日本血吸蟲重組Bb(pGEX-Sj26GST-Sj32)疫苗免疫BALB/c鼠血清抗體及細胞因子的動態觀察
2014-04-09 00:51:34向進平李文桂
中國人獸共患病學報 2014年3期
向進平,李文桂,張 麗
日本血吸蟲病是衛生部規劃防治的重點寄生蟲病之一,研制疫苗防治該病成為當前研究的熱點領域。人們發現血清抗體和細胞因子在疫苗誘導的保護性免疫機制中起重要作用。本文在成功構建重組Bb(pGEX-Sj26GST-Sj32)疫苗的基礎上,將其免疫BALB/c小鼠,動態觀察血清抗體及細胞因子,為研究該疫苗的作用機制提供材料。
1 材料與方法
1.1實驗動物 10~12周齡BALB/c鼠96只,雌性,15~20 g,購自重慶醫科大學實驗動物中心。
1.2主要試劑及器材 羊抗小鼠IgG、IgG1、IgG2a、IgG2b、IgG3、IgE和IgA均購自美國Southern Biotech公司;IL-10、IL-12試劑盒購自深圳欣博盛生物科技有限公司;其它試劑為國產分析純;SjAWA和rBb(pGEX-Sj26GST-Sj32)疫苗由本實驗室制備及保存;Elx800UV酶標儀購自美國Bio-tek公司。
1.3免疫方案 將96只BALB/c鼠隨機分為口服灌胃(per os,PO)組、鼻腔粘膜(intranasal,IN)組兩組,每組各48只。PO組給予108CFU/100 μL單次口服灌胃,IN組給予105CFU/10 μL單次鼻腔粘膜接種。至實驗結束時無小鼠死亡。
1.4血清收集:兩組小鼠分別于免疫后0、2、4、6、8、10、12、14、16、18、20和22周各隨機剖殺4只,取眼球血,室溫靜置6 h,5 000 r/min 7 min離心(離心半徑9.5 cm),分離血清,并于-20 ℃冰箱保存備用。
1.5血清IgG及其亞類、IgE和IgA抗體檢測 采用常規ELISA法。方陣滴定法確定SjAWA(1 μg/μL)的包被液濃度為2.4 μg/孔,小鼠血清的稀釋度為1∶50,酶標抗體的稀釋度為1∶5 000。酶標儀測定OD450值。
1.6血清IL-10及IL-12水平檢測 采用雙抗夾心ELISA法。按照IL-10和IL-12試劑盒的說明書進行操作。測OD450值;制作標準曲線,通過其OD值即可計算出各對應的細胞因子濃度,以pg/ml為單位(IL-10與IL-12檢測步驟相同)。
1.7統計學分析 采用SPSS 17.0對實驗數據進行分析,每周每組四份標本,多組間比較用單因素方差分析,組間兩兩比用Dunnett’s T3法,P<0.05有統計學意義。
2 結 果
2.1血清抗體檢測 口服免疫組小鼠血清中IgG、IgG1、IgG2a、IgG2b、IgG3、IgE和IgA水平分別在免疫后4~22周、8~22周、4~22周、2~8周、2~14周、8~16周和4~22周升高,并分別在6、8、8、4、6、12和6周達到峰值,此時與0周相比差異均有統計學意義(P<0.05);鼻腔粘膜接種組小鼠血清中IgG、IgG1、IgG2a、IgG2b、IgG3、IgE和IgA水平分別在免疫后4~22周、2~16周、2~20周、2~8周、4~10周、2~18周和2~22周升高,并分別在4、2、6、2、4、8和8周達到峰值,此時與0周相比差異均有統計學意義(P<0.05)。且鼻腔粘膜組各抗體水平均較口服免疫組先達峰值(P<0.05)。(見表1、圖1及圖2)
