更傾向于利用淺層土壤水[13];氣候越干旱,植物越傾向于利用深層土壤水以及地下水[14]。也有研究認為,不同季節植物對水源的利用比例也存在轉換[15- 16]。但是地處青藏高原的柴達木盆地,荒漠生態系統對全球變化具有敏感響應和強烈影響[17],至今也未見對柴達木盆地植物水分來源報道,并且在以往的研究中,并沒有注意到河水、降水對植物水分來源的影響。柴達木盆地優勢植物在整個生長季對不同水源的利用比例如何,生長季初期和末期水分利用比例是否一致以及優勢種之間是否存在水源競爭等問題也并未知曉。
本文對柴達木盆地東部諾木洪地區5種荒漠植物水分來源進行定量分析,通過荒漠植物水分利用的總體特征、季節規律和物種間的差別,研究柴達木盆地植物水分利用機理,探討干旱地區植物適應環境策略,為我國干旱半干旱地區荒漠生態系統的維護和穩定提供基礎理論依據。
1 材料和方法
1.1 研究區概況
研究區位于我國青藏高原東北部,青海省海西蒙古族藏族自治州都蘭縣境內(圖1)。屬于高原大陸性氣候,平均海拔較高,氣候干旱,生長季較短,集中在每年的6—9月。根據全國氣象站點的逐日數據計算得出研究區56年(1957—2012)平均氣溫4.9 ℃,多年平均降水量43.5 mm。2010年均溫6.2 ℃,植物生長季6—9月降水量分別為81、17.2、19、14.8 mm,生長季降水總量為132.0 mm,占年降水量(149.0 mm)的88.6%,蒸發量2800 mm遠大于降水量。土壤多以荒漠土壤為主,植被則以抗旱能力好的荒漠灌叢為主。諾木洪樣地采樣點位于: 36°22′49.26″ N, 96°24′36.06″ E,海拔2858 m。
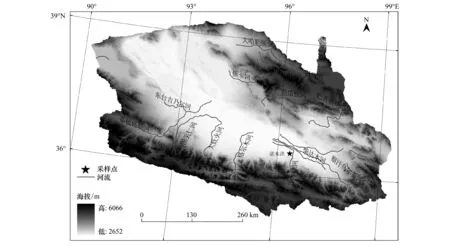
圖1 研究區及采樣點位置圖Fig.1 Location of the study area and the sampling site
1.2 實驗方法與樣品分析
1.2.1 取樣方法
植物樣品包括當地所有優勢種:駝絨藜(Ceratoideslatens)、白刺(Nitrariatangutorum)、麻黃(Ephedrasinica)、檉柳(Tamarixramosissima)、沙拐棗(Calligonummongolicum)。
2010年6—9月每15d采集1次樣品,采樣開始時間為6月15日,結束時間為9月30日。為了盡量減少光照強度等外界條件對同位素分析結果的影響,采樣過程保證在9:00到11:00間完成。在樣地選擇生長狀況較良好、高度較為一致的植株作為植物莖樣品的采集對象。選取5棵有代表性的駝絨藜、白刺、麻黃、檉柳、沙拐棗,在朝向一致的大枝條上選取一根小的樹枝,截取直徑0.2 cm左右的栓質化小莖,快速裝入已經準備好的4 mL螺紋玻璃樣品瓶中,擰緊瓶蓋。在上述植物分布的樣點附近選擇地勢相對均一的地方挖3個1 m深的土壤剖面,分別采集0—10 cm、10—30 cm、30—50 cm、50—70 cm、70—90 cm土層的土壤,裝入4 mL螺紋樣品瓶中,擰緊瓶蓋。植物取樣同時收集距離取樣點最近的24 m深井井水代表樣點的地下水。該井位于36°25′48″ N,96°24′36″ E,距離采樣點大約5.6 km。采集樣地附近河水,攪動表層水以保證樣品的均質性。收集取樣前最近一次降水,裝入取樣瓶中。所有樣品均取3個重復,用封口膜密封冷藏保存,共采集樣品306個。
1.2.2 樣品分析
先使用低溫真空抽提法抽提植物莖和土壤樣品中的水分,過0.22 μm濾紙后放入2 mL樣品瓶。水樣直接過0.22 μm濾紙放入2 mL樣品瓶待測。本次實驗中全部水樣使用Picarro L2130-i 超高精度液態水和水汽同位素分析儀進行同位素的測定。液態水測量的典型精度δ18O為 0.011‰;δD為0.038‰。該儀器自帶ChemCorrect有機污染物校驗軟件可以通過光譜分析對不同污染物進行標記,從而得出該結果是否可靠。存在有機污染的全部植物莖樣品以及部分土壤樣品在中國科學院地理科學與資源研究所同位素質譜儀Finnegan MAT- 253分析。分析得出的δD、δ18O是同標準平均海洋水SMOW的千分差。
δX=[R樣品/R標準-1] ×1000
式中,R樣品為待測樣品的氫或氧同位素自然豐度比,R標準為標準海洋水SMOW的氫或氧同位素自然豐度比。隨機抽取諾木洪地區無有機污染的樣品17個同時用Picarro和質譜儀測定,測定結果采用Anova單因素方差分析顯示:δ18O和δD均值分別為-6.577‰、-6.173‰和-58.118‰、-53.253‰,兩種測定方法結果無顯著性差異(P>0.05)。土壤含水量使用Decagon公司Em50Em50R儀器進行測定。
