不同氮源對骨條藻光合活性、生長和中性脂積累的影響
2014-05-08 11:52:02付寶龍潘光華朱大玲才金玲張寶玉
天津科技大學學報 2014年3期
關鍵詞:油脂
付寶龍,潘光華,朱大玲,才金玲,張寶玉
(1. 天津科技大學海洋科學與工程學院,天津 300457;2. 中國科學院海洋研究所,青島 266071)
中國擁有近 300萬平方千米海域,海岸線長達32,000千米,海洋藻類資源豐富,尤其是海洋微藻資源.這些微藻是海洋初級生產力,為其他海洋生物提供食物來源,維持近海生態系統.然而,一旦近海發生富營養化,某些微藻會出現短時間內的爆發性增長,形成赤潮,造成嚴重的生態災難.因此,研究赤潮微藻的生物學特性、揭示其快速增殖的機制、解釋赤潮產生的原因是目前研究的熱點.如果從另外的視角來看赤潮微藻,是否可以利用赤潮微藻的快速增殖,為人類解決能源問題,則是一個值得高度關注的研究課題.
赤潮微藻主要集中在甲藻、硅藻、針胞藻和定鞭藻等門類中,其中以甲藻類和硅藻類所占的種類最多[1].一般微藻的細胞中均含有油脂,但不同微藻的油脂含量有明顯差異,甚至同一種的不同品系也存在著很大差別,油脂含量較高的微藻主要集中于綠藻、硅藻、金藻等真核微藻中[2].目前,微藻生物能源的研究主要集中在綠藻和硅藻上[3],其中骨條藻屬的中肋骨條藻是一種常見的硅藻屬赤潮藻,具有很高的油脂含量(14%~51%)[4],同時作為海產動物養殖中蝦蟹的餌料,其培養技術已經相當成熟,有一定的培養規模;但是,以骨條藻作為生產生物能源的原料,關注其油脂積累過程和油脂含量提高的研究還不多見.因此,本實驗研究了培養液中以不同濃度的硝酸鈉、氯化銨和尿素作為氮源對骨條藻的光合活性、生長和中性脂積累的影響,希望能夠為生產生物能源的大規模培養提供經驗.
1 材料與方法
1.1 實驗材料與培養方法
骨條藻采集自天津塘沽河口水域,經本實驗室分離純化并進行形態學和分子生物學鑒定.
微藻在實驗室的光照培養箱中進行培養,培養溫度為(20±1)℃,光照度為 5,000,lx,光/暗周期為12,h/12,h,每個150,mL三角瓶中培養100,mL藻液,所用培養基為人工海水配制的 f/2培養基[5],單藻種靜置培養,培養周期為 8,d.實驗采用 3種氮源分別為硝酸鈉、氯化銨和尿素,以 f/2培養基的標準氮源濃度 8.8×10-4,mol/L 為基準,按照 1/100、1/50、1/20、1/10、1/5、1/2和標準f/2設定氮源濃度由低到高依次為 8.8×10-6、1.8×10-5、4.4×10-5、8.8×10-5、1.8×10-4、4.4×10-4、8.8×10-4,mol/L.
1.2 分析方法
1.2.1 光合活性的測定
本實驗使用IMAGING–PAM型調制脈沖熒光儀(德國 Heinz Walz GmbH 公司),利用調制葉綠素熒光技術(pulse amplitude modulation,PAM),測量樣品的葉綠素熒光參數來反映藻細胞的光合活性[6].具體方法為:每瓶藻液各取250,μL分別加到96孔板的3個孔內,使用 IMAGING–PAM 型調制脈沖熒光儀測量樣品的潛在最大光合效率 Fv/Fm和實際光合效率Y(Ⅱ).每個樣品在測定前暗適應 15,min.設定光合有效輻射(photosynthetically active radiation,PAR)為24,μmol/(m2·s).
1.2.2 生長測定
對處于指數生長末期的骨條藻的藻液進行500~750,nm 的吸光度光譜掃描,發現在 655,nm 處有1個吸光度低峰,680,nm處有1個吸光度高峰(葉綠素 a的吸收峰).由于不同氮源濃度會對骨條藻的葉綠素a的含量產生影響,而 655,nm處的吸光度是整個吸收光譜的最低點,該點處光吸收值受色素含量影響最小,可以看作僅與藻體細胞密度相關,因此選擇 655,nm處的吸光度來測量骨條藻的生長.測定藻液在 655,nm 下的吸光度,藻細胞數用血球計數板計數,根據多次實驗得到的回歸方程 Y=22.94×A655+0.032,3(R2=0.999,9)計算細胞密度 Y(106,mL–1).
