O1群霍亂弧菌PFGE分子分型和菌株流行能力關(guān)系研究
2014-04-09 00:51:34陳愛平楊勁松李曲文徐海濱王靈嵐鄭金鳳嚴(yán)延生
中國人獸共患病學(xué)報(bào) 2014年3期
關(guān)鍵詞:優(yōu)勢
陳愛平,楊勁松,李曲文,徐海濱,王靈嵐,鄭金鳳,嚴(yán)延生
福建省自1962年發(fā)生埃爾托型霍亂弧菌(EVC)引起的霍亂以來,疫情起伏不定,先后發(fā)生了幾次較大流行。首次為1962-1964年,流行優(yōu)勢血清型為O1群小川;第2次為1978-1989年,流行優(yōu)勢血清型先為1978年O1群小川型,后轉(zhuǎn)為稻葉型,流行時(shí)間持續(xù)近10年;第3次1994-2000年,流行優(yōu)勢血清型為O1群小川;2005年我省又發(fā)生了O1群稻葉的霍亂流行,疫情僅限于2005年;在福建省50年的霍亂流行過程中病原和流行特征都發(fā)生著改變。為闡明福建省不同時(shí)期、地域、不同血清群霍亂分離菌株間的遺傳聯(lián)系和遺傳多樣性,我們選擇福建省1962-2005年代表性霍亂弧菌,運(yùn)用脈沖場凝膠電泳(PFGE)技術(shù)進(jìn)行分子分型,從細(xì)菌全基因組水平探索霍亂弧菌在福建省傳播及流行規(guī)律;同時(shí)結(jié)合流行病學(xué)資料,根據(jù)疫情流行持續(xù)時(shí)間、相關(guān)疫情病例數(shù)等,分析不同PFGE型(簇)的霍亂菌株的流行能力。
1 材料方法
1.1實(shí)驗(yàn)用菌株 選擇1962-2005年福建省歷次霍亂流行,病人中分離保存的菌種77株;包括3次大流行的菌株(1962-1964年、 1978-1989年, 1994-2000年)、2005年菌株和2001-2003年散發(fā)病例的菌株;實(shí)驗(yàn)用菌株均經(jīng)過血清學(xué)重新復(fù)核鑒定為O1群霍亂弧菌。菌株信息見圖1。
1.2PFGE實(shí)驗(yàn)方法 根據(jù) PulseNet USA(美國CDC)網(wǎng)絡(luò)實(shí)驗(yàn)室推薦的標(biāo)準(zhǔn)方法,經(jīng)國家CDC實(shí)驗(yàn)優(yōu)化條件的基礎(chǔ)上進(jìn)行PFGE實(shí)驗(yàn)[1]。采用NotI限制性內(nèi)切酶對(duì)實(shí)驗(yàn)菌株進(jìn)行酶切,標(biāo)準(zhǔn)菌株H9812采用XbaI限制性內(nèi)切酶消化,主要的電泳參數(shù)為電壓梯度6 V/cm,電泳夾角120°,電泳溫度14 ℃;電泳參數(shù):2 s~10 s,13 h, 20 s~25 s ,6 h(ctxA基因陰性菌株在電泳液中加硫脲,終濃度為50 μmol/L)。
1.3數(shù)據(jù)分析 PFGE圖像導(dǎo)入BioNumerics(version 6.0,Applied Maths,Inc)軟件包進(jìn)行處理,選擇Dice相關(guān)系數(shù)和UPGMA方法,tolerance設(shè)置為1.5%。
