利用種群增長研究鏡湖夏季萼花臂尾輪蟲對水溫的適應
2014-03-29 02:01:12席貽龍
水生生物學報 2014年6期
王 敏 席貽龍 周 彬 張 燁
(安徽師范大學生命科學學院, 安徽省高校生物環境與生態安全省級重點實驗室, 蕪湖 241000)
利用種群增長研究鏡湖夏季萼花臂尾輪蟲對水溫的適應
王 敏 席貽龍 周 彬 張 燁
(安徽師范大學生命科學學院, 安徽省高校生物環境與生態安全省級重點實驗室, 蕪湖 241000)
運用種群增長實驗方法, 在4個溫度(20℃、24℃、28℃和32 )℃和3個斜生柵藻[Scenedesmus obliquus (Turpin) Kuetzing]密度(1.0×106、2.0×106和4.0×106cells/mL)共12個條件組合下研究了鏡湖夏季出現的萼花臂尾輪蟲(Brachionus calyciflorus Pallas)的種群增長率、平均混交率、平均受精率和休眠卵產量等生殖參數。結果表明, 在各食物密度下, 輪蟲種群增長率均在 28℃和 32℃下最高。平均混交率、平均受精率和休眠卵產量等與溫度間的關系因食物密度的不同而異。溫度對種群增長率、平均混交率、平均受精率和休眠卵產量均有極顯著性影響(P<0.01), 食物密度對種群增長率和休眠卵產量均有極顯著性影響(P<0.01), 溫度和食物密度的交互作用對平均混交率、平均受精率和休眠卵產量均有極顯著性影響(P<0.01)。在1.0×106、2.0×106和4.0×106cells/mL的斜生柵藻密度以及28℃和32℃下, 輪蟲具有較高的種群增長率(1.50—1.95/d)表明該種群能夠適應較高的水溫, 水溫不是導致其在夏季水體中具有極低的種群密度(遠低于1個/L)的直接因素。在種群存在于鏡湖水體期間(7月初至12月初), 特別是當水溫降至20℃左右時, 其較高的休眠卵產量為該種群在次年的同一時段在水體中重新出現提供了保證。
萼花臂尾輪蟲; 斜生柵藻密度; 溫度; 生殖參數
輪蟲是各類水體中廣泛分布的一類浮游動物,在水生態系統的結構組成、物質循環和能量流動過程中具有重要的作用。由于水環境中的物理 、化學和生物因子等普遍具有隨著時間的流逝或季節的更替而發生變化的特點, 棲息于其中的輪蟲種群密度也隨之發生變化。為探明自然水體中輪蟲種群數量的時空變動及其調節機制, 眾多學者作出了不懈地努力。溫新利[1]曾以蕪湖市汀棠湖中的萼花臂尾輪蟲等為對象, 研究了其種群數量的周年變動, 探討了水溫、Chl.a濃度、捕食者密度等外源性因素對種群數量周年變動的影響; 但尚未涉及輪蟲種群對特定環境的適應性等內源性因素。王愛民等[2]利用生命表統計學方法研究了汀棠湖冬季出現的萼花臂尾輪蟲對水溫的適應, 發現其種群在16℃下的凈生殖率和種群內稟增長率均最高, 推測這或許是該水體中萼花臂尾輪蟲種群密度在3月中旬(此時水溫為17 )℃達到春季最高峰的重要原因之一。種群增長率通常被認為是種群的短期適合度指標, 它代表種群在特定環境下的增殖能力[3]; 休眠卵產量是種群的長期適合度指標, 因為休眠卵是種群度過不利環境條件的唯一途徑[4,5], 因此, 通過研究種群增長率和休眠卵產量等可以揭示種群對特定環境條件的適應,幫助我們理解種群變動的內源性機制。
鏡湖為一亞熱帶小型淺水湖泊, 平均水深為1.3 m, 水域面積約0.079 km2, 處于富營養化初期階段。鏡湖主要為公園景觀水域, 水體中有魚類的分布; 但常年缺乏枝角類和橈足類, 且大型浮游植物也偏少[6]。該湖位于蕪湖市中心, 市區年平均溫度約16 , ℃ 最高溫度為41 , ℃ 最低溫度為–15℃。在湖泊內, 萼花臂尾輪蟲主要出現于冬春季, 而夏秋季幾近消失(種群密度遠低于1個/L)[1]。夏季, 水溫的波動范圍在28—33℃。為了解鏡湖夏季出現的萼花臂尾輪蟲對水溫的適應性, 探討溫度對其種群在夏季幾近消失的可能作用, 本文研究了溫度和食物密度對鏡湖夏季水體中出現的萼花臂尾輪蟲生殖參數的影響。
1 材料與方法
1.1 輪蟲的采集與培養
實驗用萼花臂尾輪蟲于2011年7月3日—8月13日用25號浮游生物網自蕪湖市鏡湖水體中大范圍拖撈獲得。在此期間, 水溫的變動范圍為28—33℃。采樣后, 實驗室內解剖鏡下隨機挑取萼花臂尾輪蟲非混交雌體, 于(25±1)℃、自然光照[光照強度約1300 μmol/(m2·s)]條件下進行克隆培養。培養液為新鮮配制的輪蟲培養液[7], 食物為HB-4培養基[8]培養的、處于指數增長期的斜生柵藻, 投喂密度為2.0×106cells/mL。藻培養期間, 每天添加20%的HB-4培養液以保持其處于指數增長期; 使用前, 經4500 r/min離心7min濃縮, 并采用浮游植物計數框于顯微鏡下計數。從最終建立的輪蟲克隆中隨機選取4個克隆用于本實驗研究。
1.2 輪蟲的生殖參數研究