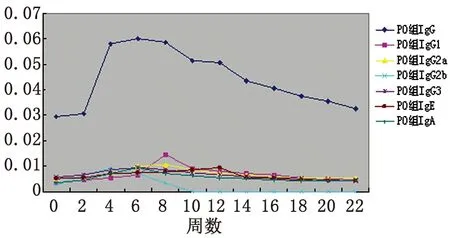
圖1rBb(pGEX-Sj26GST-Sj32)疫苗口服免疫小鼠血清抗體水平的動態變化
Fig.1DynamicchangesonthelevelsofantibodiesinseraofBALB/cmiceorallyimmunizedwithrBb(pGEX-Sj26GST-Sj32)vaccine
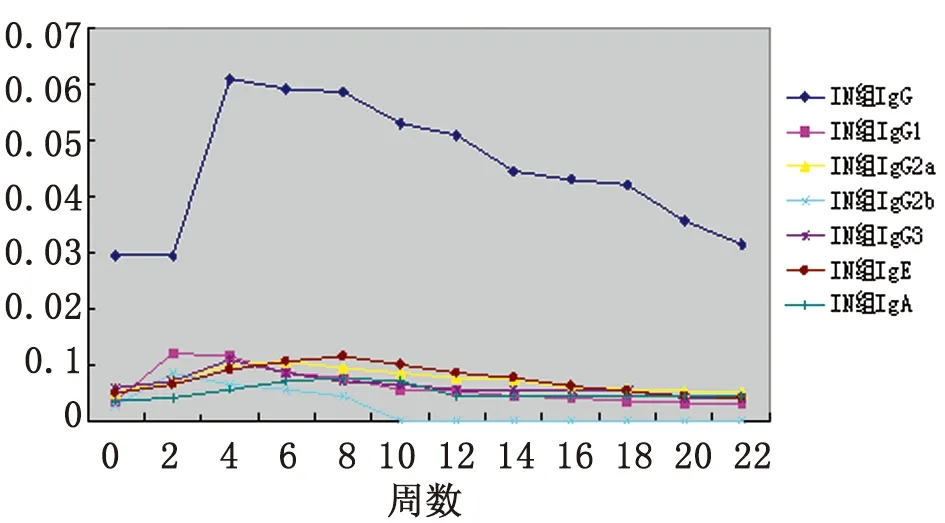
圖2rBb(pGEX-Sj26GST-Sj32)疫苗鼻腔粘膜免疫小鼠血清抗體水平的動態變化
Fig.2DynamicchangesonthelevelsofantibodiesinseraofBALB/cmiceintranasallyimmunizedwithrBb(pGEX-Sj26GST-Sj32)vaccine
2.2血清IL-10及 IL-12水平檢測 與0周相比,口服免疫組血清IL-10水平于2~22周顯著升高,并于12周達到峰值(P<0.05);鼻腔粘膜免疫組亦于2~22周顯著升高,且于6周達峰值(P<0.05)。口服組及鼻腔粘膜組IL-12水平均于2~14周升高,且均于6周達到最大值(P<0.05)(表2,圖3)。
3 討 論
抗血吸蟲感染過程中,IgG是抗感染的主要抗體,它具有調理作用,可增強吞噬細胞的吞噬作用;gG2a、IgG2b及IgE與保護力呈正相關,尤其是血吸蟲抗原特異性IgE抗體依賴的、嗜酸性粒細胞等細胞介導的細胞毒效應;IgG1及IgG2a也參與細胞毒效應,從而對靶細胞產生非特異性殺傷作用[1]。抗體依賴細胞介導的細胞毒作用(ADCC)殺傷血吸蟲童蟲被認為是抗血吸蟲感染中重要的免疫效應機制之一。Widhe等[2]發現IgG3可能通過IFN-γ激活效應細胞發揮殺傷血吸蟲作用。
表1rBb(pGEX-Sj26GST-Sj32)疫苗免疫小鼠后血清抗體的動態觀察
Tab.1DynamicobservationonthelevelsofantibodiesinseraofBALB/cmiceimmunizedwithrBb(pGEX-Sj26GST-Sj32)vaccine

WeekGroupIgGIgG1IgG2aIgG2bIgG3IgEIgA0PO0.030±0.0020.004±0.0010.005±0.0010.003±0.0010.006±0.0010.005±0.0010.004±0.001IN0.030±0.0020.004±0.0010.005±0.0010.003±0.0010.006±0.0010.005±0.0010.004±0.0012PO0.031±0.0020.005±0.0020.006±0.0010.007±0.003?0.007±0.0010.006±0.0010.005±0.001IN0.030±0.0010.012±0.001??0.007±0.0010.009±0.001??0.007±0.0010.007±0.0010.004±0.0014PO0.058±0.001?0.006±0.0010.007±0.001?0.009±0.001?0.009±0.001?0.007±0.0010.007±0.001?IN0.061±0.003??0.012±0.001??0.010±0.001?0.007