南京地區2011-2012年度門診嬰幼兒輪狀病毒腹瀉的分子流行病學研究
2014-04-09 00:51:34蔣翠蓮顧平清葉莉莉付建光
中國人獸共患病學報 2014年3期
關鍵詞:嬰幼兒
蔣翠蓮,顧平清,葉莉莉,楊 艷,付建光
A組輪狀病毒(Rotavirus, RV)是引起嬰幼兒及多種動物急性重癥腹瀉的最主要病原,全球每年約有1.3億嬰幼兒感染RV,其中200萬到400萬病例住院,死亡病例更是高達60萬,死亡病例大部分發生在發展中國家[1-3]。
RV屬于呼腸孤病毒屬,核心為雙股RNA,由11個不連續的RNA片段組成,每個片段編碼1個或2個結構蛋白(VP1~VP7)或非結構蛋白(NSP1~NSP6)[4]。其中VP4和VP7是病毒的表面中和抗原,根據VP4和VP7的抗原性差異可分為35個P基因型(相對應14個P血清型)和27個G基因型(15個G血清型),其中幾個基因型可同時感染人和動物[5-7]。
本文對2011年7月至2012年6月南京市婦幼保健院門診腹瀉嬰幼兒的RV感染特征及基因分型進行了初步的研究。
1 材料與方法
1.1標本 對2011年7月-2012年6月間在南京市婦幼保健院門診就診的腹瀉患兒(包括新生兒)糞便進行RV膠體金檢測,試劑盒采用輪狀病毒腺病毒抗原檢測試劑盒(杭州,艾博生物醫藥有限公司),陽性標本送至江蘇省疾病預防控制中心進行分型檢測,所有標本-70 ℃冰凍保存。
1.2標本處理 加入1 mL生理鹽水至1.5 mLEP管中,加入0.1 g固體糞便標本或0.1 mL液體糞便標本,置于漩渦震蕩器混勻,室溫靜置10 min,室溫下≥5 000 r/min離心5 min,吸取上清液,置-20 ℃冰箱保存。
1.3核酸提取 利用ABI自動核酸提取儀提取,試劑采用ABI 5X MagMAXTM-96 Viral Isolation Kit(美國ABI公司),提取方法按照試劑盒說明書。
1.4巢式逆轉錄聚合酶鏈反應(nRT-PCR)擴增片段 將RV陽性標本進行nRT-PCR擴增。試劑盒為Qiagen One Step RT-PCR Kit(德國QIAGEN公司)。反應采用國家CDC推薦的引物及反應條件[8-10],反應結束后取4 μL擴增產物, 在2%的瓊脂糖凝膠上以110V恒壓狀態電泳50 min, 通過GEL Doc 2000(Bio-Rad)凝膠成像系統觀察RV特異性擴增條帶。每批電泳用已測序證實為RV陽性的RT-PCR產物作為陽性對照。-20 ℃保存 PCR產物。