實驗參照夏季自然水溫的波動范圍, 將溫度設計為20℃、24℃、28℃和32℃等4個溫度。同時, 考慮到與水溫變化密切相關的藻類等食物所產生的上行效應也可能是影響輪蟲種群密度季節變動的重要因素, 而處于輕度富營養狀態的鏡湖水體中藻類和有機碎屑等輪蟲食物較為豐富[1], 將食物密度設計為1.0×106、2.0×106和4.0×106cells/mL等3個斜生柵藻密度。在實驗前, 將每個輪蟲克隆分別置于20℃、24℃、28℃和32℃等4個溫度以及1.0×106、2.0×106和4.0×106cells/mL等3個斜生柵藻密度下進行為期一周的預培養。在預培養時, 以直徑為1 cm、長度為12 cm的玻璃試管為容器, 培養體積為5 mL。在預培養過程中, 每天更換輪蟲培養液并投喂藻類食物一次, 同時通過去除一部分個體使得輪蟲種群始終處于指數增長期。
在實驗開始時, 從每個克隆中挑取 100個以上攜卵的非混交雌體置于試管中繼續培養。4h后, 從各試管中挑取齡長在 4h以內的幼體于玻璃杯中繼續培養, 待其產出第一枚非混交卵時接種至玻璃試管內開始實驗, 接種密度為 2 ind./mL, 培養液體積為5 mL。在實驗過程中, 每12h輕輕吹浮沉積于試管底部的藻類食物; 每 24h對培養液中輪蟲各類型雌體計數1次, 并投喂餌料。待種群密度較大時, 采用抽樣計數, 用玻璃微吸管將培養液輕輕攪勻后,從試管的3個不同部位于水體中部吸取(0.5—2) mL的培養物進行計數。雌體類型的劃分和鑒定與席貽龍和黃祥飛[9]相同。
1.3 相關參數的定義和計算方法
種群增長率: r=(lnNt–lnN0)/t, N0為起始輪蟲密度, 為2 ind./mL; Nt為試驗進行到t天時的輪蟲種群密度; t為試驗進行的天數。根據Dumont等[10]的方法, 從輪蟲種群的指數增長期選取4—6個數據計算種群增長率并最終獲得每個實驗重復的平均值。
混交雌體百分率: 種群中的混交雌體數占總雌體數的百分比, 本文使用的是試驗期間每日混交雌體百分率的平均值; 混交雌體受精率: 種群中產休眠卵的混交雌體數占總混交雌體數的百分比, 本文使用的是試驗期間每日混交雌體受精率的平均值;休眠卵產量: 5 mL培養液中的輪蟲在4d內所產的休眠卵數量。
1.4 數據的統計分析
采用SPSS 16.0統計分析軟件對上述數據進行統計分析。對所得的各生殖參數進行正態分布檢驗后, 對符合正態分布的數據進行單因素和雙因素方差分析以揭示溫度和食物密度對輪蟲各生殖參數的影響, 應用 LSD檢驗法對同一溫度下各食物密度間、同一食物密度下各溫度間有統計學意義(P<0.05)的輪蟲各生殖參數的均值間作多重比較。
2 結果
由圖1可見, 在各溫度下, 輪蟲總雌體密度的最大值隨著培養溫度的升高而增大; 產雄卵的混交雌體密度的最大值也隨著培養溫度的升高而呈現增大的趨勢, 24℃除外; 在20℃和28℃下, 產休眠卵的混交雌體密度的最大值也隨著培養溫度的升高而呈現增大的趨勢。在各食物密度下, 輪蟲總雌體密度的最大值出現在 28℃; 產雄卵的混交雌體密度的最大值出現在 32℃; 產休眠卵的混交雌體密度的最大值出現在20℃, 1.0×106cells/mL食物密度下除外。
單因素方差分析表明, 在1.0×106cells/mL食物密度下, 溫度對輪蟲種群增長率、平均混交率和平均受精率均有極顯著的影響(P<0.01), 但對休眠卵產量無顯著影響(P>0.05); 在 2.0×106和 4.0×106cells/mL食物密度下, 溫度對輪蟲種群增長率、平均混交率、平均受精率和休眠卵產量均有極顯著的影響(P<0.01)。多重比較顯示, 在各食物密度下, 輪蟲種群增長率隨著溫度的升高而增大, 但28℃和32℃間無顯著的差異。在1.0×106cells/mL食物密度下, 平均混交率在20℃和24℃顯著低于32 , ℃ 但28℃的平均混交率與其他三溫度間無顯著的差異; 平均受精率在24℃和28℃最高, 32℃最低。在2.0×106cells/mL食物密度下, 平均混交率在20℃最高, 24℃顯著低于32 , ℃ 而28℃的平均混交率與24℃和32℃間均無顯著的差異; 平均受精率在 20℃、24℃和 28℃較高, 32℃較低; 休眠卵產量在 20℃較高, 其他三個溫度下較低。在 4.0×106cells/mL食物密度下, 平均混交率在 20℃最高, 其他三個溫度間均無顯著的差異;平均受精率在20℃和24℃最高, 32℃最低; 休眠卵產量在20℃較高, 其他三個溫度下較低(表1)。
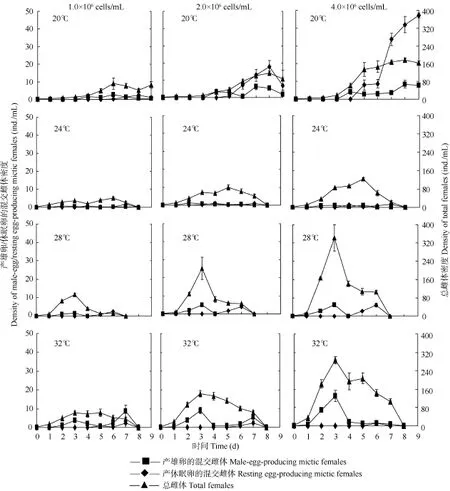
