油茶脂肪酸代謝途徑中關鍵酶基因調控油脂合成的規律研究
2014-06-11 00:46:02曾艷玲譚曉風張黨權陳鴻鵬曾曉峰許淑嫻
中國糧油學報 2014年2期
曾艷玲 譚曉風 張黨權 陳鴻鵬 曾曉峰 朱 勇 許淑嫻 陳 力
(經濟林育種與栽培國家林業局重點實驗室 中南林業科技大學林學院1,長沙 410004)(國家林業局桉樹研究開發中心2,湛江 524022)(中南林業科技大學食品科學與工程學院3,長沙 410004)
油茶是我國特色食用油料樹種,其產物茶油不僅不飽和脂肪酸質量分數高達80%,而且富含十六烷酰胺和角鯊烯等具保健功能的生物活性物質[1],長期食用可降血脂血壓,預防心腦血管疾病[2]。茶油之所以品質優良,主要是由于在茶油形成過程中油脂代謝途徑各調控酶起著關鍵作用。酰基載體蛋白(ACP)是脂肪酸合成中的關鍵蛋白質之一,位于脂肪酸合成酶系的中央,作為脂酰基的載體將脂酰基從一個酶反應轉移到另一個酶反應[3-4],因此ACP基因是調控飽和脂肪酸與不飽和脂肪酸比例的必要基因;硬脂酰-ACP 脫飽和酶(SAD)基因調控硬脂酸脫氫形成油酸,因此,直接決定了不飽和脂肪酸的總含量以及飽和脂肪酸與不飽和脂肪酸的比例[5-6];油酸脫氫形成亞油酸和亞麻酸則由油酸脫氫酶(FAD)基因家族調控,其中FAD2調控油酸的第12和13位碳原子之間形成雙鍵,生成亞油酸,因此FAD2決定了油酸和亞油酸的比例[7-8]。這幾個基因的表達調控作用主導著茶油不飽和脂肪酸含量的多少及品質的差異,因此本試驗擬通過研究油茶種仁中ACP、SAD和FAD2基因的表達規律,結合相應發育時期油茶種仁含油率及脂肪酸成分變化,揭示油脂合成關鍵酶基因表達對油脂合成的影響。
1 材料與方法
1.1 材料與主要試劑
油茶籽:湖南省長沙市望城縣東城鎮油茶基地,國審品種 “華碩”和普通油茶“望城1號”,取材時間為5月5日、6月5日、7月4日、8月4日、8月15日、8月25日、9月4日、9月11日、9月19日、9月26日、10月3日、10月10日、10月24日。
石油醚(30~60 ℃)、正庚烷(色譜純)、甲醇、氫氧化鉀、無水硫酸鈉:國藥集團化學試劑有限公司;RNA抽提試劑盒(PureLinkTMRNA Mini Kit):美國Invitrogen公司;熒光定量試劑盒(QuantiFast SYBR Green PCR Kit):德國QIAGEN公司;逆轉錄試劑盒(PrimeScript RT reagent Kit With gDNA Eraser):日本TaKaRa公司,所有RNA提取所用溶液均用滅菌的DEPC水配制。 所用引物均由北京華大生物技術有限公司合成。
1.2 主要儀器與設備
FOSS SCINO ST310油脂萃取系統:諾斯高斯分析儀器蘇州有限公司;G6890A氣相色譜儀(FID 檢測器):美國安捷倫科技公司;實時熒光PCR儀(CFX96)、核酸濃度測定儀:伯樂生命醫學產品(上海)有限公司;高速冷凍臺式離心機:美國貝克曼庫爾特有限公司;空氣加熱器:英國TECHNE公司。
1.3 試驗方法
1.3.1 油脂提取