±0.001??0.011±0.001?0.009±0.0010.006±0.0016PO0.060±0.002?0.007±0.002?0.01±0.001?0.007±0.001?0.010±0.001?0.008±0.0010.010±0.002?IN0.059±0.002??0.009±0.001??0.011±0.001?0.006±0.001??0.009±0.001?0.011±0.0020.007±0.0018PO0.059±0.001?0.015±0.003?0.011±0.002?0.004±0.0010.009±0.001?0.008±0.001?0.007±0.001?IN0.059±0.004??0.008±0.001??0.010±0.001?0.005±0.0010.007±0.0010.012±0.002?0.008±0.001??10PO0.052±0.001?0.009±0.001?0.009±0.001?00.008±0.0010.009±0.001?0.006±0.001IN0.053±0.001??0.006±0.0010.009±0.00100.007±0.0010.010±0.001?0.007±0.00112PO0.051±0.001?0.008±0.001?0.007±0.00100.007±0.0010.010±0.001?0.006±0.001IN0.051±0.001??0.005±0.0010.008±0.00100.006±0.0010.009±0.0010.005±0.00114PO0.044±0.002?0.007±0.001?0.007±0.00100.006±0.0010.006±0.0010.005±0.001IN0.045±0.003??0.005±0.0010.007±0.00100.006±0.0010.008±0.0010.005±0.00116PO0.041±0.002?0.007±0.001?0.006±0.00100.006±0.0010.005±0.0010.005±0.001IN0.043±0.002??0.004±0.0010.006±0.00100.006±0.0010.006±0.0010.005±0.00118PO0.038±0.0030.006±0.0020.006±0.00100.005±0.0010.005±0.0010.004±0.001IN0.042±0.002??0.004±0.0010.006±0.00100.005±0.0010.005±0.00050.005±0.00120PO0.036±0.0030.005±0.0020.006±0.00100.005±0.0010.004±0.0010.004±0.001IN0.036±0.0010.003±0.0010.005±0.00100.004±0.0010.005±0.0010.005±0.00122PO0.033±0.0030.005±0.0020.006±0.00200.004±0.0010.004±0.0010.004±0.001IN0.032±0.0020.003±0.0010.005±0.00100.004±0.0010.004±0.0010.005±0.001
Note: Compared with 0 week,*P<0.05, **P<0.01.