圖1 溫度和食物密度對鏡湖夏季萼花臂尾輪蟲種群中產雄卵的混交雌體密度、產休眠卵的混交雌體密度和總雌體密度的影響Fig. 1 The effects of four temperatures and three food concentrations on the density of male-egg-producing mictic females, resting egg-producing mictic females and total females in B. calyciflorus populations from Lake Jinghu
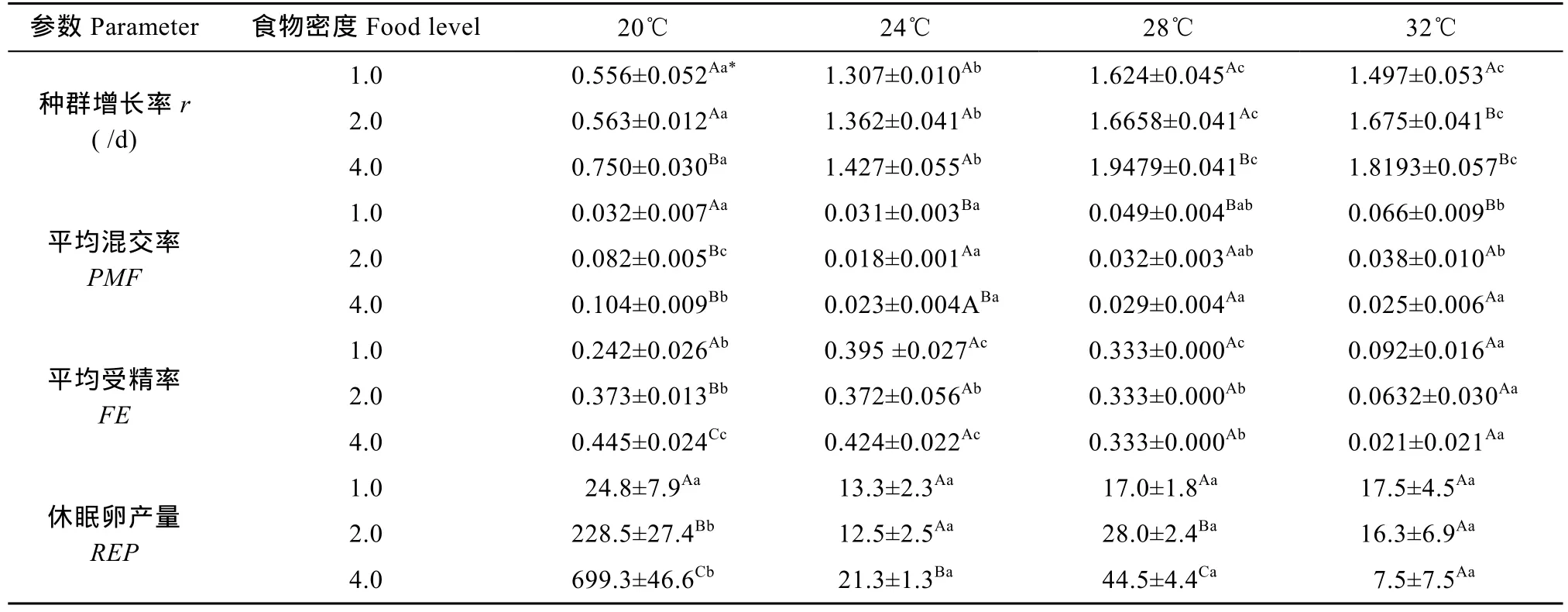
表1 四個溫度和三個食物密度(×106cells/mL)下萼花臂尾輪蟲的種群增長率、平均混交率、平均受精率和休眠卵產量(均數±標準誤)Tab. 1 Population growth rate (r), average proportion of mictic females (PMF), average fertilization rate of mictic females (FR), and resting egg production (REP) of B. calyciflorus at four temperatures and three food levels (Mean±SE)
在 20℃下, 食物密度對種群增長率、平均混交率、平均受精率和休眠卵產量有極顯著影響(P<0.01);在 24℃下, 食物密度對種群增長率和平均受精率無顯著影響(P>0.05), 而對平均混交率和休眠卵產量有顯著影響(P<0.05); 在 28℃下, 食物密度對種群增長率、平均混交率和休眠卵產量有極顯著影響(P<0.01), 而對平均受精率無顯著的影響(P>0.05);在 32℃下, 食物密度對種群增長率和平均混交率均有顯著影響(P<0.05), 但對平均受精率和休眠卵產量無顯著影響(P>0.05)。多重比較顯示, 在 20℃下,輪蟲種群增長率在1.0×106和2.0×106cells/mL食物密度下較低, 4.0×106cells/mL食物密度下較高; 平均混交率在 1.0×106cells/mL 食物密度下較低, 2.0×106和4.0×106cells/mL食物密度下較高; 平均受精率和休眠卵產量均隨著食物密度的升高而升高。在24℃下, 平均混交率在1.0×106cells/mL食物密度下顯著高于 2.0×106cells/mL食物密度下, 而 4.0× 106cells/mL食物密度下的平均混交率與 1.0×106和2.0×106cells/mL食物密度間均無顯著的差異; 休眠卵產量在1.0×106和2.0×106cells/mL食物密度下較低, 4.0×106cells/mL食物密度下較高。在28℃下, 輪蟲種群增長率在1.0×106和2.0×106cells/mL食物密度下較低, 4.0×106cells/mL食物密度下較高; 平均混交率在1.0×106cells/mL食物密度下較高, 2.0×106和 4.0×106cells/mL食物密度下較低; 休眠卵產量隨著食物密度的升高而升高。在 32℃下, 輪蟲種群增長率在1.0×106cells/mL較低, 2.0×106和4.0×106cells/mL食物密度下較高; 平均混交率在1.0×106cells/mL食物密度下較高, 2.0×106和4.0×106cells/mL食物密度下較低(表1)。
雙因素方差分析表明, 溫度對種群增長率、平均混交率、平均受精率均和休眠卵產量均有極顯著性影響(P<0.01), 食物密度對群增長率和休眠卵產量均有極顯著性影響(P<0.01), 溫度和食物密度的交互作用對平均混交率、平均受精率和休眠卵產量均有極顯著性影響(P<0.01)(表2)。
3 討 論
3.1 輪蟲種群對水溫的適應
已有研究結果表明, 輪蟲種群增長率對升高的溫度的反應因輪蟲種類的不同而異。哈瓦那臂尾輪蟲(B. havanaensis Ehrenberg)、壺狀臂尾輪蟲(B. urceolaris Müller)、紅臂尾輪蟲(B. rubens Ehrenberg)、剪形臂尾輪蟲(B. forcatus Wierzejski)和尾突臂尾輪蟲(B. caudatus Barrois et Daday)的種群內稟增長率均隨著溫度的升高而顯著升高[11—13]; 但萼花臂尾輪蟲種復合體內姐妹種LE9的種群內稟增長率在28℃下最高, 姐妹種HE1的種群內稟增長率在23℃下趨于最高, 姐妹種HE3的種群內稟增長率在23℃和 28℃下最高[14]; 熱帶龜甲輪蟲(Keratella tropica Apstein)的種群內稟增長率在 30℃下最高,曲腿龜甲輪蟲(Keratella valga Ehrenberg)的種群內稟增長率在25℃下最高[15]。在1.0×106、3.0×106和5.0×106cells/mL斜生柵藻密度下, 溫度對裂足臂尾輪蟲的種群內稟增長率均無顯著性影響[16]; 在本研究中, 在1.0×106、2.0×106和4.0×106cells/mL斜生柵藻密度下, 鏡湖夏季出現的萼花臂尾輪蟲種群增長率均在28℃和32℃下最高, 表明該種群適應于較高的水溫; 食物密度的升高并不明顯地改變其對較高的水溫的適應性。