1.5序列測定和分析 將PCR陽性產物送公司進行純化和測序。從GenBank的核酸序列數據庫中下載多株國內外RV的核酸序列片段,將下載序列同我們檢測到的本地區的RV序列用DNAStar公司的MegA5.1軟件,采用CLUSTALW方法進行序列比對,利用neighbor-joining法構建進化樹。
1.6統計方法 采用SPSS 13.0軟件統計,計數資料比較采用χ2檢驗。
2 結 果
2.1流行病學特征 我們對2011年7月-2012年6月間的2 081例腹瀉患兒進行了RV膠體金檢測,共發現陽性病例165例,陽性率為7.9%。全年均有RV感染發生,高峰季節為10月份至次年的2月份(圖1)。94%的感染患兒都在2歲以下,其中1~3月齡感染患兒最多,13~24月齡為高發年齡段(圖2)。
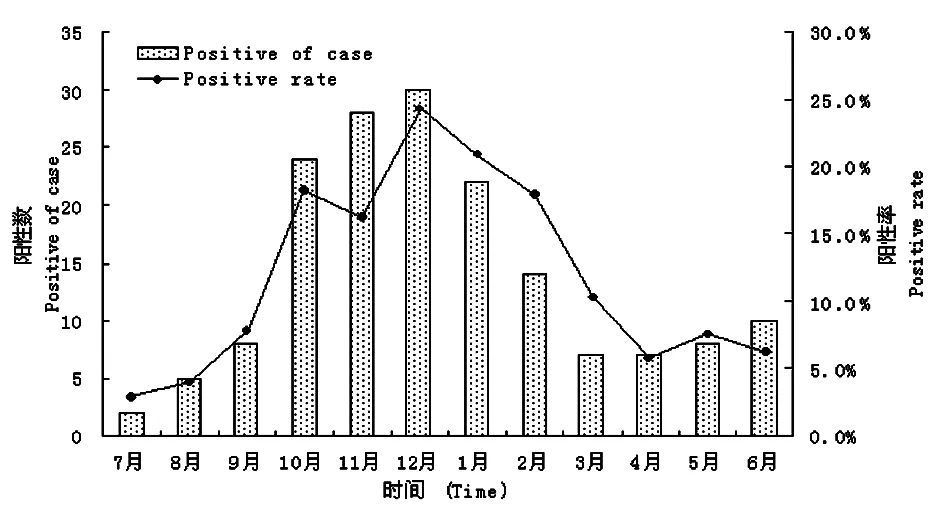
圖12011-2012年南京地區RV腹瀉兒童病例數的季節分布
Fig.1SeasonaldistributionofrotavirusdiarrheachildreninNanjingfrom2011to2012
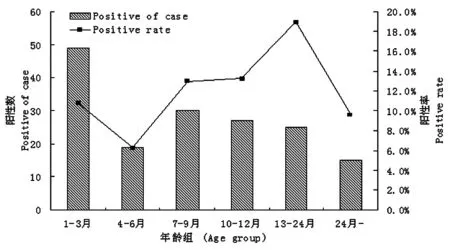
圖22011—2012年南京地區RV腹瀉兒童病例數的年齡分布
Fig.2AgedistributionofrotavirusdiarrheachildreninNanjingfrom2009to2012
2.2G/P分型 將165例RV陽性標本進行G/P分型。如表1所示,G分型結果中G1, G3, G9為主要G分型,占G分型的78.2%,這其中G9所占比例最大,約為35.2%,G未分型(GNT)及G混合型(GMIX)也占有一定比重為15.8%。P分型中P[8]則是最常見的P分型,占P分型的64.2%,P[4]為16.4%,P未分型(PNT)及P混合型(PMIX)共占19.4%。常見的G/P組合為:G1P[8](16.4%), G3P[8](13.9%), G9P[8](24.8%)。
表12011年至2012年南京地區輪狀病毒G/P分型
Tab.1RotavirusGandPgenotypesinNanjingfrom2011to2012
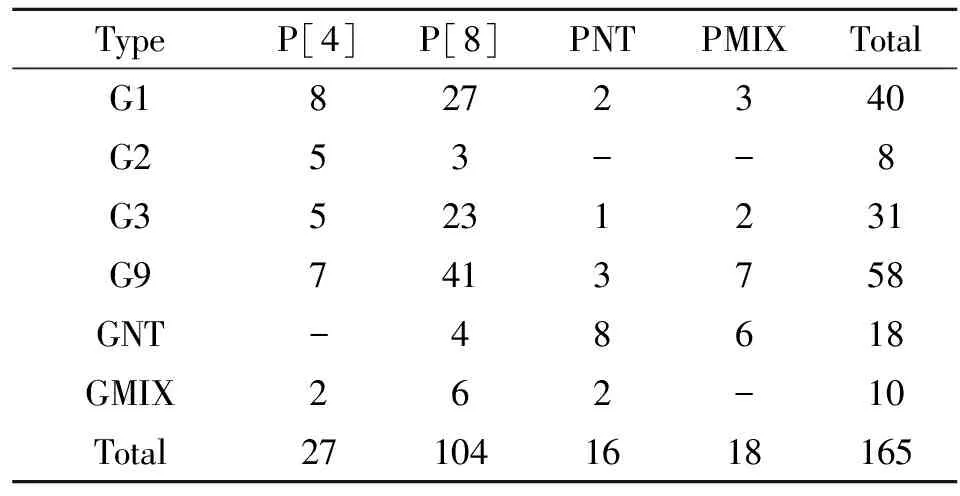
TypeP[4]P[8]PNTPMIXTotalG18272340G253--8G35231231G97413758GNT-48618GMIX262-10Total271041618165