1.2.3 數據分析
不同土壤層含水量以及土壤水δ18O的差異用SPSS 17.0統計軟件進行分析。不同水源對植物的貢獻率用多源線性混合模型(IsoSource)分析,計算時增幅為1%,不確定水平為0.1%。
2 結果與討論
2.1 諾木洪降水同位素特征
Craig[18]根據全球降水中氫氧同位素存在的線性關系,通過計算得出全球尺度下二者之間的關系,將其定義為大氣水線(Meteoric Water Line,即MWL):δD=8δ18O+10。Dansgaard提出過量氘(deuterium excess)的概念:d=δD-8δ18O。d值在全球尺度下為10,即全球尺度下大氣水線的截距。過量氘反映了降水形成時局部地理要素,以及海水蒸發時的條件,因此比較普遍地運用到降雨水汽來源的研究中。
根據測得的諾木洪降水樣品中穩定性氫氧同位素的值,得出該地大氣水線y=7.019x-3.217(R2=0.970,P<0.001) (圖2),斜率為7.019,截距為-3.217,斜率小于全球平均值說明該地區氣候干燥,截距呈現負值說明該地區氣溫高、濕度低。對諾木洪地區進行過量氘計算,均值為(5.674±3.18)‰,低于大部分地區雨水d值(10‰左右)說明本地夏季水汽輻合較強,在干燥氣候條件下形成降雨,水滴降落過程蒸發強烈。通過對諾木洪地區植物莖、河水、分層土壤水中δD—δ18O分布點與該樣地大氣降水線進行比較,分析結果表明:由于地下水位較淺,河水入滲量較大,地下水、河水δ18O值偏小,均位于大氣降水線的上方[19]。土壤水、植物水的δ18O、δD值多位于該地大氣降水線右下方,說明該地區存在強烈的蒸發作用,和降水相比重同位素富集明顯。圖中土壤水、地下水、河水除少量點距離大氣降水線較遠外,其余都分布于大氣降水線附近,說明諾木洪地區降水對土壤水、地下水、河水等都有補給作用[20]。
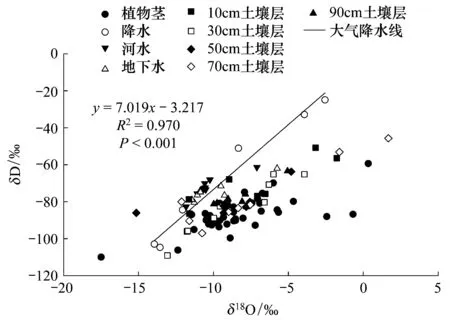
圖2 諾木洪植物莖水以及各種水分來源δD—δ18O與當地大氣降水線關系比較Fig.2 Relations between δD and δ18O values of plant stem water, precipitation, soil water, river water, groundwater, and local meteoric water line
2.2 諾木洪土壤含水量及不同水體δ18O的特征
由圖3可知諾木洪在2010年生長季降雨事件較多,但是大于15 mm的降水只有兩次:6月7日和8月3日,降水量分別為52 mm和16.7 mm。由于夏季水汽輻合能力較強,雨滴在降落過程中蒸發強烈,使得δ18O較高的局地水汽進入降水云團,致使降水中過量氘較低,而δ18O較高。降水量的變化對土壤、植被、水文等方面都會產生影響。降水可以直接補充土壤中的水分,降水量的大小對水分的補充有效程度卻不同,有研究認為小于5 mm的降水不能對土壤水分起到有效的補充,而只能降溫和緩解旱情[21]。杜建會[22]等人的研究認為甘肅民勤地區發生47.6 mm的強降雨事件后,發育中和活化階段的沙堆補給深度可達50 cm,穩定階段的白刺沙堆也可以達到30 cm。2010年諾木洪地區生長季的降雨總量達到132.0 mm,尤其是6月7日的降水,超過有歷史記錄以來的日降水極值,達到暴雨級別,對土壤水分進行了很好的補給。對比諾木洪地區土壤剖面δ18O與降水δ18O值(圖5),降水δ18O與50 cm土壤水δ18O值最接近,因此筆者認為諾木洪降水補給深度可以達到50 cm。
對比不同土層土壤含水量(圖4),50 cm土壤層含水量較高(8.5%),10 cm、30 cm以及70 cm層土壤含水量較為接近,90 cm含水量最少(7.1%)。諾木洪地區河水中δ18O的變化范圍是:-11.823‰—-7.101‰,平均值為(-10.316±0.491)‰(Mean±SE);地下水樣品中δ18O的變化范圍是:-11.437‰—-5.752‰,平均值為(-10.009±0.671)‰(Mean±SE)。河水以及地下水δ18O波動范圍較小。由于地下水較為穩定,并且循環周期慢,其中δ18O值較河水波動小。
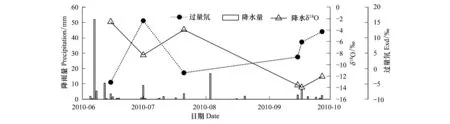