表2rBb(pGEX-Sj26GST-Sj32)疫苗免疫小鼠后血清IL-10及IL-12水平的動態觀察
Tab.2DynamicobservationonthelevelsofIL-10andIL-12inseraofBALB/cmiceimmunizedwithrBb(pGEX-Sj26GST-Sj32)vaccine
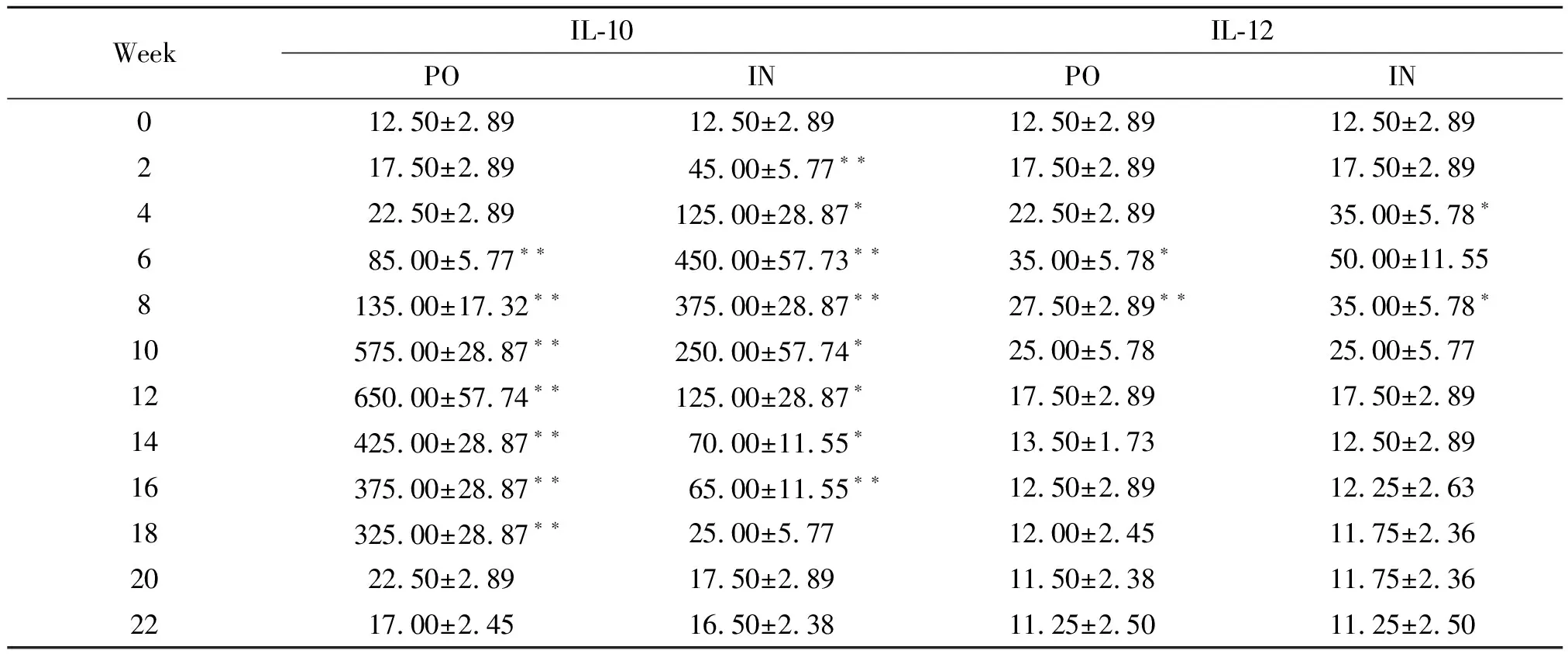
WeekIL-10IL-12PO IN PO IN012.50±2.8912.50±2.8912.50±2.8912.50±2.89217.50±2.8945.00±5.77??17.50±2.8917.50±2.89422.50±2.89125.00±28.87?22.50±2.8935.00±5.78?685.00±5.77??450.00±57.73??35.00±5.78?50.00±11.558135.00±17.32??375.00±28.87??27.50±2.89??35.00±5.78?10575.00±28.87??250.00±57.74?25.00±5.7825.00±5.7712650.00±57.74??125.00±28.87?17.50±2.8917.50±2.8914425.00±28.87??70.00±11.55?13.50±1.7312.50±2.8916375.00±28.87??65.00±11.55??12.50±2.8912.25±2.6318325.00±28.87??25.00±5.7712.00±2.4511.75±2.362022.50±2.8917.50±2.8911.50±2.3811.75±2.362217.00±2.4516.50±2.3811.25±2.5011.25±2.50
Note: Compared with 0 week,*P<0.05, **P<0.01
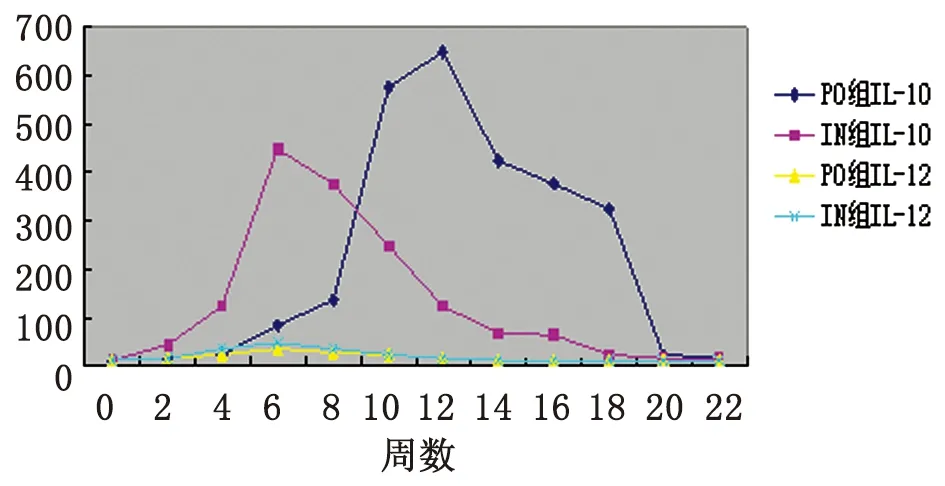
圖3rBb(pGEX-Sj26GST-Sj32)疫苗免疫小鼠后血清IL-10及IL-12水平的動態變化
Fig.3DynamicchangesonthelevelsofIL-10andIL-12inseraofBALB/cmiceimmunizedwithrBb(pGEX-Sj26GST-Sj32)vaccine