2.3序列測定 通過對部分VP7及VP4基因第一輪擴增產物進行測序并與相應參考株進行對比,我們發現G1和G3毒株彼此間的同源性很高,分別為99.6%~100%及99.5%~100%,兩者所有毒株各分屬于一個亞型G1-Ic,G3-Ia,而G9毒株間的同源性為92.6%~100%,大部分毒株屬于G9-IV亞型,同時也有一小部分歸于G9-III亞型。P分型中,P[8]毒株間的同源性為87.2%~100%,主要為P[8]-II,少數為P[8]-IV。P[4]毒株間的同源性為97.5%~100%,與P[4]-Vc亞型參考株的同源性最高。
3 討 論
在我國每年RV感染引起的嬰幼兒腹瀉約有1 200萬例,造成了沉重的經濟負擔和社會負擔[11]。考慮到尚無特效的抗病毒藥物治療,因此預防性接種RV疫苗成為了首選方法。在許多國家,RV的感染率和死亡率都隨著疫苗的廣泛接種而大幅下降[12]。目前我國接種比較廣泛的是蘭州生物制品生產的G10P[12]型LLR RV疫苗,接種后感染率也已經得到了降低[13]。但是隨著時間推移,RV不斷變異,流行毒株也在發生著變化,因此掌握更全面的毒株信息可以為將來研發更加有效的疫苗提供必要的科學依據。
在本研究中,我們發現RV感染是引起南京地區嬰幼兒腹瀉的重要原因。我們對2011年7月-2012年6月間共2 081例腹瀉患兒進行了RV膠體金檢測,陽性病例165例,陽性率為7.9%。這明顯低于李梅等[14]及林謙等[15]報道的南京地區的陽性率,分析原因可能是我們樣本來源的醫院為婦幼保健院,病例以新生兒和3月齡以下的嬰幼兒為主,而這部分群體的腹瀉多為功能性腹瀉,與病毒感染無關;此外,我們在進行初篩時所應用的檢測試劑盒為杭州艾博生物的試劑盒,而未采用更加普及的萬泰的試劑盒。兩者之間的檢測特異性和靈敏性未曾做過比較,也未見相關報道,考慮有可能是試劑盒靈敏度低導致總體陽性率偏低。RV腹瀉又稱為小兒秋冬季腹瀉,圖1表明全年均有RV感染發生,但高峰季節為10月份至次年的2月份,這與多數地區的報道一致[16-18]。對不同年齡組的RV腹瀉患兒進行分析發現94%的感染患兒都在2歲以下,其中1~3月齡感染患兒最多,13~24月齡為高發年齡段,這可能與該年齡段患兒體內的母體抗體已經基本消失,而且學會走路接觸的東西繁多喜歡吃手指等多種原因有關。
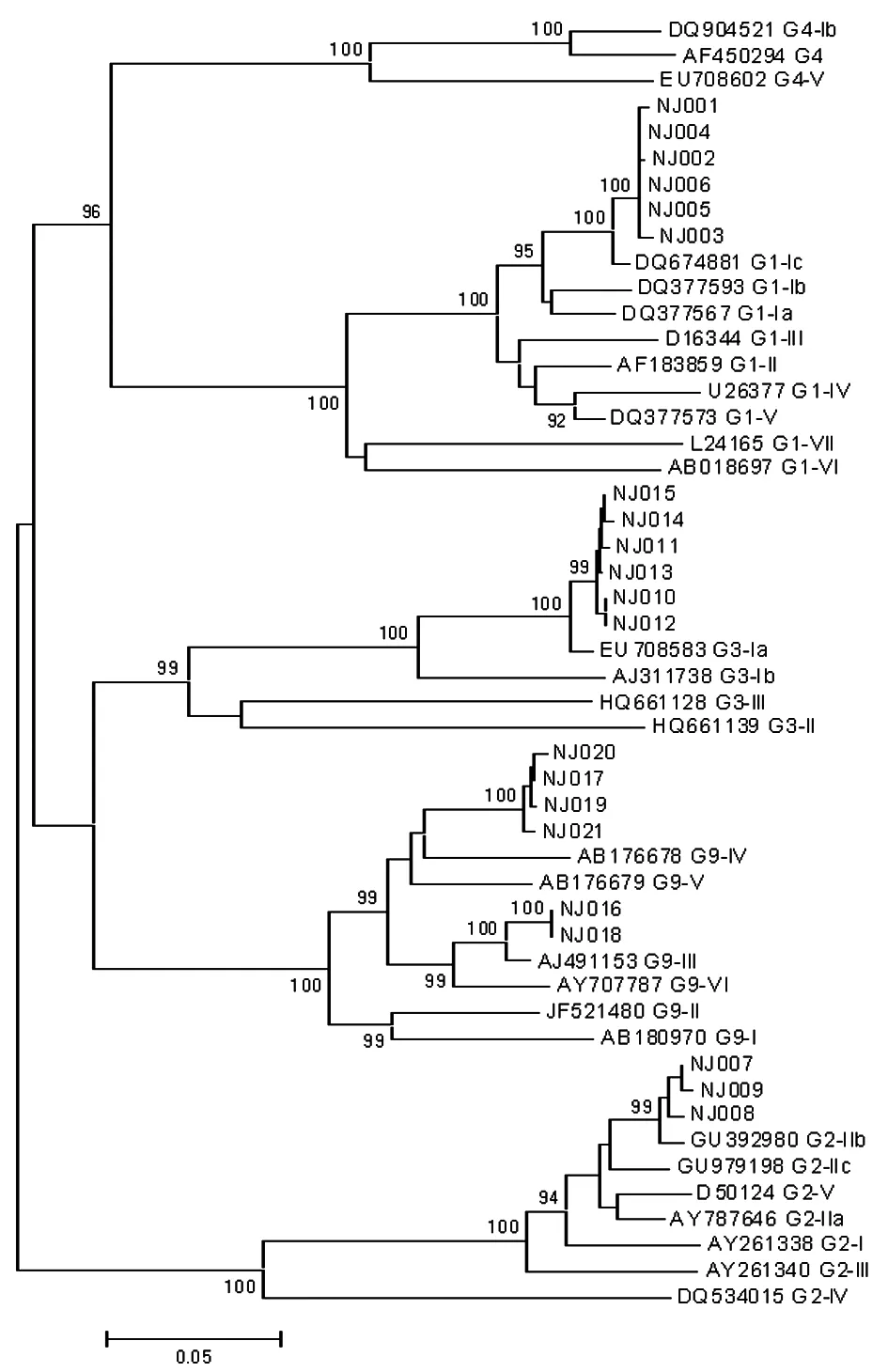