1.4試劑儀器 Seakem Gold Agarose PFGE 級(jí)瓊脂糖購自Lonza公司,XbaⅠ和NotⅠ限制性內(nèi)切酶以及蛋白酶K購自美國NEB公司,BIO-RAD公司的Basic電泳儀、Gel Doc XR+讀膠儀、CHEF MAPPER脈沖場凝膠電泳儀,BioMerieux Vitek公司的DENSIMAT細(xì)菌濁度儀。
2 結(jié) 果
2.177株霍亂弧菌PFGE分子分型結(jié)果 77株1962-2005年霍亂弧菌用NotⅠ酶切并進(jìn)行脈沖場電泳后,DNA片段可得到較好地分離,可見大小不一的電泳條帶,分子量從20~690 kb;菌株條帶數(shù)目在21~26之間。利用Bionumerics軟件的Cluster analysis模塊進(jìn)行聚類分析,菌株間條帶相似度在75.3%~100%之間;按照100%的相似度可將菌株酶切圖譜分為61個(gè)PFGE型,其中P1型有6株,P13、P14和P57型各有3株,其余各型為1-2株。根據(jù)TENOVER原則[2],只有1-3條帶差異的菌株在遺傳上可能有緊密相關(guān)性,按照90%的相似度將部分菌株酶切圖譜劃為同一個(gè)簇,可得到5個(gè)主要的PFGE簇(G1-G5)。見圖1。

圖177株霍亂弧菌PFGE分型結(jié)果
Fig.1NotI-digestedPFGEpatternsof77V.choleraeO1serogroupisolates
2.277株霍亂弧菌PFGE簇不同流行時(shí)期分布 第1次流行的菌株以G5 PFGE簇為主(6/9),同時(shí)存在G4和G1 PFGE簇;第2次流行的菌株以G1和G4 PFGE簇為主(18/24,6/24),第3次流行的菌株以G3和G4 PFGE簇為主(11/30,10/30),2005年流行的菌株以G2為主(6/6),2001-2003年散發(fā)的菌株沒有優(yōu)勢的PFGE簇,具體見表1。
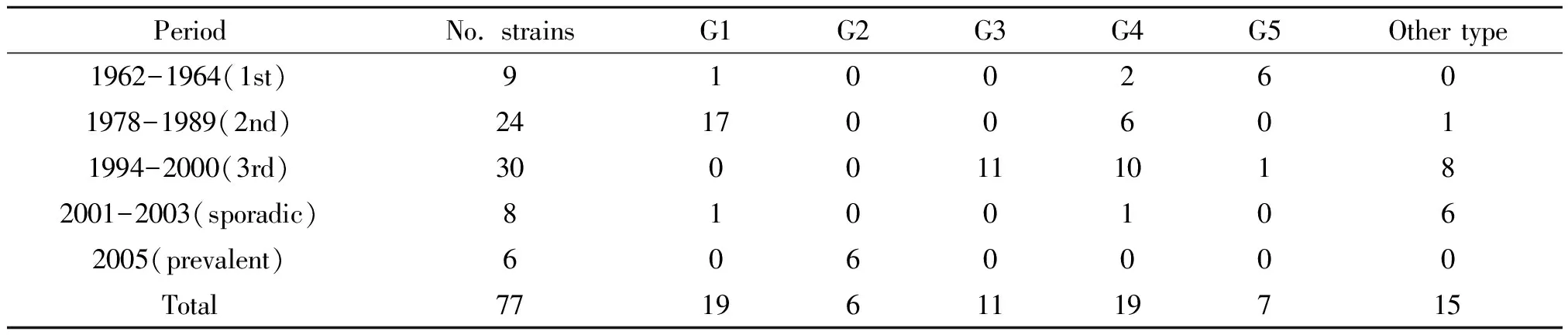
表1 77株1962-2005年霍亂弧菌PFGE 簇不同流行時(shí)期分布情況
2.352株小川血清型PFGE分子分型結(jié)果 52株小川型霍亂弧菌,按照100%的相似度可將菌株酶切圖譜分為47個(gè)PFGE型。按照90%的相似度將部分菌株酶切圖譜劃為同一個(gè)簇,可得到4個(gè)主要的PFGE簇,命名為G1(o)-G4(o),o是小川型(Ogawa)的第一個(gè)字母;具體見圖2。
2.4小川型PFGE 簇不同流行時(shí)期的分布情況 1962-1964年流行的菌株以G4(o)PFGE簇為主(7/9);1978年流行的菌株以G3(o)PFGE簇為主(6/8);1994-2000年流行的菌株以G2(o)PFGE簇為主(14/29);1994-2000年屬于其它型別的小川型菌株有9/29;2001-2003散發(fā)的小川型菌株沒有優(yōu)勢PFGE簇;具體見表2。
2.5稻葉血清型PFGE分子分型結(jié)果 25株稻葉型霍亂弧菌按照100%的相似度可將菌株酶切圖譜分為15個(gè)PFGE型。按照90%的相似度將部分菌株酶切圖譜劃為同一個(gè)簇,可得到2個(gè)主要的PFGE簇,命名為G1(i)-G2(i),i是稻葉型(Inaba)的第一個(gè)字母;具體見圖3。1979-1989年的菌株屬于G1(i) PFGE簇(14/14),2005年的菌株屬于G2(i)PFGE簇(6/6);其余年份散發(fā)的稻葉型菌株沒有優(yōu)勢的PFGE簇;具體見圖3。