圖3 諾木洪2010年6—9月降水量、降水δ18O及過量氘Fig.3 Precipitation,δ18O values and deuterium excess from June to September, 2010 in Nuomuhong area

圖4 諾木洪土壤剖面含水量變化Fig.4 Variations of soil water content along soil profiles in Nuomuhong area
對諾木洪地區河水、地下水以及土壤水分析發現(圖5),河水中δ18O與地下水、土壤水相比偏負。
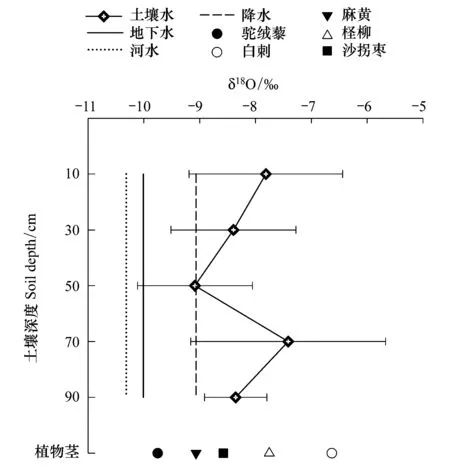
圖5 諾木洪不同植物莖及各水源δ18O均值Fig.5 δ18O values profiles of different plant stem water and water sources in Nuomuhong area
造成這一現象的原因可能有兩個:一是河水在入滲補給地下水和土壤水的過程中會有分餾作用導致重同位素的富集;二是當河水已經轉化為土壤水之后,土壤剖面中的水分也會受到蒸發的影響,δ18O值因此升高[23]。諾木洪地區河水的補給主要來自于南部昆侖山冰雪融水[24],本次實驗在玉珠峰采樣的冰川水δ18O值為-11.2‰,與諾木洪河水δ18O值很接近。本研究區內分層土壤中的δ18O值在0—10 cm表層土壤中以及50—70 cm土壤層值偏高。0—10 cm層δ18O偏高的原因是干旱地區表層土壤水通過δ16O強烈蒸發而使重同位素δ18O在表層土壤剖面相對富集;50—70 cm土壤層δ18O偏高可能是由于該土層水被植物利用較多。30—50 cm層土壤水δ18O變小,結合土壤含水量進行分析,由于含水量的增多導致δ18O值下降,降水δ18O值同30—50 cm土壤層重合,說明雨水能補給到該土壤層30—50 cm深處。田立德[25]等在研究青藏高原那曲地區時發現,大約7d之前的降水δ18O值與土壤層20—30 cm處δ18O相對應。
2.3 植物水分來源分析
2.3.1 植物利用各種來源水分定量分析
將采集到的所有水源樣品加入分析,對比每個采樣日期每種植物對水分的吸收利用情況(圖6)。所有植物對水分利用來源的選擇顯示(圖7),土壤水是植物水分利用的一個重要來源,其次是較為穩定的地下水。對于河水以及降水的分配比例出現了兩種情況,駝絨藜、麻黃、檉柳使用河水大于降水,白刺、沙拐棗使用降水大于河水。

圖6 諾木洪所有樣品δ18O值Fig.6 δ18O values of all samples
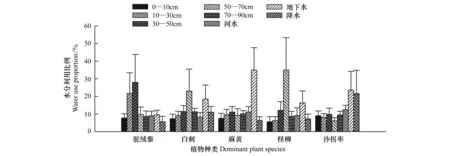
圖7 諾木洪地區優勢種生長季平均水分利用比例Fig.7 The water use proportion of dominant plant species in growth season in Nuomuhong area
分析5種植物在不同季節水源利用變化(表1)發現,在生長季初期和末期,駝絨藜對各種來源水分利用比例較為平均,6月15日對10—30cm土壤層水的利用達到67.6%,可能與6月7日強降雨的持續影響有關。生長旺季主要利用30—50 cm土壤水;白刺在生長季初期使用50—70 cm土壤層中的水分,生長旺季最多利用地下水,其次為表層土壤水,生長季末期則對地下水利用比例顯著增大;麻黃在生長季初期對各來源水分利用比例較為平均,生長旺季開始對地下水利用比例逐漸升高,到生長季末期則對地下水利用比例升高至71.6%;檉柳在生長季初期對50—70 cm土壤層中的水分利用比例最多,生長旺季可同時利用50—70 cm土壤水以及地下水,到生長季末期則更傾向于使用地下水;沙拐棗在生長季初期對各種來源水分利用較為平均,生長旺季對降水(31%)以及地下水(25.7%)平均利用比例較大,生長季末期對地下水利用比例也達到24.6%。
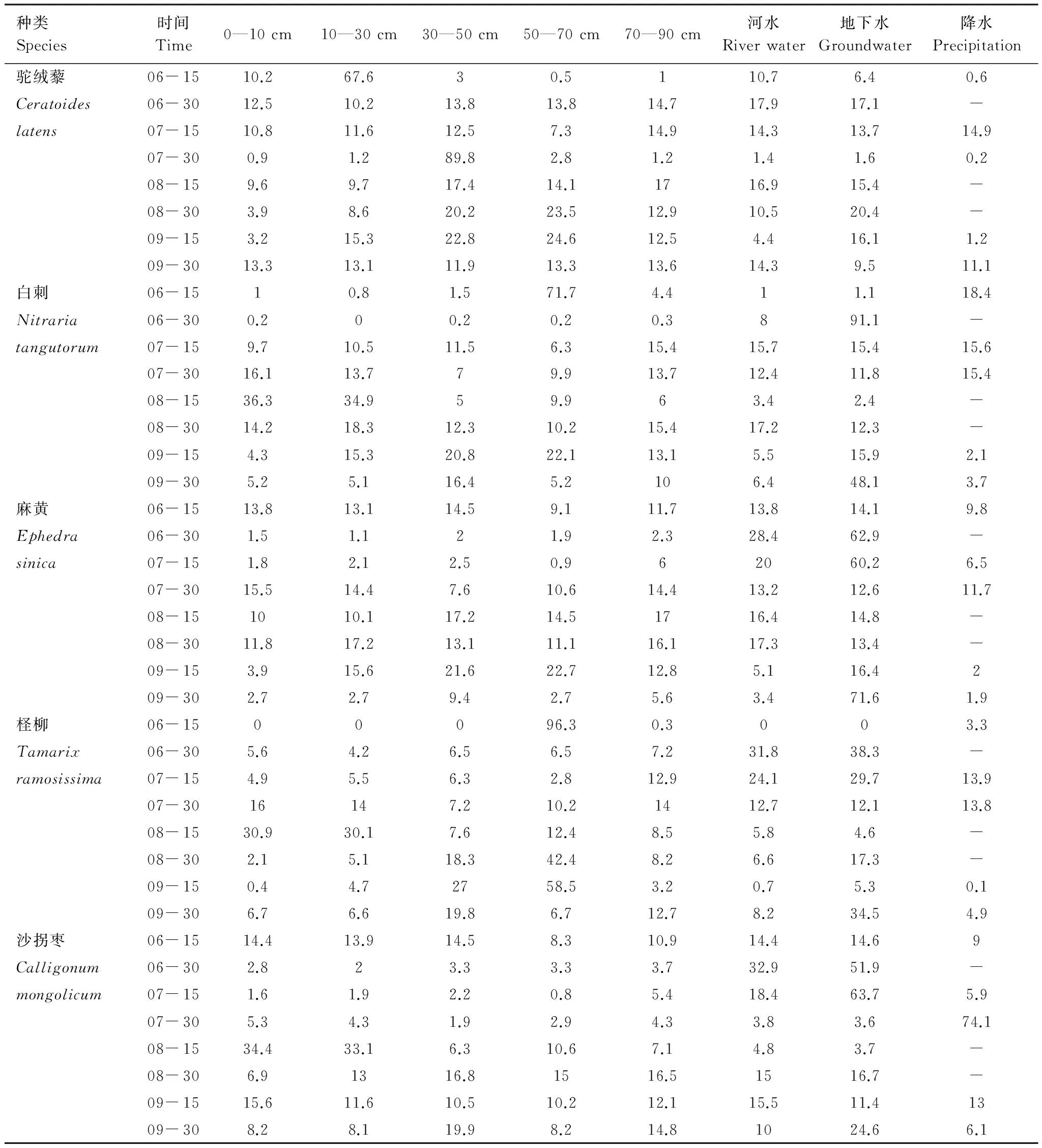
表1 諾木洪地區5種優勢植物生長季水分利用比例/%
植物對資源的競爭是一種普遍現象,多數生態系統的競爭主要發生在地下根系對水分和養分的競爭[26- 27],同一生境中吸水層位相似的植物間存在對水源的競爭現象[28]。因此,在生長季初期,白刺和檉柳對50—70 cm土壤層中的水分存在競爭,駝絨藜、麻黃和沙拐棗則對每層土壤水都有競爭現象;生長旺期,4種灌木白刺、麻黃、檉柳和沙拐棗開始對地下水競爭,到生長末期,對地下水源的競爭加劇。
2.3.2 植物利用土壤水分定量分析