圖3輪狀病毒G分型VP7核苷酸序列進化分析
Fig.3PhylogenetictreesforthenucleotidesequencesoftheVP7regionofGgenotyping
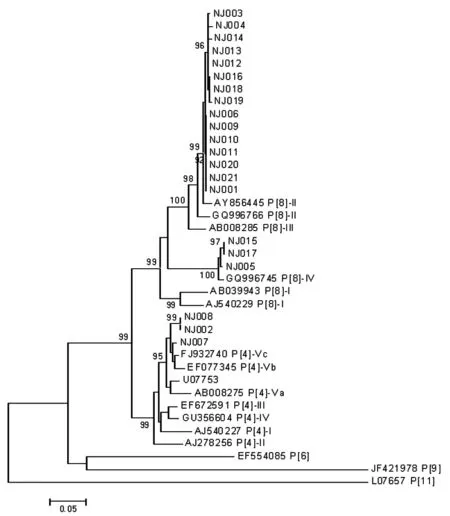
圖4輪狀病毒P分型VP4核苷酸序列進化分析
Fig.4PhylogenetictreesforthenucleotidesequencesoftheVP4regionofPgenotyping
RV的遺傳多樣性與不同區域內新流行毒株的出現有關,常見的RV流行株包括G1P[8],G2P[4],G3P[8],G4P[8]和G9P[8][19]。國內的相關研究表明,我國最常見的流行株為G1P[8]和G3P[8],同時也有其他毒株的報道(G1P[4],G2P[4],G3P[4],G4P[8]和 G9P[8])[20-21]。此次研究發現南京地區G分型結果中G1, G3, G9為主要G分型,占G分型的78.2%,這其中G9所占比例最大,約為35.2%,G未分型(GNT)及G混合型(GMIX)也占有一定比重為15.8%。這與林謙等[15]的報道不同,G9型毒株開始占有最大比重,這提示G型流行毒株可能已經開始出現改變。1983年美國首次報道了G9型毒株,隨后在許多國家都開始出現G9感染的報道,并逐漸成為重要的流行株[22-24]。在我國,1994年首次出現關于G9毒株的報道,隨后常有散發報道[25-28]。章等[29]對1998年至2004年我國9個地區的G9型毒株進行監測分析發現,G9型檢出率有逐漸升高趨勢,總體檢出率為3.5%,早期型別組合為G9P[6],從2001年以后逐漸變為G9P[8]。在P分型的檢測中P[8]則是最常見的P分型,占P分型的64.2%,P[4]為16.4%,P未分型(PNT)及P混合型(PMIX)共占19.4%。這同世界大多數地區的報道一致,未有明顯改變。常見的G/P組合為:G1P[8](16.4%), G3P[8](13.9%), G9P[8](24.8%)。這同林等[15]報道的南京地區的G/P組合相似,但是G9P[8]取代了G3P[8]成為了優勢型別組合。