周霞等[3]將BALB/c小鼠經紫外減毒尾蚴免疫后,血清中針對成蟲抗原(SWAP)的IgG水平感染組小鼠上升較免疫組小鼠快,并于第5周達峰值。而免疫組小鼠血清IgG水平于第6周達峰值,且后者的峰值略高于前者。提示BALB/c鼠經免疫可使機體產生較強針對SWAP的抗體。龔唯等[4]將rSjc26GST加活化劑免疫BALB/c鼠后經皮膚感染尾蚴,發現用AWA檢測免疫小鼠血清IgG及IgG1水平均顯著高于對照組,能明顯增強免疫小鼠的體液免疫應答,使肝內血吸蟲卵肉芽腫病變顯著減輕,成熟蟲卵比例明顯下降。蔣守富等[5]將日本血吸蟲攻擊感染東方田鼠1~4周后,產生針對SAWA的IgG3抗體,抗體水平在攻擊感染后第3周增幅較大,第4周達高峰,較感染前增加49.6%;感染昆明種小鼠后,抗SAWA的IgG3抗體水平在攻擊感染后第3周增加36.7%;并發現該抗體主要針對Sj童蟲和成蟲。Wynn等[6]用曼氏血吸蟲(Sm)減毒活疫苗加IL-12多次免疫小鼠,攻擊感染后發現IgG2a 、IgG2b和 IgG1同時升高。李永等[7]采用非糖蛋白31/32 kDa抗原檢測慢性血吸蟲病人血清中IgG1、IgG3、IgG4各亞類均明顯增高,而IgG1高出正常對照組11.9倍。該結果與Jassim的報道基本一致[8]。Dupré等[9]用曼氏血吸蟲28 kDa谷胱甘肽-S-轉移酶的cDNA構建的DNA疫苗pNlp28于皮內注射免疫大鼠后,對血清抗體IgG及其亞類進行了觀察。結果表明,血清抗體可持續至第126 d(未繼續觀察),抗體亞類檢測表明主要是IgG2a和IgG2b。Etard等[10]認為血吸蟲感染免疫力的表達可能是“效應免疫應答” (主要是抗血吸蟲成蟲表面抗原,抗童蟲表面抗原的IgA、IgG1、IgG2a、IgG3和IgE)與“阻斷免疫應答”(主要是抗蟲卵多糖抗原的IgM、IgG2c和IgG4 )之間動態平衡的結果。
本研究發現,口服免疫組小鼠血清中IgG、IgG1、IgG2a、IgG2b和IgG3水平分別在免疫后4~22周、8~22周、4~22周、2~8周和2~14周升高,并分別在6、8、8、4和6周達到峰值(P<0.05);鼻腔粘膜接種組小鼠血清中IgG、IgG1、IgG2a、IgG2b和IgG3水平分別在免疫后4~22周、2~16周、2~20周、2~8周和4~10周升高,并分別在4、2、6、2和4周達到峰值(P<0.05)。且鼻腔粘膜組各抗體水平均較口服免疫組先達峰值(P<0.05)。提示重組Bb(pGEX-Sj26GST-Sj32)疫苗在早期就能產生有效的免疫應答,從而獲得較好的保護性免疫力。IgG水平顯著升高,說明其在抗日本血吸蟲感染過程中發揮抗感染作用,而IgG亞類則輔助其發揮免疫功能。且鼻腔粘膜免疫較口服免疫更早出現特異性抗體,提示粘膜接種途徑可有效誘導全身性免疫反應,因而免疫效率更高。