對諾木洪地區5種植物土壤水分利用比例進行深入分析。駝絨藜在整個生長季對表層0—10 cm土壤層利用比例最少,其次為50—70 cm土壤層,對30—50 cm土壤層中的水分利用比例最高,平均可以達到32.33%。白刺利用50—70 cm土壤層中的水分最多。麻黃對各層土壤水分利用比例較為平均。檉柳對于50—70 cm的土壤層利用比例最多,情況與白刺相似。沙拐棗對各土壤層中水分利用比例較為均勻,沒有出現對某層水分的過于依賴。
植物生長于土壤之中,土壤水是植物水的直接來源。植物根系吸收土壤水分這一過程,控制著水分狀態和光合作用等[29]。對于水分稀少的荒漠地區來說,荒漠灌木根系表現型或者空間結構特征是劃分根系功能的重要依據[30]。駝絨藜在土壤水使用方面與其他植物不同。駝絨藜屬于半灌木,而白刺、麻黃、檉柳、沙拐棗都屬于灌木。白刺主根粗壯,入土較淺,根系以0—40 cm土層比例較大,但是平均根深80 cm,甚至1 m以上[31]。麻黃主根發達,毛根分布范圍廣[32],根系發達,呈網絡狀固結土壤[33]。深根系植物檉柳,在天山地區的挖掘發現其主要根系分布范圍為50—160 cm,最深可達300 cm,0—60 cm根系分布較少[34]。檉柳對50—70 cm以及深層地下水利用比例較高,同徐貴青,朱林[35]等的研究結果一致。沙拐棗根系較深,側根在10—20 cm層分布最多,吸水能力也更強[36]。前人對沙拐棗的研究中認為,其水平與垂直根系均發達,可以利用到深層土壤水。在本研究中發現該植被對地下水源的利用比例很大,這一現象同麻黃相似。因此,植物根系可能會存在于整個土壤剖面,但是根的存在卻并不意味著對土壤水分吸收活躍。植物用來吸收水分的活根往往會因為外界條件的刺激而改變。4種灌木白刺、麻黃、檉柳、沙拐棗根系都比較深,在整個生長季對不同水源的利用比例卻存在差異。生長季初期白刺、檉柳對50—70 cm土壤水利用比例最大,麻黃和沙拐棗則對各種水源利用比例較為平均,但是在生長季的末期,這4種植物都對地下水利用比例最高。這一結果表明,過去利用根系結構進行植物水分來源判斷的方法存在一定的局限性。植物根系分布的位置,不一定是其吸收水分的位置。
地下水是多數荒漠植物重要的水分來源[37]。地下水位下降可以直接影響土壤水分和養分分布,引起植物對水資源競爭,從而影響荒漠植被分布、生長、種群演替等群落格局[38]。樊自立等認為西北干旱區生態地下水埋深深度可以在很大程度上影響非地帶性中生和中旱生植被[39]。楊自輝[40]等通過對民勤地區地下水和降水監測研究白刺群落消長,發現地下水位的下降驅動綠洲外圍環境的旱化。劉發民[41]等認為巴丹吉林沙漠西南緣檉柳可在0—200 cm范圍內土壤含水量較低的條件下生存。梁少民[42]等對塔克拉瑪干腹地灌水量分析認為,根系區土壤水分低于2.5%沙拐棗就會受到干旱脅迫。諾木洪地區系潛水,隨著城市化建設進程加快,用水量逐漸加大。在本研究中發現,生長季末期,本研究區植物對地下水利用比例普遍較高,因此維持地下水水位將對本區植物的生長極為重要。
3 結論
(1) 諾木洪地區大氣降水線為y=7.019x-3.217 (R2=0.970,P<0.001),很好地反應了干旱區的氣候特點。斜率小于8,截距為負,表明該地溫度高濕度低。大氣降水過量氘值較低,表明本地降水中來自局地蒸發的水分較多。諾木洪地區河水、地下水、土壤水都位于大氣降水線附近,說明降水都對他們有補給作用。河水主要來源于山區冰雪融水,對地下水補給量大,所以δ18O值較小,位于大氣降水線左上方;此地氣候干旱蒸發強烈,造成土壤水中δ18O的富集,所以位于大氣降水線下方。
(2) 諾木洪地區5種優勢種植物都使用土壤水比例最大,其次為地下水。半灌木駝絨藜使用10—50 cm層土壤水分,灌木白刺、麻黃、檉柳、沙拐棗在土壤水分利用比例方面存在顯著不同,白刺、檉柳吸水層位相同,都對50—70 cm土壤層水分吸收最大,存在對該層水分的競爭;麻黃和沙拐棗對每層土壤水分的利用比例較為平均,出現對土壤水分的競爭現象。
4種灌木白刺、麻黃、檉柳、沙拐棗生長季的不同時期都存在對利用水源的轉換現象,但是到生長季末期,都對地下水的需求較多。因此維持干旱半干旱地區地下水水位對荒漠植物的生長具有重要意義。
(3) 植物根系貫穿于整個土壤剖面,但是根的存在卻并不意味著對土壤水分吸收活躍。植物用來吸收水分的活根往往會因為外界條件的刺激而發生改變。因此,植物根系分布的位置,不一定是其吸收水分的位置。過去利用根系結構進行植物水分來源判斷的方法存在一定的局限性。