1.2.3 油脂測定
前人的研究結果[7]表明,經尼羅紅染色后的微藻在 480,nm光激發下,位于 580,nm左右的熒光峰的相對熒光強度與稱質量法測定的油脂含量呈線性正相關.因此,對處于指數生長末期的骨條藻的藻液使用 Hitachi F–4500型熒光分光光度計(日本日立公司)進行 500~750,nm 掃描,確定骨條藻尼羅紅染色的熒光峰在575,nm.
對不同濃度下藻細胞在該熒光峰的相對熒光強度進行測定,同時用稱質量法測定各個樣品的油脂含量,可以得出該熒光峰的相對熒光強度與藻細胞的油脂含量的關系式.具體方法為:1,mL藻液加 0.01,mL尼羅紅染料(丙酮溶解,質量濃度為 0.1,g/L)染色20,min后,使用 Hitachi F-4500型熒光分光光度計,以 480,nm 光激發,測定 500~750,nm 熒光發射光譜,以確定特殊熒光峰,再以該熒光峰的熒光強度減去藻細胞在該波長處的自身熒光強度和尼羅紅在該波長處的熒光強度,得到相對熒光強度.通過該熒光峰的相對熒光強度與藻細胞的油脂含量的關系式可得到細胞中油脂的含量.
1.2.4 數據處理
用SPSS 17.0軟件對數據進行單因素方差分析.使用Origin 8.0軟件計算平均值和標準偏差.
2 結果與分析
2.1 不同濃度的3種氮源對骨條藻光合活性的影響
圖 1—圖 6是 3種氮源對骨條藻 Fv/Fm和Y(Ⅱ)的影響.從圖中可以看出,各組的 Fv/Fm 和Y(Ⅱ)在接種后第 1天都有所升高,同時能正常生長的氮源種類相同的各組之間的差距不大,表明此時培養基中的營養鹽充足,骨條藻的光合活性正常.隨著培養的進行,各組的Fv/Fm和Y(Ⅱ)都出現了下降,氮源濃度越低下降的速度越快.在第 7天時,3種氮源都只有濃度為 4.4×10-4,mol/L和 8.8×10-4,mol/L的兩組的 Fv/Fm 和 Y(Ⅱ)能被測出,表明隨著氮源的消耗,各組的光合活性都受到了抑制.在第 6、7天,氯化銨組后兩組的 Fv/Fm 和 Y(Ⅱ)都高于硝酸鈉組和尿素組后兩組的Fv/Fm和Y(Ⅱ).在第8天時,硝酸鈉組和尿素組后兩組的 Fv/Fm 和 Y(Ⅱ)都出現了上升,而氯化銨組后兩組的 Fv/Fm 和 Y(Ⅱ)則持續下降,表明硝酸鈉組和尿素組中開始有部分藻細胞死亡,釋放了一些氮源,而氯化銨組則能繼續保持生長.因此,氯化銨是比較適合骨條藻生長的氮源.