流行病學調查資料顯示,血吸蟲病流行區居民存在年齡依賴的與血吸蟲特異性IgE抗體相關的對再感染的抗力,提示血吸蟲特異性IgE抗體在血吸蟲病抗感染免疫中起到重要作用[11-12]。研究表明,日本血吸蟲血清IgE抗體水平與血吸蟲再感染和感染度顯著負相關。有報道指出,血吸蟲26GST免疫BALB/c小鼠能引起特異性IgE抗體的產生[13]。同樣,IgA在抗血吸蟲感染中也起著重要作用。Hernandez等[14]發現蠕蟲成蟲抗原IgA為保護性抗體,并可能在抗成蟲生殖力和蟲卵活性方面發揮重要作用。本課題組曾用日本血吸蟲重組Bb(pGEX-Sj4-3-3)疫苗以口服灌胃及鼻腔粘膜接種方式免疫BALB/c小鼠,發現兩種途徑均可誘導較高水平的IgE和IgA抗體,從而提高宿主抗血吸蟲感染的保護性免疫反應[15]。
本實驗中口服組及鼻腔粘膜組在疫苗免疫后IgE及IgA抗體水平均升高,口服組分別于8周~16周和4周~22周升高,并分別于12周和6周達到峰值(差異有統計學意義,P<0.05);鼻腔粘膜組分別于2周~18周和2周~22周升高,并均于8周達到峰值(P<0.05)。提示該重組Bb疫苗能引起抗日本血吸蟲感染免疫力,并在對抗成蟲生殖力、抗蟲卵活性及防止再感染方面起到一定作用。相比于口服組,鼻腔粘膜組能更早產生相應特異性抗體,減少蟲體及蟲卵對機體的損害。
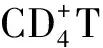
對血吸蟲輻照致弱尾蚴誘導的早期免疫應答的研究發現,IL-10是一種重要的負向調控因子,在IL-12介導的早期獲得性免疫應答調節中起著重要作用,包括減輕過度的皮膚炎癥和有效阻止高度極化Th1型應答的發生[19]。Hoffmann等認為 IL-10雖然被視為Th2效應因子,但研究發現IL-10對Th1和Th2反應都有重要的調控作用,影響疾病的發展和轉歸[20]。另外,人們對IL-12的生物學活性、功能、基因結構以及與疫苗相關性研究發現,IL-12在寄生蟲感染中具有保護性免疫作用。IL-12主要由單核-巨噬細胞、B細胞及肥大細胞產生, IL-12對T細胞和NK細胞具有顯著的生物學作用,能促進NK細胞活化、增殖,增強NK細胞和細胞毒T細胞(CTL)的殺傷活性,誘導活化的T細胞和NK細胞分泌IFN-γ,從而間接地激活巨噬細胞,協同IL-2誘導NK細胞活性的產生[21];同時IL-12使巨噬細胞增多,增加NO殺童蟲能力[22],從而增強宿主抗日本血吸蟲感染的保護性免疫力。曼氏血吸蟲感染早期主要是Thl為主的細胞免疫,IL-2、IL-12和IFN-γ水平升高,臨床表現為急性期反應;產卵后宿主體內的免疫反應由Th1型轉變為Th2型為主的體液免疫,以 IL-4、IL-5及IL-10等細胞因子升高為特點[23],在肉芽腫形成和纖維化過程中起著重要作用[24]。
參考文獻:
[1]Capron M, Capron A. Immunoglobulin E and effector cells in schistosomiasis[J]. Science, 1994, 264(5167): 1876-1877.