參照國標GB /T 14772—2008 索氏抽提法。
1.3.2 脂肪酸成分測定
脂肪酸成分測定釆用堿式甲酯化法測定[9]。氣相色譜分析30 m×0.25 μm×0.25 mm FFAP毛細管柱;色譜柱溫度:60 ℃→180 ℃(25 ℃/min,停留1 min) →210 ℃(3 ℃/min,停留 1 min) →212 ℃(0.3 ℃/min,停留1 min) →240 ℃(8 ℃/min,停留 2 min),進樣口溫度: 240 ℃,檢測器溫度:240 ℃,分流比為1∶50,載氣流流速:N21.23 mL/min,壓縮空氣 400 mL/min,H240 mL/min,進樣量為 1 μL。
1.3.3 實時熒光定量PCR檢測
油茶種仁RNA提取:PureLinkTMRNA Mini Kit說明書;RNA逆轉錄cDNA:PrimeScript RT Rreagent Kit With gDNA Eraser 說明書。
實時熒光定量PCR引物分別為:qACPF:5’-ATTCAAGCAAAACCAGGCG-3’/ qACPR:5’-CACACGAAATCCGAAAACG-3’,qSADF:5’-GTTCAAGTAACGCACTCCAT-3’/ qSADR:5’-TTGCCAACATTTCTCCACAG-3’,q FAD2F: 5’-CCCAGCAACCAAACATGAAC-3’/ q FAD2R: 5’-GAATGAGCGGAGGAGAGAAC-3’;反應體系為:2×QuantiFast SYBR Green PCR Master Mix 12.5 μL,10 μmol/L引物對各0.5 μL,模板cDNA 1 μL,無菌純水補足25 μL;循環條件為95 ℃預變性 5 min; 95 ℃變性10 s,55 ℃退火30 s,共40循環。
2 結果與分析
2.1 種仁含油率和脂肪酸成分變化規律
根據所測種仁含油率結果分析(圖1),“華碩”和“望城1號”含油率均處于逐漸上升趨勢,8月中旬含油率僅有2%~4%,到10月下旬含油率上升至50%~60%。比較“華碩”和“望城1號”兩者差異,在各個果實發育時期,“望城1號”種仁含油率均高于“華碩”。采用軟件SPSS 17.0進行配對樣本T檢驗分析兩者差異,結果顯示為差異極顯著。
根據脂肪酸成分分析(圖2),8月至10月“華碩”棕櫚酸含量均處于逐漸遞減狀態,由 8月15日15.16%降至10月24日5.58%,油酸含量則處于逐漸遞增狀態,由8月15日53.53%增至10月24日88.07%,亞油酸含量處于逐漸遞減狀態,由8月15日28.33%減至10月24日4.01%,說明油酸積累速度快于油酸轉化亞油酸的速度,所以在油酸含量逐月遞增的情況下,亞油酸含量反而遞減;8月至10月“望城1號”棕櫚酸含量也均處于逐漸遞減狀態,由 8月15日13.88%降至10月24日7.31%,油酸含量也處于逐漸遞增狀態,由8月15日59.14%增至10月24日86.33%,亞油酸含量處于逐漸遞減狀態,由8月15日24.09%減至10月24日4.03%,變化規律與“華碩”基本相同,但是兩者對比發現“望城1號”各成分變化幅度均比“華碩’要小。