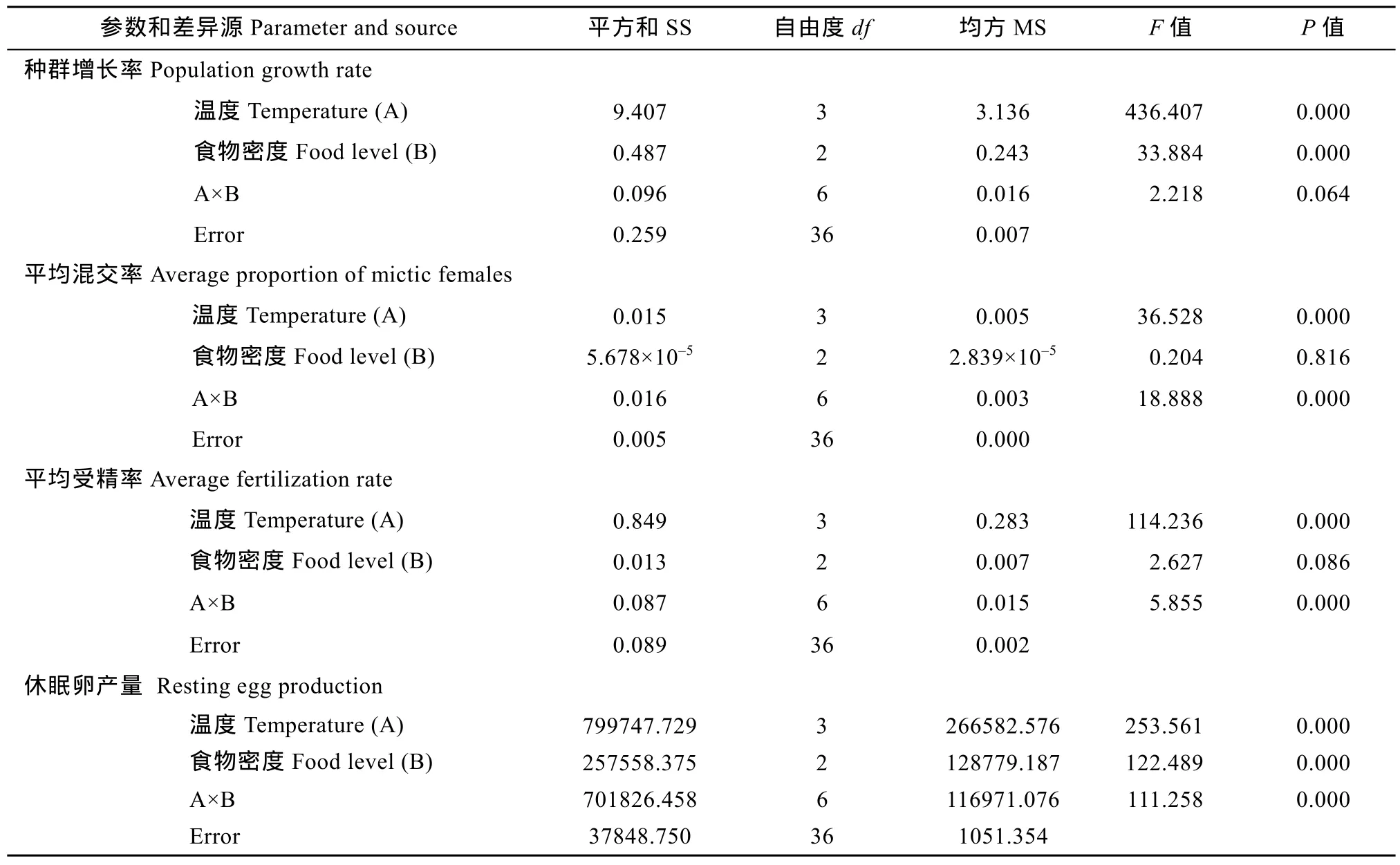
表2 溫度和食物密度對萼花臂尾輪蟲種群增長率、平均混交率、平均受精率和休眠卵產量的影響(雙因素方差分析)Tab. 2 Results of two-way Analysis of variance (ANOVA) performed for population growth rate, average proportion of mictic females and average fertilization rate of mictic females, and resting egg production of B. calyciflorus at four temperatures and three food levels
3.2 輪蟲有性生殖對水溫變化的響應
混交雌體的產生是輪蟲有性生殖的開始, 休眠卵的形成是有性生殖的終結。已有研究結果表明,褶皺臂尾輪蟲(B. plicatilis Müller)和萼花臂尾輪蟲種群中的混交雌體百分率隨著溫度的升高而降低[17—20]; 但青島、蕪湖和廣州品系的萼花臂尾輪蟲種群中的混交雌體百分率不受溫度的顯著性影響[21];在1.0×106、3.0×106和5.0×106cells/mL斜生柵藻密度下, 裂足臂尾輪蟲的后代混交率也均不受溫度的顯著性影響[16]。本研究結果表明, 萼花臂尾輪蟲種群中的混交雌體百分率受溫度和食物密度的交互作用。
已有研究結果表明, 溫度是影響輪蟲休眠卵產量的重要外源性因素, 而遺傳因素則是重要的內源性因素。溫度對休眠卵產量的影響常因輪蟲種類、品系或克隆的不同而異[17,21—24]。本研究結果表明,鏡湖夏季出現的萼花臂尾輪蟲休眠卵產量不僅受溫度的顯著性影響, 還受食物密度的顯著性影響以及溫度和食物密度的顯著性交互作用; 輪蟲在低溫(20 )℃ 和高食物密度(4.0×106cells/mL)下具有較高的休眠卵產量, 對種群度過不利的環境條件具有重要的意義。溫新利[1]研究發現, 該種群在鏡湖水體中的存在時間為7月初至12月初; 在此階段, 水溫的變動范圍為 32—12 ; ℃ 如此長的時間范圍內, 特別是當水溫降至 20℃左右時, 其較高的休眠卵產量是該種群在次年的同一時段在水體中重新出現的唯一途徑。
3.3 影響輪蟲種群密度季節變化的因素
種群數量動態及其調節機制始終是輪蟲種群生態學研究的核心問題之一。Liu等[25]對2006年3月鏡湖水體中出現的萼花臂尾輪蟲種群密度及其生殖參數進行研究時發現, 3月28日采自鏡湖的萼花臂尾輪蟲在接近當時的湖水溫度(19 )℃下進行培養時,其種群中的混交雌體百分率高達 55.8%; 當培養溫度升高至 25℃時, 種群中的混交雌體百分率達77.2%; 作者推測, 這或許是3月末鏡湖萼花臂尾輪蟲種群密度急劇下降、4月初種群幾乎從湖泊中消失的重要原因。Wen等[6]對鏡湖角突臂尾輪蟲(B. angularis Gosse)野外種群動態及其影響因素的研究發現, 水溫是控制夏-秋季節間該種輪蟲種群動態的關鍵因素, 10月初其種群逐漸消失的主要原因在于溫度誘導的較高的混交雌體百分率。王愛民等[2]利用生命表統計學方法研究了汀棠湖冬季出現的萼花臂尾輪蟲對水溫的適應, 發現其種群在 16℃下的凈生殖率和種群內稟增長率均最高, 推測這或許是該湖泊中萼花臂尾輪蟲種群密度在3月中旬(此時水溫為 17 )℃達到春季最高峰的重要原因之一。至于 4月初(此時水溫為 18 )℃該種輪蟲的種群密度降至較低水平的原因, 則與在此之前水溫等環境因子的劇烈波動導致此時輪蟲種群中的混交雌體百分率較高(37%)有關[1]。在本研究中, 在 1.0×106、2.0×106和4.0×106cells/mL斜生柵藻密度以及28℃和32℃下,輪蟲具有較高的種群增長率(1.50—1.95/d)表明該種群能夠適應較高的水溫, 水溫不是導致其在夏季水體中極低的種群密度(遠低于1個/L)的直接因素。
[1] Wen X L. Population dynamics and regulation mechanisms of Brachionus calyciflorus and Brachionus angularis (Rotifera) in two shallow eutrophic lakes [D]. Ph.D dissertation, Anhui Normal University, Wuhu, 2011 [溫新利.淺水富營養湖泊中兩種臂尾輪蟲的種群變動和機制研究.博士學位論文, 安徽師范大學. 蕪湖, 2011]