[1] Cao Y L, Lu Q,Lin G H. Review and perspective on hydrogen stable isotopes technique in tracing plant water sources researches. Acta Ecologica Sinica, 2002, 22(1): 111- 117.
[2] Dawson T E, Ehleringer J R. Streamside trees that do not use stream water. Nature, 1991, 350(6316): 335- 337.
[3] Phillips S L, Ehleringer J R. Limited uptake of summer precipitation by bigtooth maple(AcergrandidentatumNutt) and Gambel′s oak (QuercusgambeliiNutt). Tree, 1995, 9(4): 214- 219.
[4] Ehleringer J R, Dawson T E. Water uptake by plants:perspectives from stable isotope composition. Plant, Cell and Environment, 1992, 15(9), 1073- 1082.
[5] Ehlerringer J R, Hall A E, Farquhar G D, Saugier B. Stable Isotopes and Plant Carbon-Water Relations. New York:Academic Press, 1993.
[6] Dawson T E. Hydraulic lift and water use by plants: Implications for water balance, performance and plant-plant interactions. Oecologia, 1993, 95(4): 565- 574.
[7] Gonfiantini R, Gratziu S, Tongiorgi E. Oxygen isotopic composition of water in leaves// Isotopes and Radiation in Soil-Plant Nutrition Studies. Vienna: International Atonatic. Energy Agency, 1965: 405- 410.
[8] Wershaw R L, Friedman I, Heller S J. Hydrogen isotope fractionation of water passing through trees // Hobson F, Speers M, eds. Advances in Organic Geochemistry. New York: Pergamon, 1966: 55- 67.
[9] Shi H, Liu S R, Zhao X G, Application of stable hydrogen and oxygen isotope in water circulationg. Jourunal of Soil and Water Conservation, 2003, 17(2): 163- 166.
[10] Duan D Y, Ouyang H. Application of stable hydrogen and oxygen isotope in analyzing plant water use sources. Ecology and Environment, 2007, 16(2): 655- 660.
[11] Li S G, Remero-Saltosh G, Tsujimura M, Sugimoto A, Sasaki L,Davaa G, Oyunbaatar D. Plant water sources in the cold semiarid ecosystem of the upper Kherlen River catchment in Mongolia: A stable isotope approach. Journal of Hydrology, 2007, 333(1): 109- 117.