目前國內對G/P分型的亞型報道較少,我們進一步測序進行亞型比對,以便得到更加全面的信息。同相關參考株比對后我們發現G1和G3毒株彼此間的同源性很高,兩者所有毒株各分屬于一個亞型G1-Ic,G3-Ia; G9型毒株間的同源性為92.6%~100%,大部分毒株屬于G9-IV亞型,同時也有一小部分歸于G9-III亞型。章青等[29]的研究中1998年至2004年我國檢出的G9都屬于G9-III亞型,而我們的研究發現G9-IV亞型占據了更大的比例,這種亞型的轉變可能使得毒株的組織血型抗原(Histo-blood group antigens, HBGA)識別模序即中和抗原表位發生了變化,而這些抗原表位的改變也許同樣驅動著HBGA親和力的改變及人群易感性的改變[30],因此最終導致G9型毒株超越了G1型毒株成為流行株。P分型中,P[8]毒株間的同源性為87.2%~100%,主要為P[8]-II,少數為P[8]-IV。這與國外報道常見的P[8]-III有所差異[31-32],但是與武漢的報道相似[33],說明我國P[8]毒株主要為P[8]-II 亞型。3個P[4]毒株間的同源性為97.5%~100%,與P[4]-Vc亞型參考株的同源性最高。
綜上所述, RV感染是引起南京地區嬰幼兒腹瀉的重要病因,這其中G9P[8]是最主要的流行型別,同時G1P[8]和G3P[8]也占較大比例。亞型比對結果表明G9亞型已經出現了轉變,由G9-III亞型轉為G9-IV亞型為主,P[8]仍以P[8]-II 亞型為主。有鑒于此,進一步的加強監測,密切關注流行毒株的變化,同時得到完整的基因組信息也將是我們的下一步工作重點。
參考文獻:
[1]Parashar UD, Hummelman EG, Bresee JS, et al. Global illness and deaths caused by rotavirus disease in children[J]. Emerg Infect Dis, 2003, 9: 565-572.
[2]Musher DM, Musher BL. Contagious acute gastrointestinal infections[J]. N Engl J Med, 2003, 351: 2417-2427.
[3]Grimwood K, Bines JE. Rotavirus vaccines must perform in low-income countries too[J]. Lancet, 2003, 370(9601): 1739-1740.
[4]Bishop RF. Natural history of human rotavirus infection[J]. Arch Virol Suppl, 2003, 12, 119-128.