[2] Wang A M, Xi Y L, Niu X X, et al. Adaptation of Brachionus calyciflorus (Rotifera) collected in winter from Lake Tingtang to increasing water temperature in spring: a life table demography study [J]. Acta Ecologica Sinica, 2014, (16). http://dx.doi.org/10.5846/ stxb201212221842 [王愛民,席貽龍, 牛翔翔, 等. 汀棠湖冬季出現的萼花臂尾輪蟲對水溫的適應. 生態學報, 2014, (16). http://dx.doi.org/ 10.5846/stxb201212221842]
[3] Stelzer C P. Evolution of rotifer life histories [J]. Hydrobiologia, 2005, 546: 335—346
[4] Campillo S, García-Roger E M, Carmona M J, et al. Local adaptation in rotifer populations [J]. Evolutionary Ecology, 2010, 25: 933—947
[5] Xi Y L, Huang X F. Advances in studies on the ecological mechanism of the formation and hatching of resting eggs of rotifers [J]. Acta Hydrobiologica Sinica, 1999, 23(1): 73—82 [席貽龍, 黃祥飛. 輪蟲休眠卵形成和萌發的生態機理研究進展. 水生生物學報, 1999, 23(1): 73—82]
[6] Wen X L, Xi Y L, Yang Y F, et al. Temperature is the key factor controlling population dynamics of Brachionus angularis in Lake Jinghu during summer and autumn [J]. Journal of Freshwater Ecology, 2011, 26: 277—286
[7] Gilbert J J. Mictic female production in rotifer Brachionus calyciflorus [J]. Journal of Experimental Zoology, 1963, 153 (2): 113—124
[8] Zhang Z S, Huang X F. Method for Study on Freshwater Plankton [M]. Beijing: Science Press. 1991, 410—411 [章宗涉, 黃祥飛. 淡水浮游動物研究方法. 北京: 科學出版社. 1991, 410—411]
[9] Xi Y L, Huang X F. Effect of pH on population dynamics and resting egg of Brachionus calyciflorus [J]. Journal of Fishery Sciences of China, 1999, 6(3): 19—22 [席貽龍, 黃祥飛. pH對萼花臂尾輪蟲休眠卵形成的影響. 中國水產科學, 1999, 6(3): 19—22]
[10] Dumont H J, Sarma S S S, Ali A J. Laboratory studies on the population dynamics of Anuraeopsis fissa (Rotifera) in relation to food density [J]. Freshwater Biology, 1995, 33: 39—46
[11] Pavón-Meza E L, Sarma S S S, Nandini S. Combined effects of algal (Chlorella vulgaris) food level and temperature on the demography of Brachionus havanaensis (Rotifera): a life table study [J]. Hydrobiologia, 2005, 546: 353—360