[12] Zhu L, Xu X, Mao G L. Water sources of shrubs grown in the northern Ningxia Plain of China characterized by shallow groundwater table. Chinese Journal of Plant Ecology, 2012, 36(7): 618- 628.
[13] Zhou Y D, Chen S P, Song W M, Lu Q, Lin G H. Water-use strategies of two desert plants along a precipitation gradient in northwestern China. Chinese Journal of Plant Ecology, 2011, 35(8): 789- 800.
[14] Zhao L J, Xiao H L, Chen G D, Song Y X, Zhao L, Li C Z, Yang Q.A preliminary study of water sources of riparian plants in the Lower Reaches of the Heihe Basin. Acta Geoscientica Sinica, 2008, 29(6): 709- 718.
[15] Zhu Y J, Jia Z Q, Lu Q, Hao Y G, Zhang J B, Li L, Qi Y L. Water use strategy of five shrubs in Ulanbuh desert. Scientia Silvae Sinicae, 2010, 46(4): 15- 21.
[16] McCole A A, Stern L A. Seasonal water use patterns ofJuniperusasheion the Edwards Plateau, Texas, based on stable isotopes in water. Journal of Hydrology, 2007, 34(3/4): 238- 248.
[17] Yao T D, Zhu L P. The response of environmental changes on Tibetan Plateau to global changes and adaptation strategy.Advances in Earth Science, 2006, 21(5): 459- 464.
[18] Craig H. Isotopic variations in meteoric water. Science, 1961, 133(3465): 1702- 1703.
[19] Xu S G, Liu Y F, Sun W G. Research on the stable isotope for soil water vertical transporting in unsaturated zone of Zhaolong Wetland. Journal of China Hydrology, 2006, 26(5): 1- 6.
[20] Yin L, Zhao L J, Ruan Y F, Xiao H L, Cheng G D, Zhou M X, Wang F, Li C Z. Study of the replenishment sources of typical ecosystems water and dominant plant water in the lower reaches of the Heihe, China. Journal of Glaciology and Geocryology, 2012, 34(6): 1478- 1486.
[21] Wei Y F, Guo K, Chen J Q. Effect of precipitation pattern on recruitment of soil water in Kubuqi Desert, Northwestern China. Journal of Plant Ecology, 2008, 32(6): 1346- 1355.
[22] Du J H, Yan P, E Y H, Han F G. Influence of heavy rain event on soil water content ofNitrariatangutorumnebkhas in different succession periods. Bulletin of Soil and Water Conservation, 2007, 27(6): 20- 24.
[23] Zimmermann U, Ehhalt D, Muennich K O. Soil-water movement and evapotranspiration : Changes in the isotopic composition of the water. Isotopes in Hydrology, 1968, 31(10): 567- 585.
[24] Liu Z M, Wang G L, Yang Z J, Ma M Z. Exploitation and utilization of future water resources and its environment effects at Geermu-Nuomuhong region. Geography and Territorial Research, 2001, 17(3): 68- 72.
[25] Tian L D, Yao T D, Sun W Z, Tsujimura M. Stable isotope in soil water in the middle of Tibetan Plateau. Acta Pedologica Sinica, 2002, 39(3): 289- 295.
[26] Zhang Y D,Bai S B,Wang Z Q,Shen Y X. Effect of the mixed on root growth and distribution ofFraxinusmandshuricaandLarixgmelinii. Scientia Silvae Sinicae, 2001, 37(5): 16- 23.
[27] Wang Z Q, Zhang Y D, Wang Q C. Study on the root interactions betweenFraxinusmandshuricaandLarixgmelinii. Acta Phytoecologica Sinica, 2000, 24(3): 346- 350.
[28] Zeng Q, Ma J Y. Plant water sources of different habitats and its environmental indication in Heihe River Basin. Journal of Glaciology and Geocryology, 2013, 35(1): 148- 155.
[29] Macfall J S, John G A, Kramer P J. Comparative water uptake by roots of different ages in seedlings of loblolly pine(PinustaedaL.). New Phytologist, 1991, 119(4): 551- 560.
[30] Atkinson D. Plant Root Growth: An Ecological Perspective.UK: Blackwell Scientific Publications, 1991.
[31] Sun X, Yu Z. A study on root system ofNitrariaTangutorum.Journal of Desert Research, 1992, 12(4): 50- 54.
[32] Huang M. Studies on relationship between sprinkle irrigation system orEphedrain dry sandy area and distribution of root system. Journal of Ningxia Agricultural College, 2003, 24(3): 34- 36.
[33] Li G H.Ephedradistachya. Soil and Water Conservation in China, 1989, (10): 48- 48.
[34] Xu G Q, Li Y. Roots distribution of three desert shrubs and their response to precipitation under co-occurring conditions. Acta Ecologica Sinica, 2009, 29(1): 130- 137.
[35] Zhu L, Xu X, Tang J N. Study on the water sources of 4 bushes based on the18O stable isotope grew in Yinchuan plain northern area in the beginning of summer. Journal of Fujian Forestry Science and Technology, 2012, 39(1): 28- 34.
[36] Li H T. The primary study on the relation betweenCalligonumleucocladumand its environment. Journal of Xinjiang Agricultural University, 1996, 19(1): 56- 61, 72- 72.