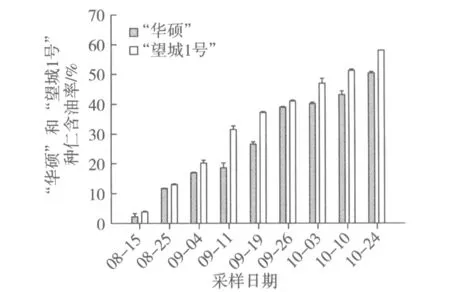
圖1 油茶種仁不同發育時期含油率的變化
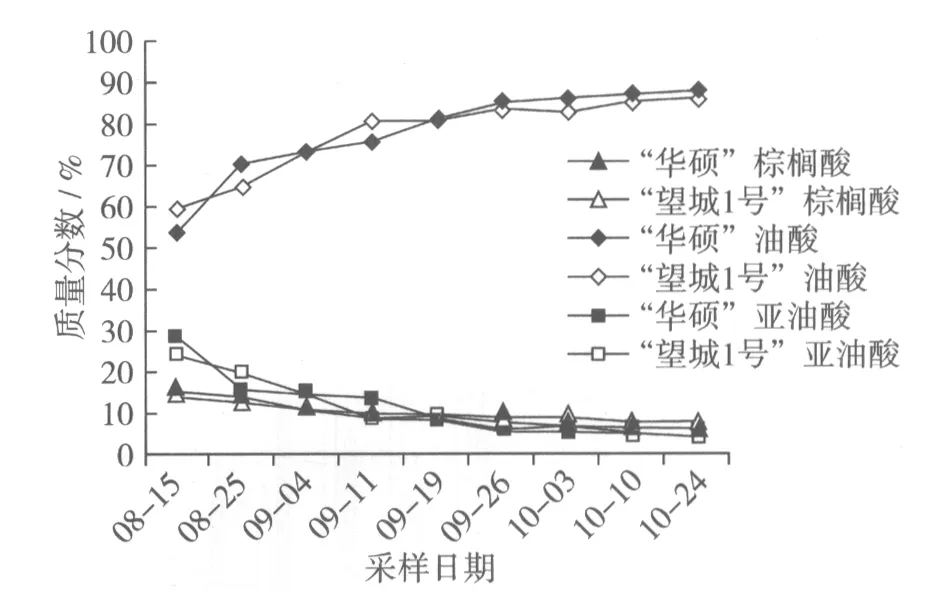
圖2 油茶種仁不同發育時期棕櫚酸、油酸和亞油酸的變化
2.2 調控脂肪酸代謝關鍵酶時空表達規律
本試驗選取穩定表達的油茶甘油醛-3-磷酸脫氫酶基因(GAPDH)為內參基因[10],以 5月5日、6月5日、7月4日、8月4日、9月4日、9月11日、9月26日和10月24日采集的油茶種仁為材料,分析油茶酰基載體蛋白基因(CoACP)、油茶硬脂酰-ACP 脫飽和酶基因(CoSAD)和油茶油酸脫飽和酶基因(CoFAD2)的相對表達量(圖3~圖5)。
根據圖3結果分析, 5月至10月期間,“華碩”CoACP相對表達量整體趨勢為先上調后下調。7月至8月CoACP表達量迅猛增加,出現第一個表達高峰期,9月初有稍微回落,9月中旬重新上調,至9月下旬達到最大值,10月開始下調。這一結果與8月中旬棕櫚酸含量是8月至9月期間最高值15.16%基本吻合(圖2)。脂肪酸成分分析結果顯示棕櫚酸含量一直穩步下降,熒光定量結果顯示“望城1號”CoACP相對表達量自8月后一直下調,這與棕櫚酸含量下降規律一致,但是“華碩”熒光定量結果卻顯示9月初CoACP相對表達量有些許下調后9月中下旬才重新上調,這可能是因為脂肪酸合成酶是一個多酶復合體,在油茶種仁不同發育時期,發揮主效作用的單酶不同所致。
根據圖4結果分析,5月至10月期間,“華碩”CoSAD相對表達量呈現先上調后下調的趨勢。7月至8月CoSAD表達量迅猛增加,9月下旬達到最大值,10月開始下調。這與脂肪酸成分測定8月下旬油酸質量分數70.25%增至9月下旬85.08%后,油酸含量平穩增長的規律基本一致(圖2)。“望城1號”CoSAD相對表達量也是呈現先上調后下調的趨勢。但是其表達高峰期是在8月,比“華碩”早2個月,隨后下調表達。這可能是 “望城1號”的油酸含量增幅比“華碩”要小的原因之一。
根據圖5結果分析,5月至10月期間,“華碩”CoFAD2相對表達量呈現先上調后下調的趨勢。7月至8月CoFAD2表達量迅猛增加,9月初表達量與8月表達量基本持平,9月中旬些許有些回落后,9月下旬達到最大值,10月開始下調。這與脂肪酸成分測定8月下旬亞油酸質量分數15.27%減至9月下旬5.51%后,亞油酸含量平緩減少的規律有所不符(圖2)。這可能是因為CoFAD2是FAD基因家族[11]的成員之一,雖然是主效調控亞油酸合成的基因,但是還是受其他家族成員的調控影響。“望城1號”CoFAD2熒光定量結果則與脂肪酸成分中亞油酸含量下降的規律一致。
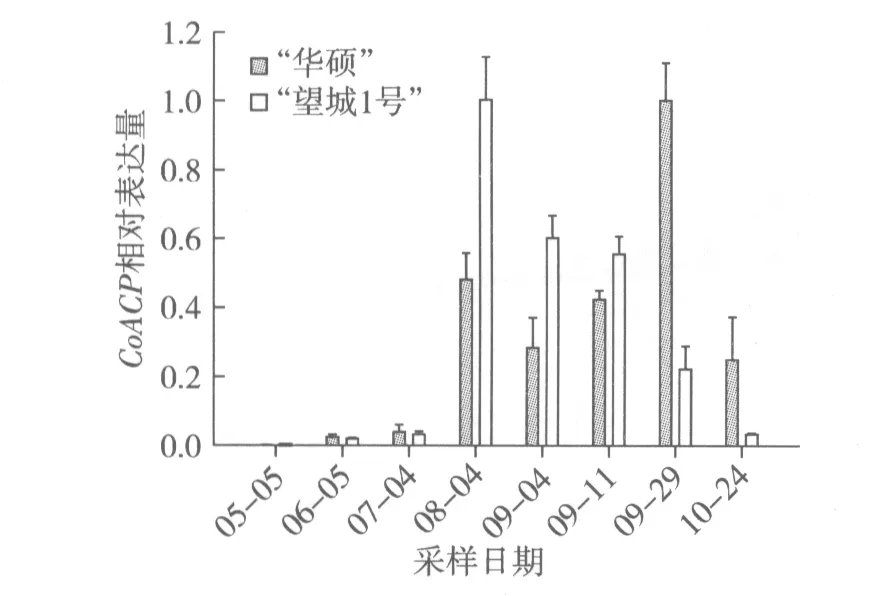
圖3 油茶種仁不同發育時期CoACP相對表達量的變化
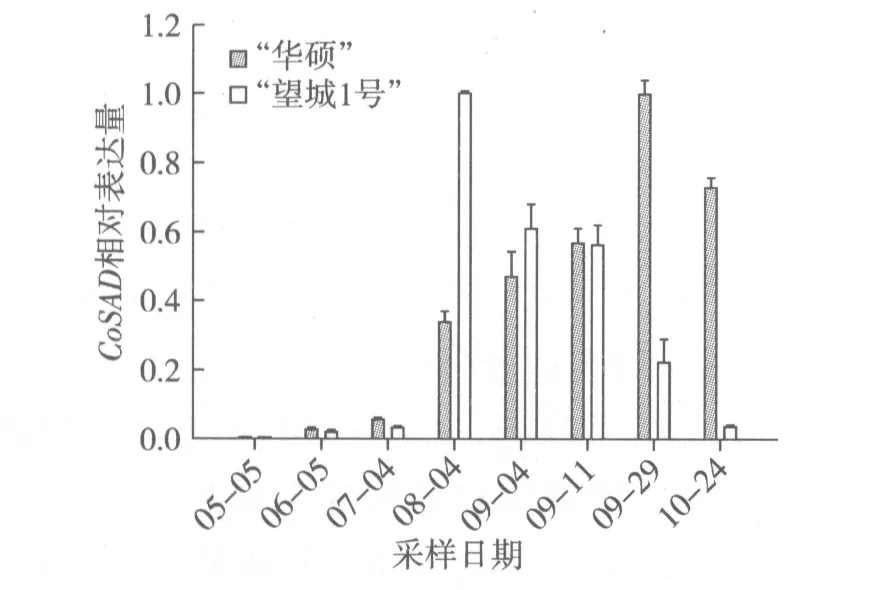
圖4 油茶種仁不同發育時期CoSAD相對表達量的變化