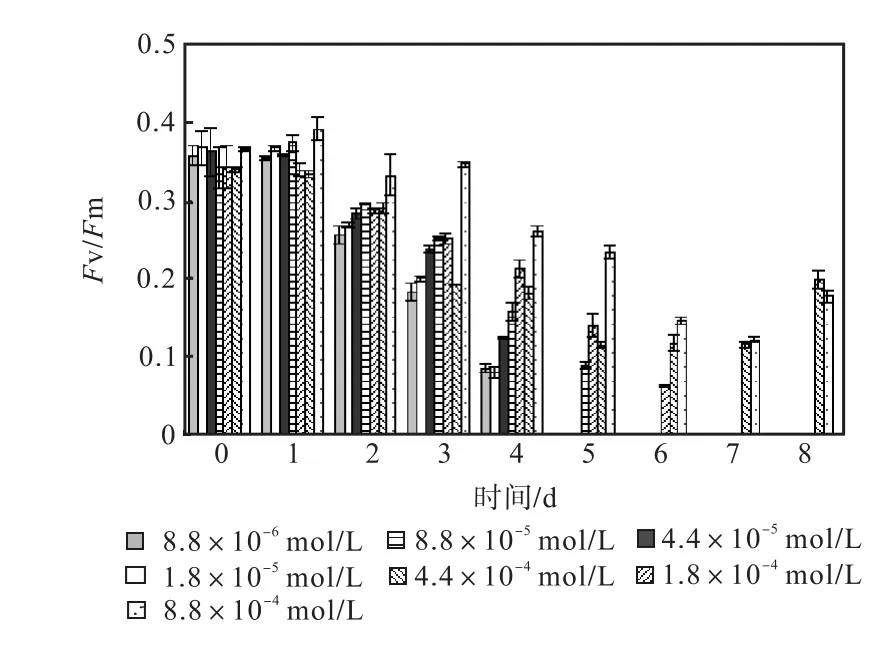
圖1 硝酸鈉對骨條藻Fv/Fm的影響Fig.1 Effect of NaNO3 on Fv/Fm of Skeletonema
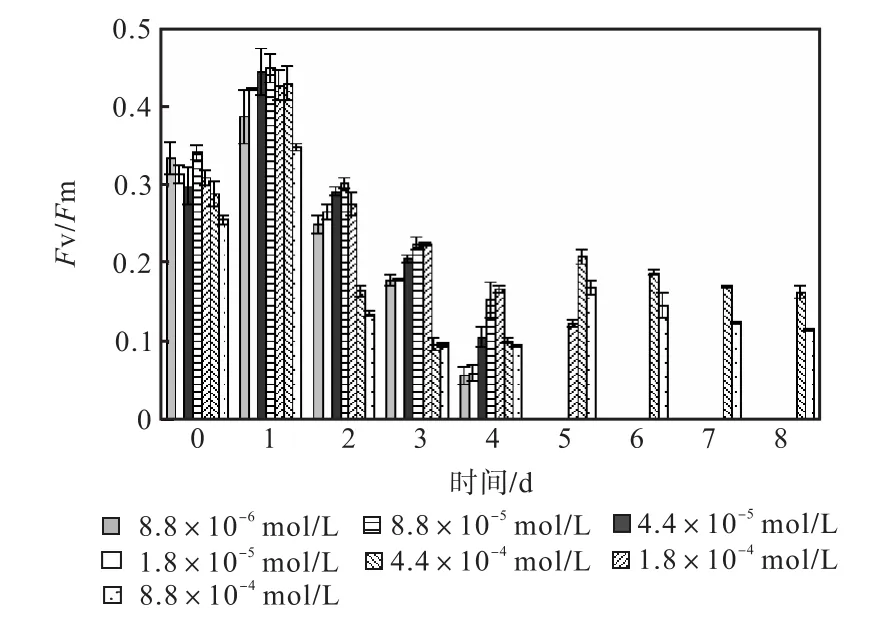
圖3 氯化銨對骨條藻Fv/Fm的影響Fig.3 Effect of NH4Cl on Fv/Fm of Skeletonema

圖5 尿素對骨條藻Fv/Fm的影響Fig.5 Effect of urea on Fv/Fm of Skeletonema
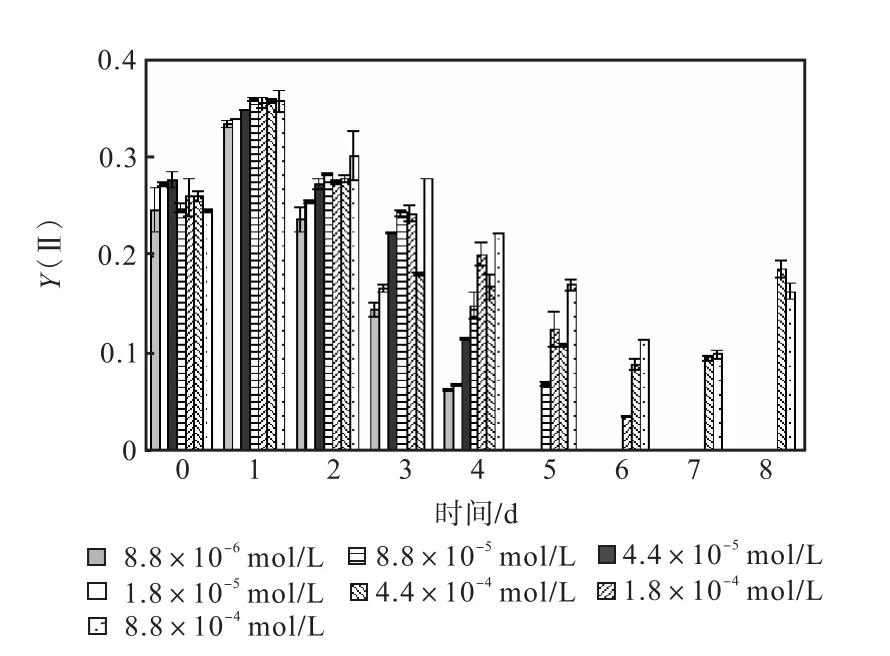
圖2 硝酸鈉對骨條藻Y(Ⅱ)的影響Fig.2 Effect of NaNO3 on Y(Ⅱ)of Skeletonema
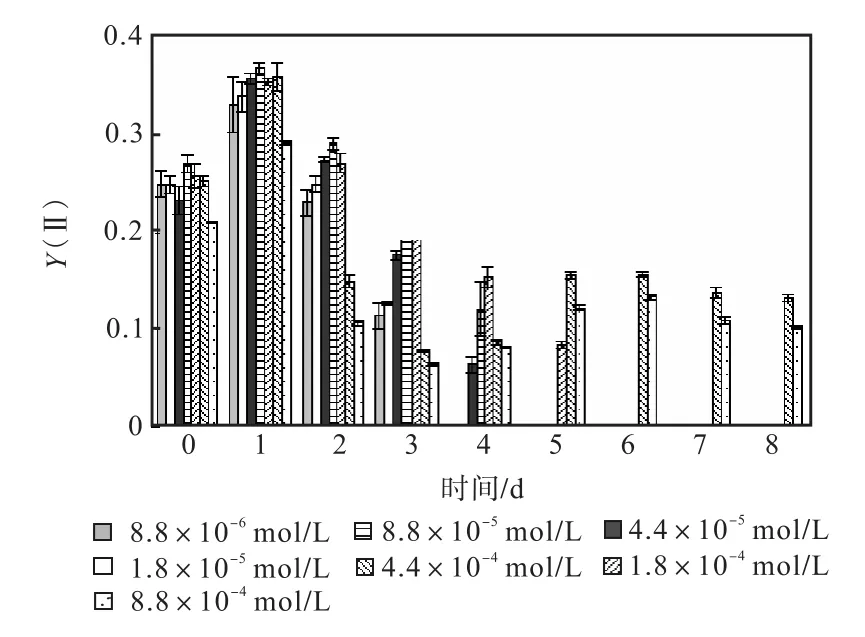
圖4 氯化銨對骨條藻Y(Ⅱ)的影響Fig.4 Effect of NH4Cl on Y(Ⅱ)of Skeletonema