[37] Synder K A, William D C. Water resources used by riparian trees varies among stream types on the Pedro River, Arizona.Agricultural and Forest Metrology, 2000, 105(1/3): 227- 240.
[38] Chen Y N, Li W H, Chen Y P, Xu C C, Zhang L H. Water conveyance in dried-up riverway and ecological restoration in the lower reaches of Tarim River, China. Acta Ecologica Sinica, 2007, 27(2): 538- 545.
[39] Fan Z L, Chen Y N, Li H P, Ma Y J, Alishir K,Abdimijit.Determinationof suitable ecological groundwater depth in arid areas in northwest part of China. Journal of Arid Land Resources and Environment, 2008, 22(2): 1- 4.
[40] Yang Z H, Gao Z H. Impact of precipitation and underground water level in the edge of oases on growth and decline ofNitrariatangugtorumCommunity. Chinese Journal of Applied Ecology, 2000, 11(6): 923- 926.
[41] Liu F M, Jin Y, Zhang X J. Soil water regime ofTamarixramosissimasandbreak in the desert region. Acta Botanica Boreali-Occidentalia, Sinica, 2001, 21(5): 937- 943.
[42] Liang S M, Zhang X M, Zeng F J, Zhang Z W, Qiu S K, Yan H L. Ecophysiological responses ofCalligonumarborescensLitv.to irrigation volume in the hinterland of the Taklimakan desert. Journal of Desert Research, 2010, 30(6): 1348- 1353.
參考文獻:
[1] 曹燕麗, 盧琦, 林光輝.氫穩定性同位素確定植物水源的應用與前景.生態學報, 2002, 22(1): 111- 117.
[9] 石輝, 劉世榮, 趙曉廣. 穩定性氫氧同位素在水分循環中的應用.水土保持學報, 2003, 17(2): 163- 166.
[10] 段德玉, 歐陽華. 穩定氫氧同位素在定量區分植物水分利用來源中的應用. 生態環境, 2007, 16(2): 655- 660.
[12] 朱林, 徐興, 毛桂蓮. 寧夏平原北部地下水埋深淺地區不同灌木的水分來源. 植物生態學報, 2012, 36(7): 618- 628.
[13] 周雅聃, 陳世蘋, 宋維民, 盧琦, 林光輝.不同降水條件下兩種荒漠植物的水分利用策略. 植物生態學報, 2011, 35(8): 789- 800.
[14] 趙良菊, 肖洪浪, 程國棟, 宋耀選, 趙亮, 李彩芝, 楊秋. 黑河下游河岸林植物水分來源初步研究. 地球學報, 2008, 29(6): 709- 718.
[15] 朱雅娟, 賈志清, 盧琦, 郝玉光, 張景波, 李磊, 綦艷林. 烏蘭布和沙漠5種灌木的水分利用策略. 林業科學, 2010, 46(4): 15- 21.
[17] 姚檀棟, 朱立平. 青藏高原環境變化對全球變化的響應及其適應對策. 地球科學進展, 2006, 21(5): 459- 464.
[19] 許士國, 劉盈斐, 孫萬光. 扎龍濕地包氣帶土壤水分垂直運移的穩定同位素研究. 水文, 2006, 26(5): 1- 6.
[20] 尹力, 趙良菊, 阮云峰, 肖洪浪, 程國棟, 周茅先, 王芳, 李彩芝. 黑河下游典型生態系統水分補給源及優勢植物水分來源研究. 冰川凍土, 2012, 34(6): 1478- 1486.
[21] 魏雅芬, 郭柯, 陳吉泉. 降雨格局對庫布齊沙漠土壤水分的補充效應. 植物生態學報, 2008, 32(6): 1346- 1355.
[22] 杜建會, 嚴平, 俄有浩, 韓富貴. 強降雨事件對不同演化階段白刺灌叢沙堆土壤水分的影響.水土保持通報, 2007, 27(6): 20- 24.
[24] 劉志明, 王貴玲, 楊振京, 馬明珠. 格爾木至諾木洪地區水資源開發利用及其環境效應分析.地理學與國土研究, 2001, 17(3): 68- 72.
[25] 田立德, 姚檀棟, 孫維貞, Tsujimura M. 青藏高原中部土壤水中穩定同位素變化. 土壤學報, 2002, 39(3): 289- 295.
[26] 張彥東, 白尚斌, 王政權, 沈有信. 混交條件下水曲柳落葉松根系的生長與分布. 林業科學, 2001, 37(5): 16- 23.
[27] 王政權, 張彥東, 王慶成. 水曲柳落葉松根系之間的相互作用研究. 植物生態學報,2000, 24(3): 346- 350.
[28] 曾巧, 馬劍英. 黑河流域不同生境植物水分來源及環境指示意義. 冰川凍土, 2013, 35(1):148- 155.
[31] 孫祥, 于卓. 白刺根系的研究. 中國沙漠, 1992, 12(4): 50- 54.
[32] 黃梅. 干旱多沙區麻黃噴灌灌溉制度和根系分布試驗研究. 寧夏農學院學報, 2003, 24(3): 34- 36.
[33] 李煥廣. 麻黃. 中國水土保持, 1989, (10): 48- 48.
[34] 徐貴青, 李彥. 共生條件下三種荒漠灌木的根系分布特征及其對降水的響應. 生態學報, 2009, 29(1): 130- 137.
[35] 朱林, 許興, 唐建寧. 基于18O穩定同位素的銀川平原北部4種灌木夏初水分來源的研究. 福建林業科技, 2012, 39(1): 28- 34.