[5]Matthijnssens J, Ciarlet M, Heiman E, et al. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains[J]. J Virol, 2008, 82: 3204-3219. DOI: 10.1128/JVI.02257-07
[6]Matthijnssens J, Ciarlet M, McDonald SM, et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG)[J]. Arch Virol, 2011, 156: 1397-1413. DOI: 10.1007/s00705-011-1006-z
[7]Schumann T, Hotzel H, Otto P, et al. Evidence of interspecies transmission and reassortment among avian group A rotaviruses[J]. Virology, 2009, 386: 334-343. DOI: 10.1016/j.virol.2009.01.040
[8]Gouvea V, Glass RI, Woods P, et al. Polymerase chain reaction amplification and typing of rotavirus nucleic acid from stool specimens[J]. J Clin Microbiol, 1990, 28(2): 276-282.
[9]Iturriza-Gomara M, Kang G, Gray J. Rotavirus genotyping: keeping up with an evolving population of human rotaviruses[J]. J Clin Virol, 2004, 31(4): 259-265.
[10]Simmonds MK, Armah G, Asmah R, et al. New oligonucleotide primers for P-typing of rotavirus strains: Strategies for typing previously untypeable strains[J]. J Clin Virol, 2008, 42(4): 368-373. DOI: 10.1016/j.jcv.2008.02.011
[11]Jin H, Wang B, Fang ZY, et al. Hospital-based study of the economic burden associated with rotavirus diarrhea in eastern China[J]. Vaccine, 2011, 29: 7801-7806. DOI: 10.1016/j.vaccine.2011.07.104.
[12]Gurgel RG, Bohland AK, Vieira SC, et al. Incidence of rotavirus and all-cause diarrhea in northeast Brazil following the introduction of a national vaccination program[J]. Gastroenterology, 2009, 137: 1970-1975. DOI: 10.1053/j.gastro.2009.07.046
[13]Yee EL, Fang ZY, Liu N, et al. Importance and challenges of accurately counting rotavirus deaths in China, 2002[J]. Vaccine, 2009, 27(5): F46-F49. DOI: 10.1016/j.vaccine.2009.08.065
[14]Li M. Status of rotavirus infection in children with diarrhea in Nanjing Children’s Hospital[J]. J Appl Clin Pediatr, 2007, 22(19): 1463-1464. (in Chinese)
李梅. 南京兒童醫院腹瀉患兒輪狀病毒感染狀況[J]. 實用兒科臨床雜志, 2007, 22(19):1463-1464.
[15]Lin Q, Zhou JS, Lu F, et al. Study on clinical characteristics and molecular epidemiology of younger than 5 years old children with diarrhea caused by rotavirus infection in Nanjing city in 2009-2010[J]. J Appl Clin Pediatr, 2011, 26(22): 1709-1711. (in Chinese)
林謙, 周進蘇, 陸芬, 等. 2009-2010年南京地區5歲以下兒童輪狀病毒性腹瀉臨床特點及分子流行病學研究[J]. 實用兒科臨床雜志, 2011, 26(22):1709-1711.
[16]Kargar M, Najafi A, Zandi K, et al. Genotypic distribution of rotavirus strains causing severe gastroenteritis in children under 5 years old in Borazjan, Iran[J]. African J Microbiol Res, 2011, 5(19): 2936-2941.
[17]Zeng M, Chen J, Gong ST, et al. Epidemiological surveillance of norovirus and rotavirus diarrhea among outpatient children in five metropolitan cities[J]. Chin J Pediatr, 2010, 48(8): 564-570. (in Chinese)
曾玫, 陳潔, 龔四堂, 等. 我國五所城市兒童醫院諾如病毒和輪狀病毒腹瀉的流行病學監測[J]. 中華兒科雜志, 2010, 48(8): 564-570.
[19]Parashar U, Bresee J, Glass R. Rotavirus. Severe childhood diarrhea[J]. Emerg Infect Dis, 2006, 12: 304-306.
[20]Wang YH, Kobayashi N, Zhou DJ, et al. Molecular epidemiologic analysis of group A rotaviruses in adults and children with diarrhea in Wuhan city, China, 2000-2006[J]. Arch Virol, 2007, 152: 669-685.
