鎘處理下平邑甜茶DNA的甲基化分析
2014-07-07 15:38:55王利李鵬飛楊洪強(qiáng)謝會(huì)成李龍謝永波
王利,李鵬飛,楊洪強(qiáng),謝會(huì)成,李龍,謝永波
1.山東農(nóng)業(yè)大學(xué)農(nóng)業(yè)生態(tài)與環(huán)境重點(diǎn)實(shí)驗(yàn)室,山東泰安271018 2.山東省東營(yíng)市林業(yè)局,山東東營(yíng)257091 3.山東農(nóng)業(yè)大學(xué)農(nóng)業(yè)資源利用博士后流動(dòng)站,山東泰安271018
鎘處理下平邑甜茶DNA的甲基化分析
王利1,3,李鵬飛2,楊洪強(qiáng)3*,謝會(huì)成1,李龍1,謝永波1
1.山東農(nóng)業(yè)大學(xué)農(nóng)業(yè)生態(tài)與環(huán)境重點(diǎn)實(shí)驗(yàn)室,山東泰安271018 2.山東省東營(yíng)市林業(yè)局,山東東營(yíng)257091 3.山東農(nóng)業(yè)大學(xué)農(nóng)業(yè)資源利用博士后流動(dòng)站,山東泰安271018
采用甲基化敏感擴(kuò)增多態(tài)性(Methylation sensitive amplified polymorphism,MSAP)技術(shù),對(duì)鎘處理的平邑甜茶葉片DNA進(jìn)行甲基化位點(diǎn)分析,研究鎘對(duì)其甲基化多態(tài)性的影響。結(jié)果表明:在0(CK)、2.5 mg·L-1、5 mg·L-1、10 mg·L-1鎘處理下,9對(duì)MSAP引物擴(kuò)增,甲基化出現(xiàn)三種不同模式即未甲基化A、半甲基化B和全甲基化C,共擴(kuò)增出3150條譜帶,單對(duì)引物最多擴(kuò)增656條,最少為610條。5 mg.L-1鎘處理的平邑甜茶甲基化較高,比例達(dá)到60.1%;10 mg.L-1鎘處理的平邑甜茶甲基化較低,比例達(dá)到57.6%;與對(duì)照相比,鎘處理的平邑甜茶甲基化水平降低,可能鎘抑制了其基因的表達(dá)。
平邑甜茶;甲基化;甲基化敏感擴(kuò)增多態(tài)性
DNA甲基化是指生物體在DNA甲基轉(zhuǎn)移酶(DNA methyltransferase)的催化下,將甲基供體S-腺苷甲硫氨酸(SAM)轉(zhuǎn)移到特定堿基上的過程。在動(dòng)物和植物中普遍存在DNA甲基化,它與植物的生長(zhǎng)發(fā)育、進(jìn)化有密切的關(guān)系。植物DNA甲基化水平受發(fā)育時(shí)期[1-2]、不同發(fā)育組織[3-4]、不同種質(zhì)資源[5-6]、DNA損傷與修復(fù)[7]及不同脅迫等的影響[8]。不同脅迫對(duì)植物甲基化影響存在差異,低溫使水稻甲基化差異片段CIDM 7的去甲基化表達(dá)增強(qiáng)[9],鹽脅迫使小麥葉片發(fā)生超甲基化[10],水分脅迫導(dǎo)致水稻葉片根DNA甲基化平均水平明顯增加,其中根部增加幅度尤為明顯[11],生物脅迫使小麥甲基化增加[12],5~100 mg/L鉻處理小麥幼苗的研究表明100 mg/L鉻濃度導(dǎo)致3 d齡小麥幼苗根系DNA胞嘧啶甲基化水平降低,其他處理濃度卻增加。重金屬鎘引起擬南芥、蘿卜甲基化程度的提高[13-14],但是,關(guān)于鎘究竟如何影響平邑甜茶甲基化還未見闡述,為此,以平邑甜茶為材料,利用鎘對(duì)其處理,探明鎘處理下平邑甜茶DNA甲基化的變化。
1 材料與方法
1.1 材料與處理
取大小一致的平邑甜茶種子,經(jīng)消毒層積1個(gè)多月后播在育苗容器中。當(dāng)幼苗第2片真葉剛出現(xiàn)時(shí),選生長(zhǎng)相近的植株移至1/2 Hoagland營(yíng)養(yǎng)液中,每3 d換1次培養(yǎng)液,培養(yǎng)至出現(xiàn)第6片葉后,在營(yíng)養(yǎng)液中加入氯化鎘,使其終濃度為0(CK)、0.5 mg·L-1、2.5 mg·L-1、5 mg·L-1、10 mg·L-1,每處理3株,重復(fù)3次,30 d后,提取葉片DNA,利用甲基化敏感擴(kuò)增多態(tài)性(Methylation sensitive amplified polymorphism,MSAP)檢測(cè)葉片DNA甲基化的多態(tài)性。