[36] 李海濤. 白皮沙拐棗(Calligonumleucocladum)根系與環境關系的初步研究.新疆農業大學學報, 1996, 19(1): 56- 61, 72- 72.
[38] 陳亞寧, 李衛紅, 陳亞鵬, 徐長春, 張麗華. 新疆塔里木河下游斷流河道輸水與生態恢復.生態學報, 2007, 27(2): 538- 545.
[39] 樊自立, 陳亞寧, 李和平, 馬英杰, 艾里西爾·庫爾班, 阿布都米吉提. 中國西北干旱區生態地下水埋深適宜深度的確定. 干旱區資源與環境, 2008, 22(2): 1- 4.
[40] 楊自輝, 高志海. 荒漠綠洲邊緣降水和地下水對白刺群落消長的影響. 應用生態學報, 2000, 11(6): 923- 926.
[41] 劉發民, 金燕, 張小軍. 荒漠地區檉柳人工固沙林土壤水分動態研究. 西北植物學報, 2001, 21(5): 937- 943.
[42] 梁少民, 張希明, 曾凡江, 張仲伍, 邱士可, 閆海龍. 沙漠腹地喬木狀沙拐棗對灌水量的生理生態響應. 中國沙漠, 2010, 30(6): 1348- 1353.
Water sources of five dominant desert plant species in Nuomuhong area of Qaidam Basin
XING Xing1,CHEN Hui1,*,ZHU Jianjia1,2,CHEN Tongtong1
1CollegeofResourcesandEnvironmentSciences,HebeiNormalUniversity,HebeiKeyLaboratoryofEnvironmentalChangeandEcologicalConstruction,Shijiazhuang050024,China2InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China
Water is one of the essential substances to maintain the plant system. Water resource selection is an important selection process in the botany cycle. Despite precipitation is the ultimate water source for all plants, factors like evaporation, difference in altitude, geological aquifer features, differences in the soil water characteristics may all influence both values of stable hydrogen and oxygen isotope (δD,δ18O). Where the plant extracts water is supposed to be revealed by comparing the waterδ18O、δD values from stem xylems with that from the available water sources. Qaidam Basin is located in the northeast of Tibetan Plateau with its typical desert ecosystem being significantly sensitive to global change. Nevertheless, there is no information found on how its plant system distributes their water exploration among the available water sources in that area. How the river water and precipitation influence the plant water sources is unknown. The objective of this study is to determine the proportions of different water sources drawn by selected dominant plant species, examine the consistence of this proportion for different plant species during the growth season, investigate the water competition conditions among the selected dominant species. Five local dominant plant species (Ceratoideslatens,Nitrariatangutorum,Ephedrasinica,Tamarixramosissima,Calligonummongolicum) were selected to test the stem waterδ18O andδD using Picarro L- 2130i and Finnegan MAT-253.δ18O andδD was also tested for different available water resources including groundwater, river water, soil water (profiled as 0—10 cm,10—30 cm,30—50 cm,50—70 cm,70—90 cm) and effective precipitation at same time, as sampled every 15 days during June to September in 2010 in Nuomuhong Area. Meanwhile Decagon′s Em50Em50R was used to measure soil water content. The Multi-source mixed linear model (IsoSource) was used to analyze the contribution of different water sources to these desert plants. The local meteoric water line (LMWL) equation wasy=7.019x-3.217(R2=0.970,P<0.001), which indicated the warm and dry climate features of the study area. The result ofδ18O andδD analysis showed that the selected plant species mainly used soil water in the whole growth season. The groundwater was the second major water sources.Ceratoideslatensmostly used soil water in 10—50 cm.NitrariatangutorumandTamarixramosissimaabsorbed most soil water from 50 to 70 cm. Soil water competition existed in 50—70 cm between these two species.EphedrasinicaandCalligonummongolicumused soil water from each layer more evenly. So their water competition exists in the whole profile.Nitrariatangutorum,Ephedrasinica,TamarixramosissimaandCalligonummongolicumchanged the absorbing proportion of different water sources during growth season. Groundwater utilization is increased gradually in the test season as the plant grows, which indicated that groundwater is an important water source to the desert vegetation. Thus maintaining the groundwater tables in arid and semi-arid area is important for desert plant system. For water scarce region, desert shrub root phenotype or spatial structure is an important aspect for root functioning. Mostly plant root system distributed throughout the whole soil profile. It does not always coincide with the water abstracting profile. So the way of judging the plant water sources only based on plant root profile has its limitation.
Qaidam Basin; Nuomuhong area; desert plants;water sources;stable hydrogen and oxygen isotope
國家自然科學基金項目(40971118);河北師范大學碩士研究生科研基金項目(201202021); 河北省自然地理學省級重點學科資助項目
2013- 10- 09;
2014- 05- 29
10.5846/stxb201310092427
*通訊作者Corresponding author.E-mail: chenhui@igsnrr.ac.cn
邢星,陳輝,朱建佳,陳同同.柴達木盆地諾木洪地區5種優勢荒漠植物水分來源.生態學報,2014,34(21):6277- 6286.
Xing X,Chen H,Zhu J J,Chen T T.Water sources of five dominant desert plant species in Nuomuhong area of Qaidam Basin.Acta Ecologica Sinica,2014,34(21):6277- 6286.
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
小讀者(2021年2期)2021-03-29 05:03:48
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