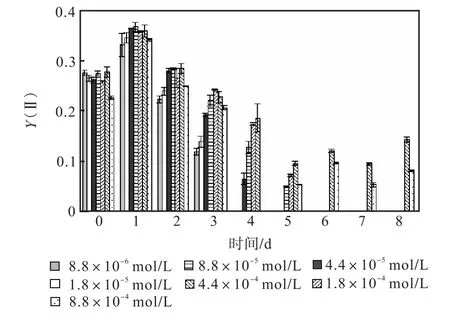
圖6 尿素對骨條藻Y(Ⅱ)的影響Fig.6 Effect of urea on Y(Ⅱ)of Skeletonema
2.2 不同氮源最適合骨條藻生長的濃度確定
各組的骨條藻在第1天的生長速度最快,除了氮濃度最高的 8.8×10-4,mol/L的氯化銨和尿素外,其他各組生物量的增長都超過110%.這與藻細胞的光合活性在第1天的升高表現一致,表明此時培養基中的營養充足,適合骨條藻的生長.而骨條藻在氮濃度為8.8×10-4,mol/L 尿素中的生長好于氮濃度為 8.8×10-4mol/L氯化銨,則表明高濃度的氯化銨對骨條藻生長的抑制比高濃度的尿素強.能正常生長的各組氮源濃度的最大生物量基本都出現在第8天,其中氯化銨組的生長是最好的,其次是尿素組,最后的是硝酸鈉組,3種氮源中生長最好的都是氮濃度為 4.4×10-4,mol/L,這與整個培養過程中藻細胞的光合活性的表現是一致的.