[2]Widhe M, Ekerfelt C, Forsberg P, et al. IgG subclasses in Lyme borreliosis: a study of specific IgG subclass distribution in an interferon-gamma-predominated disease[J]. Scand J Immunol, 1998, 47(6): 575-581.
[3]Zhou X, Zhuge HX, Gong W, et al. Dynamic changes of IgG levels and passive transfer research with sera from BALB/c mice vaccinated with attenuated cercariae ofSchistosomajaponicum[J]. Chin J Zoonoses, 2008, 24(5): 483-484. (in Chinese)
周霞,諸葛洪祥,龔唯,等.減毒日本血吸蟲尾蚴免疫BALB/c小鼠血清IgG水平動態及其被動轉移保護力研究[J].中國人獸共患病學報,2008,24(5):483-484.
[4]Gong W, Luo W, Liu SX, et al. Effect of rSjc26GST on anti-Schistosomajaponicumhepatic egg granuloma reinforced by activator HYP and its mechanism[J]. Suzhou Univ J Med Sci, 2005, 25(1): 1-3. (in Chinese)
龔唯,駱偉,劉述先,等.活化劑 HYP協同rSjc26GST的抗血吸蟲性肉芽腫的效果及其機制的研究[J].蘇州大學學報,2005,25(1):1-3(37).
[5]Jiang SF, Wei MX, Lin JJ, et al. Effect of IgG3 antibody purified from sera ofMicrotusfortisagainstSchistosomajaponicum[J]. Chin J Parasitol Parasit Dis, 2008, 26(1): 34-36. (in Chinese)
蔣守富,魏梅雄,林矯矯,等.東方田鼠IgG3抗體抗血吸蟲病作用研究[J].中國寄生蟲學與寄生蟲病雜志,2008,26(1):34-36.
[6]Wynn TA, Reynolds A, James S, et al. IL-12 enhances vaccine-induced immunity to schistosomes by augmenting both humoral and cell-mediated immune responses against the parasite[J]. J Immunol, 1996, 157(9): 4068-4078.
[7]Li Y, Zeng XF, Yi XY, et al. Detection of different class (subclass) antibodies in sera of patients with schistosomiasis japonica for diagnosis and efficiency evaluation[J]. Bull Hunan Med Univ, 1997, 22(5): 381-383. (in Chinese)
李永,曾憲芳,易新元,等.應用日本血吸蟲病患者血清特異性抗體亞類考核療效[J].湖南醫科大學學報,1997,22(5):381-383.
[8]Jassim A, Hassen K, Carty D. Antibody subclass in human schistosomiasis[J]. Parasite Immunol, 1987, 9(6): 627-650.
[9]Dupré L, Poulain-Godefroy O, Ban E, et al. Intradermal immunization of rats with plasmid DNA encodingSchistosomamansoni28 kDa glutathione S-transferase[J]. Parasite Immunol, 1997, 19(11): 505-513.
[10]Etard JF, Audibert M, Dabo A, et al. Age-acquired resistance and predisposition to reinfection withSchistosomahaematobiumafter treament with praziquantel in Mali[J]. Am J Trop Med Hyg, 1995, 52(6): 549-558.
[11]Dunne DW, Webster M, Smith P, et al. The isolation of a 22 kDa band after SDS-PAGE ofSchistosomamansoniadult worms and its use to demonstrate that IgE responses against the antigen(s) it contains are associated with human resistance to reinfection[J]. Parasite Immunol, 1997, 19(2): 79-89.
[12]Caldas IR, Correa-Oliveira R, Colosimo E, et al. Susceptibility and resistance toSchistosomamansonireinfection: parallel cellular and isotypic immunologic assessment[J]. Am J Trop Med Hyg, 2000, 62(1): 57-64.