圖252株小川血清型PFGE分子分型結(jié)果
Fig.2PFGEpatternsof52V.choleraeOgawaserotypestrains
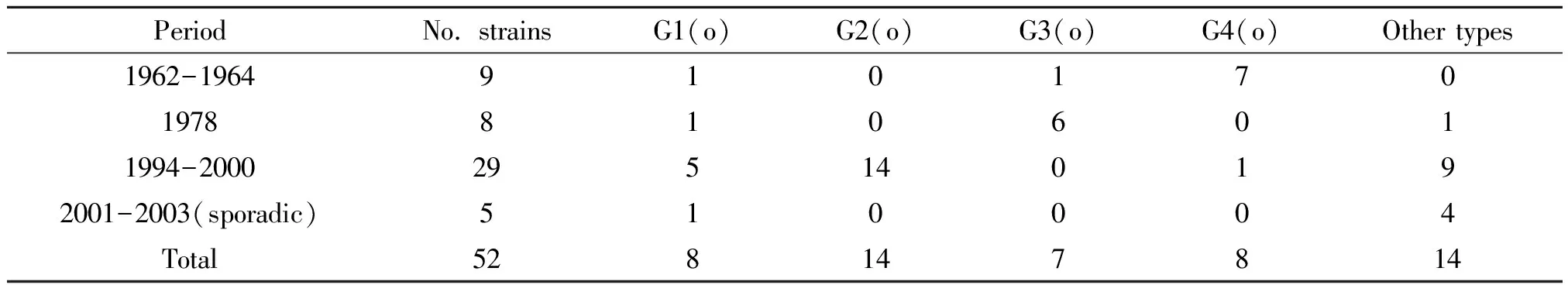
表2 52株小川型霍亂弧菌PFGE簇不同流行時(shí)期分布情況
3 討 論
隨著脈沖場凝膠電泳(PFGE)分子分型技術(shù)的標(biāo)準(zhǔn)化以及PulseNet監(jiān)測網(wǎng)絡(luò)的建立,越來越多的公共實(shí)驗(yàn)室逐漸利用該技術(shù)平臺(tái)進(jìn)行各種細(xì)菌性病原體監(jiān)測。我們對(duì)1962-2005代表性的O1群霍亂病人分離株進(jìn)行PFGE分子分型,結(jié)果發(fā)現(xiàn),按照100%的相似度可將77株菌株酶切圖譜分為61個(gè)PFGE型,沒有集中優(yōu)勢的PFGE型別。PFGE分子分型針對(duì)的是全細(xì)菌的基因組,對(duì)于同一次暴發(fā)流行的菌株,理論上菌株的基因組應(yīng)該表現(xiàn)為100%的相似度;但是針對(duì)長時(shí)間跨年度流行的分離株以及實(shí)驗(yàn)室長期保存的菌株(特別是以保種半固體方式保存的霍亂菌株),因?yàn)榫暝陂L時(shí)間流行傳播和保存過程可能產(chǎn)生基因變異,即使是有遺傳相關(guān)性的菌株P(guān)FGE分型也不一定表現(xiàn)為100%的相似度;但是這種變異一般不會(huì)產(chǎn)生大的基因組變異。根據(jù)TENOVE原則[1],PFGE基因型別只有1-3條帶差異的菌株在遺傳上可能有緊密相關(guān)性,實(shí)驗(yàn)結(jié)果按照90%的相似度將大部分菌株酶切圖譜劃分為5個(gè)主要的PFGE簇G1-G5,實(shí)際上歸于同一個(gè)簇的菌株大部分是同一次流行的菌株;散發(fā)的菌株沒有顯示優(yōu)勢的簇,表現(xiàn)為各自不同的PFGE型別,提示散發(fā)菌株沒有遺傳上的相關(guān)性。因此對(duì)于PFGE分子分型的結(jié)果分析要結(jié)合收集菌株的流行病學(xué)背景資料、研究菌株的時(shí)間跨度和菌種的保存方式等選擇適當(dāng)?shù)南嗨贫冗M(jìn)行聚類,更能客觀、準(zhǔn)確地反映菌株之間的相關(guān)性。
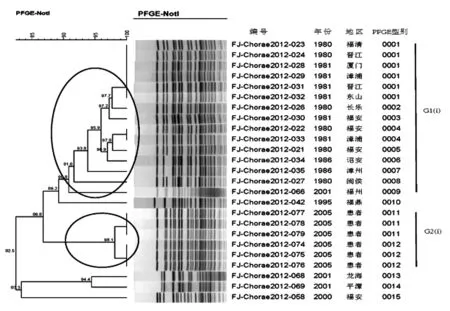
圖325株稻葉型PFGE分子分型結(jié)果
Fig.3PFGEpatternsof25V.choleraeInabaserotypestrains
從1962-2005年福建省霍亂菌株P(guān)FGE簇不同流行時(shí)期的分布情況可見,前3次流行的菌株既有上一次流行簇的延續(xù),又有新的優(yōu)勢簇的出現(xiàn);說明前3次流行的菌株在遺傳上有關(guān)聯(lián)性,但是在長時(shí)間流行過程中產(chǎn)生基因變異出現(xiàn)新的優(yōu)勢簇。相同的優(yōu)勢簇分布于福建省各個(gè)地區(qū),提示福建省各地區(qū)間的霍亂流行在傳播上有相關(guān)性。