3種氮源的最大生物量的比較如圖7所示,硝酸鈉組和尿素組的最大生物量與氯化銨組都有極顯著差異(P<0.01),硝酸鈉組的最大生物量與尿素組有顯著差異(P<0.05).因此,可以確定最適合骨條藻生長的氮源是氯化銨,其次是尿素,最后的是硝酸鈉,3種氮源的最適氮濃度都是4.4×10-4,mol/L.
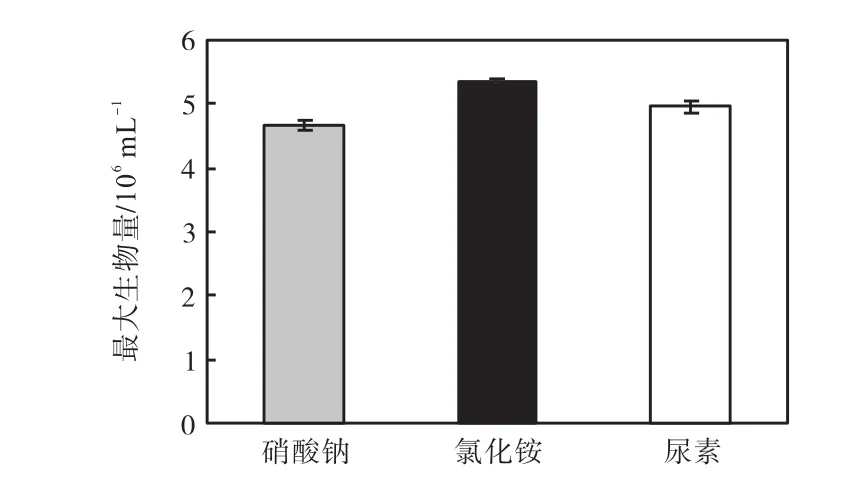
圖7 不同氮源最適合骨條藻生長的濃度下的最大生物量Fig.7 Maximum biomass of Skeletonema at the optimum concentration of different nitrogen sources for its growth
2.3 不同濃度的3種氮源對骨條藻中性脂積累的影響
2.3.1 不同濃度硝酸鈉對骨條藻中性脂積累的影響
圖 8和圖 9是培養過程硝酸鈉組各組的中性脂積累的過程.前 4,d各組每 106個細胞的尼羅紅染色相對熒光強度基本保持穩定,在第 5天,氮濃度較低的前5組的熒光強度開始出現增長,基本表現為氮源濃度越低的熒光強度越強.在第 7天各組的熒光強度都出現了大幅度的增長,在第 8天即出現了下降.其中 1,mL藻液的尼羅紅染色相對熒光強度是藻細胞中油脂總量的指標,而 106個細胞的尼羅紅染色相對熒光強度則是藻細胞中油脂百分含量的指標,兩者的最高值都出現在第 7天起始氮濃度為 1.8×10-4,mol/L的藻液,按照 106個細胞的尼羅紅染色相對熒光強度與藻細胞的油脂含量關系式得出其油脂含量為28.46%.

圖8 硝酸鈉對骨條藻 1,mL藻液尼羅紅染色相對熒光強度的影響Fig.8 Effect of NaNO3 on the relative fluorescence values of 1,mL Skeletonema solution stained with Nile red

圖9 硝酸鈉對骨條藻 106個細胞尼羅紅染色相對熒光強度的影響Fig.9 Effect of NaNO3 on the relative fluorescence values of 106 algae cells of Skeletonema stained with Nile red
2.3.2 不同濃度的氯化銨對骨條藻中性脂積累的影響
圖 10和圖 11是培養過程氯化銨組各組的中性脂積累的過程.前 4,d各組106個細胞的尼羅紅染色相對熒光強度基本保持穩定,在第 5天,氮濃度較低的前5組的熒光強度同樣開始出現增長,熒光強度與氮源濃度的負相關關系比硝酸鈉組更明顯.同樣在第7天各組的熒光強度都出現了大幅度的增長,在第8天即出現了下降.其中 1,mL藻液和 106個細胞的尼羅紅染色相對熒光強度最高的都是起始氮濃度為8.8×10-6,mol/L的藻液第 7天的值,按照 106個細胞的尼羅紅染色相對熒光強度與藻細胞的油脂含量關系式得出其油脂含量為31.07%.