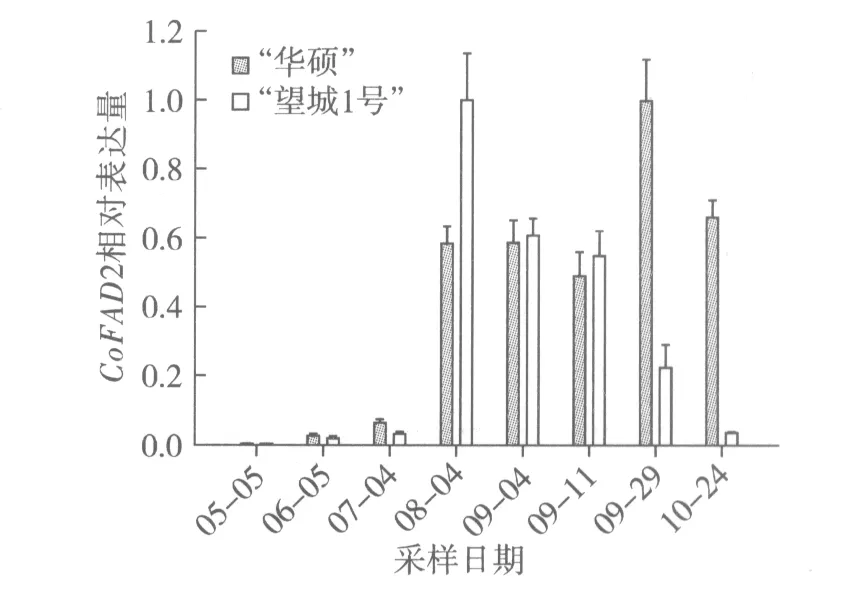
圖5 油茶種仁不同發育時期CoFAD2相對表達量的變化
圖6 不同發育時期油茶果實
3 討論與結論
生物體在發育過程中沒有永久關閉的基因。油茶果實發育過程中,隨著不同相關基因的表達,表達豐度的差異,其含油率及脂肪酸各成分含量也隨之變化。對比“華碩”和“望城1號”同時期種仁含油率,“望城1號”均高于“華碩”,但增長速率相當,說明“望城1號”較“華碩”先進入油脂合成時期,這可能與不同油茶品種果實發育周期相關。“華碩”果實發育期明顯要長于“望城1號”(圖6),“望城1號”在10月3日采摘時就已經有輕微裂果,10月24日完全成熟,而此時“華碩”還沒有裂果成熟的跡象。根據脂肪酸成分分析,雖然10月24日的“華碩”種仁含油率比“望城1號”低,但是油酸及不飽和脂肪酸總含量高些,這說明“華碩”的確是品質優良的油茶品種[12],但其最適采收期要比普通油茶晚。
CoACP、CoSAD和CoFAD2是在油茶脂肪酸代謝途徑中調控不飽和脂肪酸含量的關鍵酶,不同品種由于成熟期的差異在相同日期采摘的果實中相對表達量不同,“望城1號”的這3個基因表達高峰期均比“華碩”早1月有余,這也說明“望城1號”成熟期比“華碩”早1個月。根據相同發育時期種仁中基因表達量比較,不同油茶品種中CoACP、CoSAD和CoFAD2表達規律基本一致。結合脂肪酸成分和這3個基因表達蛋白的主調控作用分析,發現雖然調控產物含量變化和對應的基因表達規律大部分一致,但是“華碩”的CoFAD2表達規律與其亞油酸含量變化有所不符。有資料顯示油菜、玉米、大豆等作物FAD2以多拷貝形式存在[13-15],油茶FAD2也可能是多拷貝的,不同拷貝的FAD2在同一個體存在表達差異,同時相同拷貝FAD2在不同個體間也存在表達差異,因此導致本試驗中選用的CoFAD2表達規律與“望城1號”亞油酸含量變化一致,與“華碩”不符的結果。
[1]龍正海, 王道平.油茶籽油與橄欖油化學成分研究[J].中國糧油學報, 2008,23(2): 121-123
[2]廖書娟, 吉當玲, 童榮華.茶油脂肪酸組成及其營養保健功能[J].糧食與油脂, 2005 (6) : 7-91
[3]Majeru s P W. Acyl carrier protein. Structural requirements for function in fatty acid biosynthesis [J]. Journal of Biological Chemistry, 1967, 242 (10): 2325-2332
[4]譚曉風, 王威浩, 劉卓明, 等. 油茶ACP 基因的全長cDNA 克隆及序列分析[J]. 中南林業科技大學學報, 2008, 28(4): 8-14
[5]Kachroo A, Shanklin J, Whittle E, et al .The Arabidopsis stearoy-l acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis[J]. Plant Molecular Biology, 2007, 63(2) : 257-271