福建以小川型為優(yōu)勢血清型的霍亂流行時(shí)期有1962-1964、1978和1994-2000年[1];各個(gè)流行時(shí)期均有各自優(yōu)勢的PFGE簇,分別為G4(o)、G3(o)、G2(o)。小川型為優(yōu)勢血清型的3次流行,均造成較多的病例數(shù)(均>1500例),但是從流行持續(xù)的時(shí)間來看1994-2000年明顯超過1962-1964年和1978年,推測G2(o) PFGE簇的小川型菌株在引起霍亂流行時(shí)間長度能力上要強(qiáng)于G3(o)和G4(o)簇。另外,1994-2000年小川型菌株比其它流行時(shí)期的小川型菌株基因組更具有多樣性,可能原因是由于該次流行的時(shí)間跨度達(dá)10年之久,在長時(shí)間的流行過程中菌株產(chǎn)生了基因變異,特別是到了流行后期;而實(shí)際上歸于其它PFGE型別的9株菌株,有8株是1998-2000年流行后期的菌株。
福建以稻葉為優(yōu)勢血清型的霍亂流行時(shí)期有1979-1989和2005年;1979-1989年稻葉型流行了將近10年的時(shí)間,總發(fā)病數(shù)為22292例,2005年稻葉型引發(fā)的疫情僅限于當(dāng)年,發(fā)病數(shù)310;從疫情的延續(xù)時(shí)間、發(fā)病規(guī)模和引發(fā)的病例數(shù)進(jìn)行比較,1979-1989年都明顯超過2005年;1979-1989年的菌株均歸于G1(i) PFGE簇,2005年的菌株均歸于G2(i) PFGE簇;可以推測G1(i) PFGE簇的稻葉型菌株具有引起長時(shí)間、大規(guī)模霍亂流行的能力;G2(i) PFGE簇的菌株在流行能力上明顯較弱。
本文將O1群霍亂弧菌PFGE分子分型的結(jié)果和流行病學(xué)的背景資料相結(jié)合,對(duì)不同PFGE簇(型)菌株在流行時(shí)間長短、引發(fā)流行規(guī)模的能力等方面,進(jìn)行初步探索,為評(píng)估將來霍亂疫情分離株對(duì)流行趨勢的可能影響積累實(shí)驗(yàn)室研究本底資料。
參考文獻(xiàn):
[1]Cooper KL, Luey CKY, Bird M, et al. Development and validation of a PulseNet standardized pulsed-field gel electrophoresis protocol for subtyping ofVibriocholerae[J]. Foodborne Pathog Dis, 2006, 3(1): 51-58. DOI: 10.1089/fpd.2006.3.51
[2]Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing[J]. J Clin Microbiol, 1995, 33(9): 2233-2239.
猜你喜歡
我愛學(xué)·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時(shí)刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛(wèi)生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