[21]Wang YH, Zhou X, Ghosh S, et al. Prevalence of human rotavirus genotypes in Wuhan, China, during 2008 2011: Changing trend of predominant genotypes and emergence of strains with the P[8]b subtype of the VP4 gene[J]. Arch Virol, 2011, 156: 2221-2231. DOI: 10.1007/s00705-011-1122-9
[22]Clark HF, Hoshino Y, Bell LM, et al. Rotavirus isolate WI61 representing a presumptive new human serotype[J]. J Clin Microbiol, 1987, 25(9): 1757-1762.
[23]Sánchez-Fauquier A, Montero V, Moreno S, et al. Human rotavirus G9 and G3 as major cause of diarrhea in hospitalized children, Spain[J]. Emerg Infect Dis, 2006, 12: 1536-1541.
[24]Khamrin P, Peerakome S, Wangsawasdi L, et al. Emergence of human G9 rotavirus with an exceptionally high frequency in children admitted to hospital with diarrhea in Chiang Mai, Thailand[J]. J Med Virol, 2006, 78: 273-280.
[25]Qian Y, Yuan LJ, Xiong CH, et al. Identification of rotavirus G9 type from a stool specimen collected from a child with diarrhea in Beijing[J]. Chin J Virol, 1994, 10(3): 263-267.
[26]Yang XL, Matthijnssens J, Sun H, et al. Temporal changes of rotavirus strain distribution in a city in the northwest of China, 1996-2005[J]. Int J Infect Dis, 2008, 12(6): e11-17. DOI: 10.1016/j.ijid.2008.03.022
[27]Duan ZJ, Liu N, Yang SH, et al. Hospital-based surveillance of rotavirus diarrhea in the People’s Republic of China, August 2003-July 2007[J]. J Infect Dis, 2009, 200(Suppl 1): S167-173. DOI: 10.1086/605039
[28]Li DD, Yu QL, Qi SX, et al. Study on the epidemiological of rotavirus diarrhea in Lulong in 2008--2009[J]. Chin J Exp Clin Virol, 2010, 24(1): 2-4. (in Chinese)
李丹地,于秋麗,齊順祥,等.河北省盧龍地區2008-2009年度輪狀病毒流行病學研究[J].中華實驗和臨床病毒學雜志, 2010, 24(1):2-4.
[29]Zhang Q, Wang DK, Ye XH, et al. Molecular epidemiological research on rotavirus serotype G9 isolated from 9 regions in China[J]. Chin J Vaccines Immunization, 2006, 12(6): 476-479. (in Chinese)
章青, 王端可, 葉新華, 等. 中國1998-2004年G9型輪狀病毒分子流行病學研究[J]. 中國計劃免疫, 2006, 12(6):476:479.
[30]Hu L, Crawford SE, Czako R, et al. Cell attachment protein VP8* of a human rotavirus specifically interacts with A-type histo-blood group antigen[J]. Nature, 2012, 485(7397): 256-259. DOI: 10.1038/nature10996
[31]Cho MK, Jheong WH, Lee SG, et al. Full genomic analysis of a human rotavirus G1P[8]strain isolated in South Korea[J]. J Med Virol, 2013, 85(1): 157-170. DOI: 10.1002/jmv.23366
[32]Arora R, Chitambar SD. Full genomic analysis of Indian G1P[8]rotavirus strains[J]. Infect Genet Evol, 2011, 1(2): 504-511. DOI: 10.1016/j.meegid.2011.01.005
[33]Shintani T, Ghosh S, Wang YH, et al. Whole genomic analysis of human G1P[8]rotavirus strains from different age groups in China[J]. Viruses,2012, 4(8): 1289-1304.
猜你喜歡
幸福家庭(2023年8期)2023-08-17 10:06:50
紡織標準與質量(2022年1期)2022-07-12 06:01:12
家庭醫學(下半月)(2020年2期)2020-05-11 02:07:32
華人時刊(2019年23期)2019-05-21 03:31:38
健康管理(2017年4期)2017-05-20 08:17:06
媽媽寶寶(2017年4期)2017-02-25 07:00:48
媽媽寶寶(2017年2期)2017-02-21 01:21:04
中國當代醫藥(2015年20期)2015-03-01 02:04:41
中國醫療美容(2015年5期)2015-02-03 03:02:00
中國中醫藥現代遠程教育(2014年17期)2014-03-01 04:29:21