[6]張黨權, 譚曉風, 陳鴻鵬,等. 油茶SAD 基因的全長cDNA 克隆及生物信息學分析[J]. 林業科學, 2008, 44(2): 155-159
[7]Marillia E F, Taylor D. Cloning andnucleotide sequencing of a cDNA encoding a Brassica carinata FAD2 (Accession No. AF124360) [J]. Plant Physiology, 1990, 120:339
[8]譚曉風, 陳鴻鵬, 張黨權,等. 油茶FAD2 基因全長cDNA 的克隆和序列分析[J]. 林業科學, 2008, 44(3): 70-75
[9]王小藝, 曹一博, 張凌云,等. 油茶生長發育過程中脂肪酸成分的測定分析[J]. 中國農學通報, 2012, 28 (13) : 76-80
[10]孫美蓮, 王云生, 楊冬青. 茶樹實時熒光定量PCR分析中內參基因的選擇[J]. 植物學報, 2010, 45 (5): 579-587
[11]肖鋼, 張宏軍, 彭琪, 等.甘藍型油菜油酸脫氫酶基因(fad2)多個拷貝的發現及分析[J]. 作物學報, 2008, 34 (9): 1563-1568
[12]譚曉風, 袁德義, 袁軍, 等. 大果油茶良種“華碩”[J]. 林業科學, 2011, 47(12): 150
[13]Mikkilineni V, Rocheford T R. Sequence variation and genomic organization of fatty acid desaturase-2 (fad2) and fatty acid desaturase-6 (fad6) cDNAs in maize[J]. Theoretical and Applied Genetics, 2003, 106: 1326-1332
[14]Schlueter J A,Vasylenko-Sanders I F, Deshpande S, et al. TheFAD2 gene family of soybean: Insights into the structural and functional divergence of a paleopolyploid genome[J]. Crop Science, 2007, 47: 14-26
[15]Hernández M L, Mancha M, Martinez-Rivas J M. Molecular cloning and characterization of genes encoding two microsomal oleate desaturases (FAD2) from olive[J]. Phytochemistry, 2005, 66: 1417-1426.