圖10 氯化銨對骨條藻 1,mL藻液尼羅紅染色相對熒光強度的影響Fig.,10 Effect of NH4Cl on the relative fluorescence values of 1,mL Skeletonema solution stained with Nile red
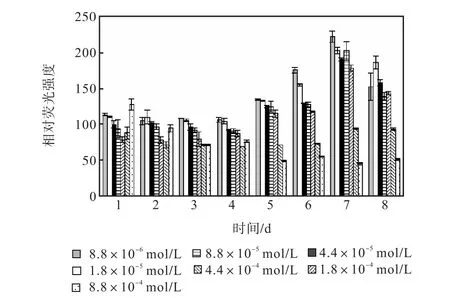
圖11 氯化銨對骨條藻 106個細胞尼羅紅染色相對熒光強度的影響Fig.,11 Effect of NH4Cl on the relative fluorescence values of 106 algae cells of Skeletonema stained with Nile red
2.3.3 不同濃度的尿素對骨條藻中性脂積累的影響
圖 12和圖 13是培養過程中尿素組各組的中性脂積累的過程.前 5,d各組 106個細胞的尼羅紅染色相對熒光強度基本保持穩定,在第 6天,氮濃度較低的前5組的熒光強度開始出現增長,熒光強度與氮源濃度同樣呈負相關關系.在第 7天各組的熒光強度都出現了大幅度的增長,在第 8天即出現了下降.其中1,mL藻液和106個細胞的尼羅紅染色相對熒光強度最高的都是起始氮濃度為1.8×10-5,mol/L的藻液第7天的值,按照 106個細胞的尼羅紅染色相對熒光強度與藻細胞的油脂含量關系式得出其油脂含量為27.99%.
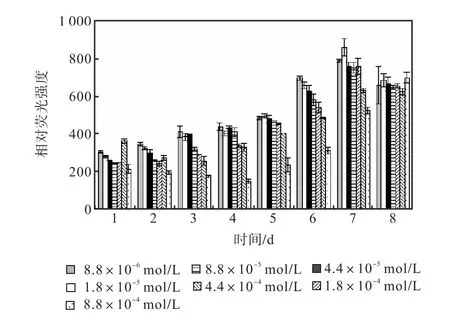
圖12 尿素對骨條藻1,mL藻液尼羅紅染色相對熒光強度的影響Fig.,12 Effect of urea on the relative fluorescence values of 1,mL Skeletonema solution stained with Nile red
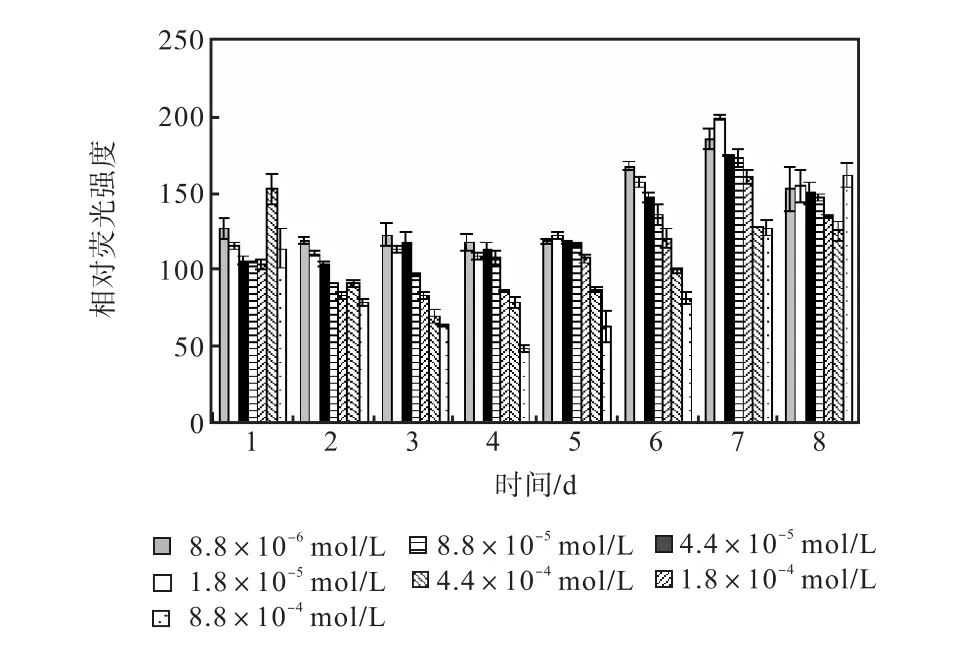
圖13 尿素對骨條藻 106個細胞尼羅紅染色相對熒光強度的影響Fig.,13 Effect of urea on the relative fluorescence values of 106 algae cells of Skeletonema stained with Nile red
2.3.4 綜合分析
3種氮源組中出現的1,mL藻液和 106個細胞尼羅紅染色相對熒光強度如圖14所示.