[12] Hu C B, Xi Y L, Tao L X. Comparative study on the life history characteristics of Brachionus rubens and B. urceolaris [J]. Acta Ecologica Sinica, 2008, 28(12): 5957—5963 [胡存兵, 席貽龍, 陶李祥. 紅臂尾輪蟲和壺狀臂尾輪蟲生活史特征比較. 生態學報, 2008, 28(12): 5957—5963]
[13] Tao L X, Xi Y L, Hu C B. Comparative study on the life history characteristics of rotifer Brachionus forcatus and B. caudatus [J]. Chinese Journal of Applied Ecology, 2008, 19(5): 1104—1109 [陶李祥, 席貽龍, 胡存兵. 鐮形臂尾輪蟲和尾突臂尾輪蟲生活史特征的比較研究. 應用生態學報, 2008, 19(5): 1104—1109]
[14] Li H B, Xi Y L, Cheng X F. Comparative studies on life history characteristics of three sibling species in Brachionus calyciflorus species complex [J]. Acta Ecologica Sinica, 2009, 29(2): 581—588 [李化炳, 席貽龍, 程新峰. 萼花臂尾輪蟲種復合體內三個姐妹種生活史特征比較. 生態學報, 2009, 29(2): 581—588]
[15] Xi Y L, Xu D D, Ma J, et al. Differences in life table parameters between Keratella tropica and Keratella valga (Rotatoria) from subtropical shallow lakes [J]. Journal of Freshwater Ecology, 2013, 28(4): 539—545
[16] Ning L F, Xi Y L, Sun Q, et al. Combined effects of temperature and algal food density on life table demography of Brachionus diversicornis (Rotifera) [J]. Journal of Lake Sciences, 2013, 25(2): 295—301 [寧樂鋒, 席貽龍, 孫強,等. 溫度和食物密度對裂足臂尾輪蟲生命表統計學參數的影響. 湖泊科學, 2013, 25(2): 295—301]
[17] Hagiwara A, Hino A, Hirano R. Effect of temperature andchlorinity on resting egg formation in the rotifer Brachionus plicatilis [J]. Nippon Suisan Gakubu, 1988a, 54: 569—575
[18] Hagiwara A, Hino A, Hirano R. Comparison of resting egg formation among five Japanese stocks of the rotifer Brachionus plicatilis [J]. Nippon Suisan Gakubu, 1988b, 54: 577—580
[19] Hagiwara A, Lee C S. Resting egg formation of the L- and S-type rotifre Brachionus plicatilis under the different water temperature [J]. Nippon Suisan Gakubu, 1991, 57(9): 1645—1650
[20] Xi Y L, Huang X F. Effect of temperature and food concentration on resting egg fromation of freshwater rotifer Brachionus calyciflorus [J]. Acta Hydrobiologica Sinica, 2000, 24(2): 107—113 [席貽龍, 黃祥飛. 溫度和食物濃度對萼花臂尾輪蟲休眠卵形成的影響. 水生生物學報, 2000, 24(2): 107—113]