1.2 引物

1.3 引物對(duì)的篩選
從5個(gè)供試樣品中選出2個(gè)具有代表性的樣品利用銀染技術(shù)對(duì)63對(duì)引物進(jìn)行篩選,篩選出E-AAC/H-TCG、E-AAC/H-TGT、E-AAC/H-TCC、E-AAG/H-TCG、E-AAG/H-TGC、E-AAG/H-TCC、E-ACA/H-TCG、E-ACA/H-TGT、E-ACA/H-TCC9對(duì)引物對(duì)供試樣品進(jìn)行擴(kuò)增。從中篩選出檢測(cè)位點(diǎn)較多,分布均勻,清晰可辨且多態(tài)位點(diǎn)百分比較高的引物對(duì)各個(gè)樣品進(jìn)行MSAP分析。
1.4MSAP分析
據(jù)北京鼎國(guó)生物技術(shù)有限責(zé)任公司FISH-MSAP試劑盒說明,先進(jìn)行EcoRI/HpaⅡ、MspⅠ酶切與T4連接酶連接,取2 μL酶切連接液做模板,用帶有1個(gè)選擇性堿基的引物進(jìn)行預(yù)擴(kuò)增,再將預(yù)擴(kuò)增液按1:15比例稀釋,以2 μL預(yù)擴(kuò)增稀釋液為模板,用帶有3個(gè)選擇性堿基的引物對(duì)進(jìn)行選擇性擴(kuò)增。變性產(chǎn)物(含有內(nèi)標(biāo)GeneScan-500)上樣于4%變性聚炳烯酰胺凝膠,恒定功率50 W、最大電壓3000 V,進(jìn)行電泳2.4 h,利用ABI 377測(cè)序儀對(duì)其進(jìn)行檢測(cè)。
圖1 DNA瓊脂糖電泳結(jié)果Fig.1 Result of DNAagarose electrophoresis
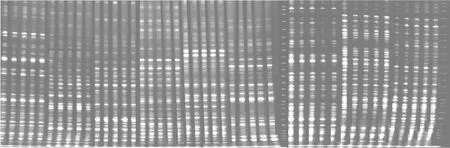
圖2 EcoR I/Hpa II酶切擴(kuò)增結(jié)果Fig.2 Results of EcoR I/Hpa II enzyme amplification

圖3 EcoR I/Msp I酶切擴(kuò)增結(jié)果Fig.3 Results of EcoR I/Msp I enzyme amplification
1.5 統(tǒng)計(jì)分析
利用GeneScan3.1軟件對(duì)圖像進(jìn)行處理,構(gòu)建0、1數(shù)學(xué)矩陣。通過比較電泳譜帶的差異,可以推測(cè)出CCGG的甲基化情況[15]。
2 結(jié)果與分析
2.1 基因組的質(zhì)量
本實(shí)驗(yàn)提取平邑甜茶DNA經(jīng)瓊脂糖電泳檢測(cè)(如圖1),主帶清晰,片段大小在23 kb左右,無降解。這說明該法提取的銀杏DNA質(zhì)量較高,符合MSAP技術(shù)要求。
2.2 鎘處理下平邑甜茶DNA甲基化模式分析
平邑甜茶DNA經(jīng)過EcoR I/Hpa II、EcoR I/Msp I酶切、連接、預(yù)選擴(kuò)增、聚丙烯酰胺凝膠電泳后得到MSAP圖譜(圖2,圖3),譜帶清晰。圖中譜帶表明其甲基化主要有三種不同模式:模式A是指EH、EM均有帶,代表非甲基化位點(diǎn),說明未發(fā)生甲基化(或單鏈酶切位點(diǎn)內(nèi)側(cè)甲基化);模式B是指EH有帶、EM無帶,代表半甲基化位點(diǎn),說明該位點(diǎn)發(fā)生了單鏈酶切位點(diǎn)外側(cè)的胞嘧啶甲基化;模式C是指EH無帶、EM有帶,代表全甲基化位點(diǎn),說明該位點(diǎn)發(fā)生了雙鏈酶切位點(diǎn)內(nèi)側(cè)的胞嘧啶甲基化。
2.3 鎘處理下平邑甜茶DNA甲基化水平分析