[13]Hsu CH, Chua KY, Huang SK, et al. Glutathione-S-transferase induces murine dermatitis that resembles human atopic dermatitis[J]. Clin Exp Allergy, 1996, 26(11): 1329-1337.
[14]Hernandez MG, Hafalla JC, Acosta LP, et al. Paramyosin is a major target of the human IgA response againstSchistosomajaponicum[J]. Parasite Immunol, 1999, 21(12): 641-647.
[15]Zhang N, Li WG. Dynamic observation on levels of antibodies in sera of BALB/c mice immunized with recombinantBifidobacteriumbifidumpGEX-Sj14-3-3 vaccine ofSchistosomajaponicum[J]. Chin J Endemiol, 2012, 31(3): 301-304. (in Chinese)
張寧,李文桂.日本血吸蟲重組兩歧雙歧桿菌pGEX-Sj14-3-3疫苗免疫BALB/c小鼠的血清抗體動態觀察[J].中國地方病學雜志,2012,31(3):301-304.
[16]Jankovic D, Wynn TA, Kullberg MC, et al. Optimal vaccination againstSchistosomamansonirequires the induction of both B cell- and IFN-gamma-dependent effector mechanisms[J]. J Immunol, 1999, 162(1): 345-351.
[17]Oswald IP, Caspar P, Jankovic D, et al. IL-12 inhibits Th2 cytokine responses induced by eggs ofSchistosomamansoni[J]. J Immunol, 1994, 153(4): 1707-1713.
[18]Xia CM, Luo W, Gong W, et al. Effects of varied immunogen on Thl/Th2 immune deviation of mice infected withSchistosomajaponicum[J]. Chin J Zoonoses, 2002, 18(1): 55-58. (in Chinese)
夏超明,駱偉,龔唯,等.日本血吸蟲不同免疫原對小鼠Th1/Th2免疫偏移的影響[J].中國人獸共患病雜志,2002,18(1):55-58.
[19]Qadir K, Metwali A, Blum AM, et al. TGF-beta and IL-10 regulation of IFN-gamma produced in Th2-type schistosome granulomas requires IL-12[J]. Am J Physiol Gastrointest Liver Physiol, 2001, 281(4): G940-946.
[20]Hoffmann KF, Cheever AW, Wynn TA. IL-10 and the dangers of immune polarization: excessive type 1 and type 2 cytokine responses induce distinct forms of lethal immunopathology in murine schistosomasis[J]. J Immunol, 2000, 164(12): 6406-6416.
[21]Scott P. IL-12: initiation cytokine for cell-mediated immunity[J]. Science, 1993, 260(5107): 496-497.
[22]Wynn TA, Oswald IP, Eltoum IA, et al. Elevated expression of Th1 cytokines and nitric oxide synthase in the lungs of vaccinated mice after challenge infection withSchistosomamansoni[J]. J Immunol, 1994, 153(11): 5200-5209.
[23]Pearce EJ, Kane CM, Sun J, et al. Th2 response polarization during infection with the helminth parasiteSchistosomamansoni[J]. Immunol Rev, 2004, 201: 117-126.
[24]Li XY, Shen JL. TH1/TH2 immune response and schistosomal fibrosis[J]. Chin J Zoonoses, 2009, 25(2): 179-182. (in Chinese)
李小月,沈繼龍.TH1/TH2免疫應答與血吸蟲性纖維化[J].中國人獸共患病學報,2009,25(2):179-182.
[25]Li Y, Gong W, Zhang HQ, et al. Reorganization and identification of fractional antigens from adult worms and eggs by sera of rabbits infected withSchistosomajaponicumin different stages of disease[J]. Chin J Zoonoses, 2008, 24(6): 551-554. (in Chinese)
李穎,龔唯,張惠琴,等.日本血吸蟲不同病期血清抗體對蟲源性及卵源性組分抗原的識別及鑒定[J].中國人獸共患病學報,2008,24(6):551-554.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2019年12期)2019-05-21 02:55:32
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