[21] Xi Y L, Dong L L, Ge Y L, et al. Resting egg production of Brachionus calyciflorus (Rotifera) at the different water temperatures [J]. Journal of Freshwater Ecology, 2004, 19: 567—573
[22] Snell T W, Hoff F H. The effect of environmental factors on resting egg production in rotifer Brachionus plicatilis [J]. Journal of World Mariculture Society, 1985, 16: 484—497
[23] Jiang D H, Xi Y L, Liu S G, et al. Seasonal variation of genetic structure and reproductive parameter of Brachionus calyciflorus (Rotifera) population in Lake Jinghu [J]. Acta Hydrobiologica Sinica, 2008, 32(6): 908—915 [江東海, 席貽龍, 劉勝國, 等. 鏡湖萼花臂尾輪蟲種群等位酶分析和生殖參數的季節變化. 水生生物學報, 2008, 32(6): 908—915]
[24] Ma Q, Xi Y L, Zhang J Y, et al. Differences in asexual and sexual reproduction among eight geographic populatons of Brachionus calyciflorus (Rotifera) from China [J]. Journal of Freshwater Ecology, 2009, 24: 273—278
[25] Liu S G, Xi Y L, Jiang D H. Temporal differences in reproductive characteristics of Brachionus calyciflrorus (Rotifera) from Lake Jinghu, China [J]. Journal of Freshwater Ecology, 2007, 22: 13—15
ADAPTATION OF BRACHIONUS CALYCIFLORUS (ROTIFERA) FROM LAKE JINGHU IN SUMMER TO WATER TEMPERATURE: POPULATION GROWTH EXPERIMENT STUDY
WANG Min, XI Yi-Long, ZHOU Bin and ZHANG Ye
(Provincial Key Laboratory of Biotic Environment and Ecological Safety, College of Life Sciences, Anhui Normal University, Wuhu 241000, China)
To study the population growth rate, average proportion of mictic females, average fertilization rate of mictic females, and resting egg production, Brachionus calyciflorus collected from Lake Jinghu were investigated at four temperatures (20℃, 24℃, 28℃and 32℃ ) and three Scenedesmus obliquus densities (1.0×106, 2.0×106 and 4.0×106 cells/mL) by population growth experiments. The results showed that the population growth rate of B. calyciflorus at 28℃ and 32℃ was the highest at all three food levels. At 1.0×106 cells/mL of S. obliquus, the average proportion of mictic females at both 20℃ and 24℃was significantly lower than that at 32℃, but it was