MSAP圖譜中每個(gè)泳道的每一條帶代表一個(gè)酶切位點(diǎn),根據(jù)有帶記為1,無帶記為0的方法分析統(tǒng)計(jì)其電泳譜帶。9對(duì)引物共擴(kuò)增出3150條譜帶,單對(duì)引物最多擴(kuò)增656條,最少為610條。在鎘處理中,5 mg.L-1鎘處理的平邑甜茶甲基化條帶最多,共385條,甲基化達(dá)到60.1%;2.5 mg.L-1鎘處理的平邑甜茶甲基化條帶最少,共355條,甲基化達(dá)到57.8%。在擴(kuò)增位點(diǎn)中,檢測(cè)到甲基化位點(diǎn)1871個(gè),甲基化比例為59.4%;其中,半甲基化位點(diǎn)941個(gè),比例為29.9%,全甲基化位點(diǎn)為930個(gè),比例為29.5%,半甲基化與全甲基化位點(diǎn)比例接近,說明鎘處理對(duì)單鏈酶切位點(diǎn)外側(cè)的胞嘧啶甲基化和雙鏈酶切位點(diǎn)內(nèi)側(cè)的胞嘧啶甲基化的影響差異不大。另外,模式B中對(duì)照擴(kuò)增譜帶最多206條,10 mg.L-1鎘處理最少167條,總體呈現(xiàn)減少的趨勢(shì);模式C中對(duì)照擴(kuò)增譜帶最多197條,2.5 mg.L-1最少168條,出現(xiàn)先減少后增加的變化。與對(duì)照相比,鎘處理使平邑甜茶甲基化位點(diǎn)數(shù)量總體出現(xiàn)減少的趨勢(shì)(表1)。
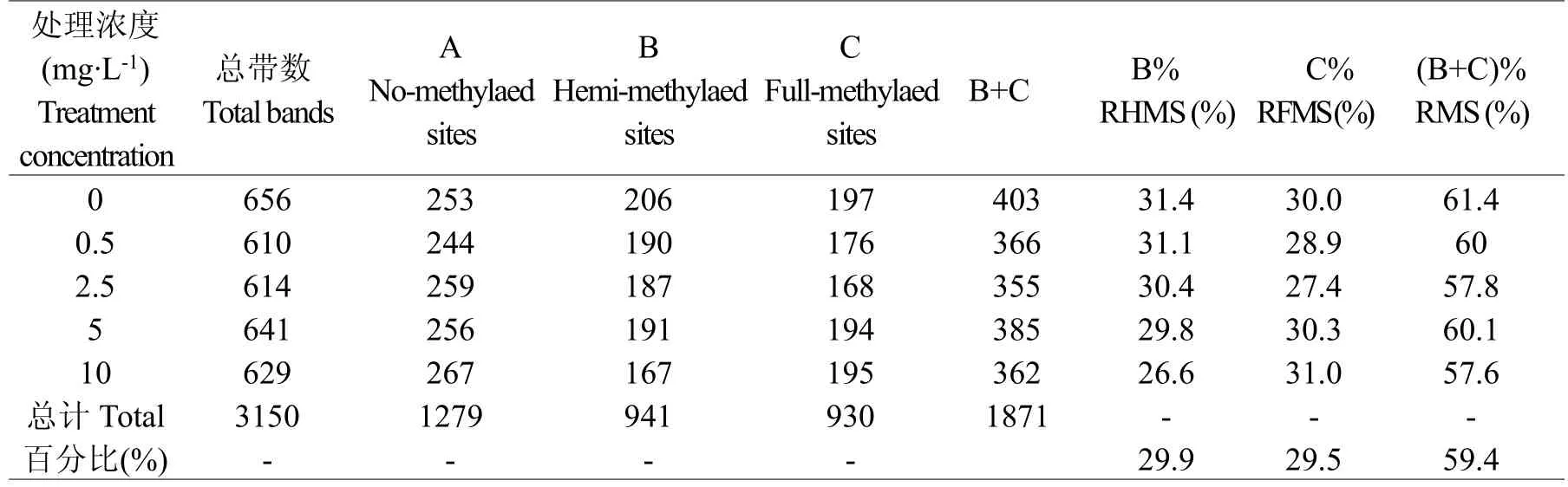
表1 鎘處理下平邑甜茶葉片甲基化多態(tài)性分析Table1Analysis of methylation polymorphism on Malus hupehensis leaves under Cd treatment
3 討論
表觀遺傳學(xué)是指在DNA序列沒有發(fā)生改變的情況下,遺傳信息在基因功能上發(fā)生可遺傳的變化,導(dǎo)致表型變異[16],近10年來逐漸受到廣泛重視,其研究成為當(dāng)今生命科學(xué)研究的熱點(diǎn)領(lǐng)域。植物DNA甲基化是在甲基轉(zhuǎn)移酶的作用下,把DNA的一個(gè)甲基添加在DNA分子的堿基上,通常加在胞嘧啶上,形成甲基胞嘧啶[16]。在植物正常發(fā)育過程中,DNA甲基化能夠調(diào)控生長(zhǎng)發(fā)育等過程,對(duì)植物生長(zhǎng)發(fā)育具有重要的調(diào)節(jié)作用。DNA甲基化的水平模式發(fā)生改變,就會(huì)影響到植物的生長(zhǎng)發(fā)育,使其生長(zhǎng)發(fā)育異常。李海林等研究巴西橡膠樹DNA甲基化模式表明,其存在非甲基化、半甲基化和全甲基化三種模式[17],本實(shí)驗(yàn)研究也得到了上述三種不同的甲基化模式。不同植物甲基化多態(tài)性水平不一[14,17]。隨著鎘處理濃度的增加,擬南芥甲基化程度增加[14]。本研究結(jié)果表明,鎘處理影響平邑甜茶葉片DNA的甲基化水平,使半甲基化和甲基化多態(tài)性發(fā)生了改變,它們的多態(tài)性比例為26.6~31.1%,尤其是使平邑甜茶葉片DNA半甲基化多態(tài)性出現(xiàn)減小的趨勢(shì),這與馬子成等研究結(jié)果相反,甲基化多態(tài)性出現(xiàn)減小可能是鎘處理抑制了基因的表達(dá)。
4 結(jié)論
(1)在鎘處理中,甲基化出現(xiàn)三種不同模式即未甲基化A(EcoR I/Hpa II、EcoR I/Msp I酶切擴(kuò)增均有帶)、半甲基化B(EcoR I/Hpa II酶切擴(kuò)增有帶、EcoR I/Msp I酶切擴(kuò)增無帶)
(2)在鎘處理中,5 mg.L-1鎘處理的平邑甜茶甲基化較高,比例達(dá)到60.1%;10 mg.L-1鎘處理的平邑甜茶甲基化較低,比例達(dá)到57.6%;與對(duì)照相比,鎘處理的平邑甜茶甲基化水平降低,可能鎘抑制了其基因的表達(dá)和全甲基化C(EcoR I/Hpa II酶切擴(kuò)增無帶、EcoR I/Msp I酶切擴(kuò)增有帶)。
[1]Finnegan EJ,Genger RK,Peacock W J.et al.DNA methylation in plant.Ann.Rev[J].Plant Mol.Biol.,1998,49:233-247