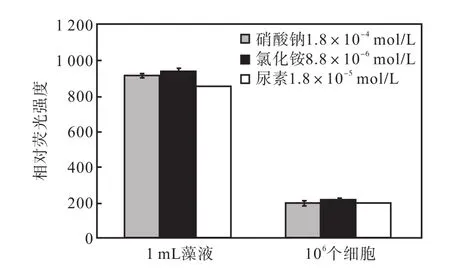
圖14 最適合油脂積累的氮濃度下3種氮源1,mL藻液和106個細胞尼羅紅染色相對熒光強度Fig.,14 Effect of three nitrogen sources at the optimum concentration on the relative fluorescence values of 1,mL algae solution and 106 algae cells staoned with Nile red
1,mL藻液的尼羅紅染色相對熒光強度表示油脂總量,106個細胞尼羅紅染色相對熒光強度可以計算出油脂的含量.可以看出,氯化銨的兩項值都是最高的,同時,所用的氮源量也是最低的,因此氯化銨是最適合骨條藻中性脂積累的氮源.
3 討 論
氮是浮游植物細胞內蛋白質、核酸、磷脂和葉綠素等的基本元素,在浮游植物生命活動中占有重要的地位.海水中的無機氮主要包括-N和-N,它們是能被海洋浮游植物直接利用的氮源,而尿素在有機氮源中是研究比較廣泛的一種[8–9].通常認為浮游植物最先吸收-N,與細胞內碳水化合物衍生的酮酸通過轉氨基作用直接合成氨基酸;而-N和-N則必須經過相應的硝酸還原酶和亞硝酸還原酶還原成-N[10].在大多數情況下,甚至當氧化態氮源(如-N 和-N)更充足的時候,浮游植物更傾向于利用還原態氮源(-N和尿素)[11],藻細胞蛋白質中的氮呈還原態,利用或者有機態的氮源顯然比利用硝酸鈉中高氧化態的要經濟.同時,對的吸收,能引起培養液中 pH 的下降,可以緩沖因光合作用而導致的藻液 pH上升,有利于藻類的生長[12].而尿素除了能為微藻的生長提供氮源外,還能夠提供碳源.因此,在3種氮源中,氯化銨是最適合骨條藻生長的氮源,尿素次之,硝酸鈉最后.
當營養鹽總水平足以滿足浮游植物生長時,海洋浮游植物的氮磷吸收比例基本上是遵循 Redfield比值N∶P=16∶1,因此常用此比值來判斷浮游植物營養鹽的相對受限情況[13].當N/P<16時,表明氮相對不足,N/P>16時,表明磷相對不足.3種氮源最適合骨條藻生長的氮起始濃度都是 4.4×10-4mol/L,這時培養液中的氮磷比是13.74∶1,稍低于Redfield比值N∶P=16∶1,表明處于缺氮狀態,但能滿足骨條藻生長的需要;當氮濃度達到 8.8×10-4,mol/L時,培養液中的氮磷比是27.48∶1,此時限制骨條藻生長的是磷元素.藻細胞利用-N作為氮源是不經濟的,同時,過高濃度的和尿素對藻類是有毒的[12].因此,過高的3種氮源濃度對骨條藻的生長并無促進作用,甚至會抑制骨條藻的生長.
Spoehr和 Mihier[14]最早證明,氮缺乏可誘導Chiorella pyrenoidosa中脂類含量的增加,該現象后來在許多其他藻類中得到了證實.到目前為止,氮缺乏誘導脂類積累的相關機制還不清楚.微藻合成油脂大體上可以分為兩個階段,即細胞增殖期和油脂積累期,細胞增殖期是微藻消耗培養基中的氮源,合成蛋白質,以保證藻體細胞代謝旺盛;接著藻體細胞開始大量消耗碳、氮源,并合成積累大量的油脂[15].低氮培養基中的骨條藻基本都是在第 5天進入油脂積累期,隨著培養時間的增加,氮源逐漸被消耗殆盡,細胞依次進入氮饑餓的狀態,細胞內的蛋白質合成代謝受到抑制而降低,相對的細胞內油脂的合成得到了加強,在一定程度上也會提高微藻細胞的油脂含量[16].因此,骨條藻的中性脂大量積累出現在培養后期.但當培養到第 8天時,培養基中的各種營養元素均被大量消耗,無法為細胞的各種代謝過程提供充足的營養,使得細胞開始消耗自身的脂類維持生物量的穩定,導致了細胞內油脂含量的降低.骨條藻是一種赤潮藻,有增殖快、衰退也快的特點,因此要利用它的脂類,需要選擇合適的采收時間.