not markedly different compared to that at 28℃. The average fertilization rate of mictic females at both 24℃ and 28℃ was the highest, and it was the lo west at 32℃. At 2.0×106 cells/mL of S. obliquus, the average proportion of mictic females at 20℃ was the highest. The average proportion of mictic females at 24℃ was lower than that at 32℃, but it was not significantly different compared to that at 28℃. The averagefertilization rate of mictic females at 20℃ , 24℃ and 28℃ was all higher than that at 32℃. The resting egg production at 20℃ was the biggest.At 4.0×106 cells/mL of S. obliquus, both the average proportion of mictic females and the resting egg production at 20℃ were higher than those at any other temperatures. The average fertilization rate of mictic females at both 20℃ and 24℃ was the highest, and it was the lowest at 32℃ . Two-way ANOVA analysis indicated that temperature significantly affected population growth rate, average proportion of mictic females, average fertilization rate of mictic females, and resting egg production (P<0.01), food level affected population growth rate and resting egg production (P<0.01), and temperature and food level cooperatively affected average proportion of mictic females and average fertilization rate of mictic females, and resting egg production (P<0.01). These results indicated that the rotifer population adapts high water temperature and water temperature does not directly account for the very low population density of B. calyciflorus in Lake Jinghu in summer, and that the high resting egg production of B. calyciflorus at low temperature may contribute to its reoccurrence in Lake Jinghu during the same period of next year.
Brachionus calyciflorus; Scenedesmus obliquus density; Temperature; Reproductive parameter
Q958
A
1000-3207(2014)06-1017-07
10.7541/2014.150
2013-09-04;
2014-03-12
國家自然科學基金(31170395); 教育部博士點基金(20093424110002); 安徽省高校生物環境與生態安全省級重點實驗室專項基金; 重要生物資源保護和利用研究安徽省重點實驗室專項基金資助
王敏(1989—), 男, 安徽省太和人; 碩士; 主要從事水生動物生態學研究。E-mail: wm321.happy@163.com
席貽龍, E-mail: ylxi1965@126.com