[2]Finnegan EJ,Peacock WJ,Dennis ES.DNA methylation,a key regulator of plant development and other proc esses[J].Current Opinion in Genetics and Development,2000,10:217-223
[3]Lu CM,J iang GM,Wang B S,et al.Photosystem II photochemistry and photosynthetic pigment composition in salt-adapted halophyte Artimisia anethfolia grown under outdoor conditions[J].J Plant Physiol,2003,160:403-408
[4]儀治本,孫毅,牛天堂,等.高梁基因組DNA胞嘧啶甲基化在雜交種和親本間差異研究[J].作物學(xué)報(bào),2005,31(9):1138-1143
[5]Guo M,Davis D,Birchler J A.Dosage effects on gene expression in a maize ploidy series[J].Genetics, 1996,142:1349-1355
[6]Keyte AL,Percifield R,Liu B,et al.Infraspecific DNA methylation polymorphism in cotton(Gossypium hirsutum L.). J Heredity,2006,97(5):444-450
[7]武立鵬,朱衛(wèi)國(guó).DNA甲基化的生物學(xué)應(yīng)用急檢測(cè)方法進(jìn)展[J].中華檢驗(yàn)醫(yī)學(xué)雜志,2004,27(7):468-474
[8]范建成,劉寶,王雋媛,等.萘脅迫對(duì)水稻基因組DNA甲基化模式及水平的影響[J].環(huán)境科學(xué),2010,31(3):793-800
[9]華揚(yáng),陳學(xué)峰,熊建華,等.水稻冷脅迫誘導(dǎo)的甲基化差異片段CIDM7的分離和分析[J].遺傳,2005,27(4):595-600
[10]鐘蘭,王建波.DNA超甲基化在小麥耐鹽脅迫中的作用[J].武漢植物學(xué)研究,2007,25(1):102-104
[11]潘雅嬌,傅彬英,王迪,等.水稻干旱脅迫誘導(dǎo)DNA甲基化時(shí)空變化特征分析[J].中國(guó)農(nóng)業(yè)科學(xué),2009,42(9):3009-3018
[12]付勝杰,王暉,馮麗娜,等.葉銹菌脅迫下的小麥基因組MSAP分析[J].遺傳,2009,31(3):297-304
[13]楊金蘭,柳李旺,龔義勤,等.鎘脅迫下蘿卜基因組DNA甲基化敏感擴(kuò)增多態(tài)性分析[J].植物生理與分子生物學(xué)學(xué)報(bào),2007,33(3):219-226
[14]王子成,馬洪霞,何艷霞.重金屬鎘對(duì)擬南芥DNA甲基化的影響[J].植物生理學(xué)通訊,2009,45(2):115-118
[15]陸光遠(yuǎn),伍曉明,陳碧云,等.油菜種子萌發(fā)過程中DNA甲基化的MSAP分析[J].科學(xué)通報(bào),2005,50(24):2750-2756
[16]薛京倫,旺旭,吳超群,等.表觀遺傳學(xué)-原理、技術(shù)與實(shí)踐[M].上海:上海科學(xué)技術(shù)出版社,2006
[17]李海林,吳春太,李衛(wèi)國(guó).巴西橡膠樹DNA甲基化的MSAP分析[J].分子植物育種,2011,9(1):89-73
Analysis of Methylation Polymorphism on Malus hupehensis Leaves under Cd Treatment
WANG Li1,3,LI Peng-fei2,YANG Hong-qiang3*,XIE Hui-cheng1,LI Long1, XIE Yong-bo1