4 結 論
最適合骨條藻的氮源是氯化銨,其次是尿素,最后是硝酸鈉,3種氮源最適合骨條藻生長的氮濃度都是 4.4×10-4,mol/L.最適合骨條藻中性脂積累的氮源是氯化銨,其最適濃度為 8.8×10-6,mol/L,其次是尿素,其最適濃度為 1.8×10-5,mol/L,最后是硝酸鈉,其最適濃度為 1.8×10-4,mol/L.在高接種量(106,mL-1)、使用 f/2培養基的條件下,不同濃度的氮源條件下骨條藻的最終生物量差距不大,通過調節氮源濃度可以使中性脂的積累量大幅度上升,油脂總量的高低與百分含量的高低基本一致.同時,氯化銨也是一種容易獲得的肥料,因此,用骨條藻生產生物能源,以氯化銨作為氮源是最適合的.
[1] 鄒景忠. 赤潮生物與赤潮災害研究[M]//曾呈奎. 中國海洋科學研究與開發. 青島:青島出版社,1992:284-287.
[2] Hu Qiang,Sommerfeld M,Jarvis E,et al. Microalgal triacyl glycerols as feedstocks for biofuel production:Perspectives and advances[J]. The Plant Journal,2008,54(4):621-639.
[3] Chisti Y. Biodiesel from microalgae[J]. Biotechnology Advances,2007,25(3):294-306.
[4] Mata T M,Martins A A,Caetano N S. Microalgae for biodiesel production and other applications:A review[J].Renewable and Sustainable Energy Reviews,2010,14(1):217-232.
[5] Guillard R R L,Ryther J H. Studies of marine planktonic diatom. I. Cyclotella nana Hustedt,and Detonula confervacea(cleve)Gran[J]. Canadian Journal of Microbiology,1962,8(2):229-239.
[6] Krause G H,Weis E. Chlorophyll fluorescence and photosysthesis:The basis[J]. Annual Review Plant Physiology and Plant Molecular Biology,1991,42:313-349.
[7] Cooksey K E,Guckert J B,Williams S A,et al. Fluorometric determination of the neutral lipid content of microalgal cell using Nile Red[J]. Journal of Microbiological Methods,1987,6(6):333-345.
[8] Storm S L,Bright K J. Inter-strain differences in nitrogen use by the coccolithophore Emiliania huxleyi,and consequences for predation by a planktonic ciliate[J]. Harmful Algae,2009,8(5):811-816.
[9] Glibert P M,Trice T M,Michael B,et al. Urea in the tributaries of the Chesapeake and Coastal Bays of Maryland[J]. Water,Air and Soil Pollution,2005,160(1/2/3/4):229-243.
[10]Berges J A,Mulholland M R. Enzymes and Nitrogen Cycling[M]// Capone D G,Bronk D A,Mulholland M R,et al. Nitrogen in the Marine Environment. Elsevier Press,2008:1385-1444.
[11]Fan C,Glibert P M,Alexander J,et al. Characterization of urease activity in three marine phytoplankton species,Aureococcus anophagefferens,Prorocentrum minimum,and Thalassiosira weissflogii[J]. Marine Biology,2003,142(5):949-958.
[12]滕亞娟. 不同氮源對骨條藻生長的影響[J]. 水利漁業,2006,26(4):55-57.
[13]Redfield A C. The biological control of chemical factors in the environment[J]. American Scientist,1958,46(3):205-222.
[14]Spoehr H A,Milner H W. The chemical composition of Chlorella;effect of environmental conditions[J]. Plant Physiology,1949,24(1):120-149.
[15]薛飛燕,張栩,譚天偉. 微生物油脂的研究進展及展望[J]. 生物加工過程,2005,3(1):23-28.
[16]繆曉玲. 藻類可再生能源的利用及藻細胞抗環境脅迫的研究[D]. 北京:清華大學,2004.
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