1.Key Laboratory of Agricultural Ecology and Environment,Shandong Agricultural University,Taian 271018,China 2.Forestry Station,Dongying Municipal Forestry Bureau,Dongying 257091,China 3.Post-doctoral Mobile Station of Agricultural Resource Utilization,Shandong Agricultural University,Taian 271018,China
Methylation site and methylation polymorphism were analyzed on DNA of Malus hupehensis Rehd.under Cd treatment with methylation sensitive amplified polymorphism(MSAP).The results showed that amplification of 9 primer combinations formed three different modes and produced 3150 bands.The maximum number of bands was 656 and the minum number of bands was 610 per primer combination under different concengtration Cd treatment of 0(CK),2.5 mg·L-1, 5 mg·L-1,10 mg·L-1.The highest rate of methylation polymorphism appeared under 5 mg·L-1Cd treatment with value of 60.1%,the lowest rate of methylation polymorphism appeared under 10 mg·L-1Cd treatment with value of 57.6%.The level of methylation polymorphism on Malus hupehensis Rehd under Cd treatment reduced compared with control sample.Its reason might be Cd incurred the expression of gene.
Malus hupehensis Rehd.;methylation;methylation sensitive amplified polymorphism
F812.4
A
1000-2324(2014)03-0372-05
2013-03-06
2013-03-11
山東農(nóng)業(yè)大學(xué)博士后課題,蘋果優(yōu)質(zhì)高效生產(chǎn)關(guān)鍵技術(shù)研究與示范(2014BAD16B02)
王利(1968-),男,山東高密人,博士,研究方向?yàn)榱帜具z傳與生理.
*通訊作者:Author for correspondence.E–mail:labft@sdau.edu.cn
 山東農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版)2014年3期
山東農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版)2014年3期
- 山東農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版)的其它文章
- 基于SVM的稿件質(zhì)量評(píng)價(jià)系統(tǒng)
- 旅游信息科學(xué)化管理及智能化評(píng)價(jià)研究
- 山東省濟(jì)南西南山區(qū)農(nóng)業(yè)生態(tài)園能值分析及可持續(xù)性評(píng)價(jià)
- 區(qū)域模糊綜合評(píng)價(jià)中的空間隸屬度尺度轉(zhuǎn)換分析
- 基于因子分析法對(duì)滬深農(nóng)業(yè)類上市公司財(cái)務(wù)績(jī)效實(shí)證分析
- 改良Carrousel氧化溝工藝處理礦區(qū)污水效果研究