基于mtDNA COⅠ和Cytb基因序列對南中國海不同海域波紋唇魚群體遺傳多樣性的研究
2014-03-29 02:01:10靜侯新遠(yuǎn)尹紹武祝斐賈一何王小軍
水生生物學(xué)報(bào) 2014年6期
關(guān)鍵詞:分析
胡 靜侯新遠(yuǎn)尹紹武祝 斐賈一何王小軍
(1. 南京師范大學(xué)生命科學(xué)學(xué)院, 南京 210023; 2. 中國水產(chǎn)科學(xué)院南海水產(chǎn)研究所熱帶水產(chǎn)研究中心, 三亞 572018)
基于mtDNA COⅠ和Cytb基因序列對南中國海不同海域波紋唇魚群體遺傳多樣性的研究
胡 靜1,2侯新遠(yuǎn)1尹紹武1祝 斐1賈一何1王小軍1
(1. 南京師范大學(xué)生命科學(xué)學(xué)院, 南京 210023; 2. 中國水產(chǎn)科學(xué)院南海水產(chǎn)研究所熱帶水產(chǎn)研究中心, 三亞 572018)
研究以海南陵水、馬來西亞、西沙、南沙4個(gè)海域共101尾波紋唇魚作為研究對象, 通過線粒體DNA的COⅠ和Cytb基因序列分析方法對波紋唇魚進(jìn)行了遺傳多樣性研究。經(jīng)PCR擴(kuò)增、克隆與序列測定, 分別獲得1560 bp COⅠ基因和1141 bp Cytb基因序列。兩者多態(tài)性遺傳參數(shù)統(tǒng)計(jì)顯示, 101尾個(gè)體分別存在23 (COⅠ)和30 (Cytb)個(gè)變異位點(diǎn), 分別檢測出20 (COⅠ)和27 (Cytb)個(gè)單倍型, 總?cè)后w單倍型多樣性(Hd)分別為0.629 (COⅠ)和0.755 (Cytb), 平均核苷酸差異數(shù)(K)分別為1.195 (COⅠ)和1.424 (Cytb), 核苷酸多樣性指數(shù)(Pi)分別為0.00077 (COⅠ)和0.00126 (Cytb)。分子方差分析(AMOVA)結(jié)果分別為26.26% (COⅠ)和4.22% (Cytb)的變異來自群體間, 73.74% (COⅠ)和95.78% (Cytb)的變異來自群體內(nèi)。同時(shí), 兩個(gè)基因的單倍型網(wǎng)絡(luò)圖呈星狀放射結(jié)構(gòu), 不同地理來源的單倍型無明顯分支, 呈交錯分布, 沒有體現(xiàn)地理差異性。研究初步認(rèn)為,波紋唇魚的遺傳多樣性處于較低水平, 遺傳分化存在但不顯著, 該結(jié)果可為今后波紋唇魚的種質(zhì)資源保護(hù)工作提供必要的科學(xué)依據(jù)。
波紋唇魚; CO ; CytⅠ b; 遺傳多樣性; 遺傳分化
波紋唇魚(Cheilinus undulatus)在分類學(xué)上隸屬鱸形目(Perciformes)、隆頭魚科(Labridae)、唇魚屬(Cheilinus),是大型的珊瑚礁魚類之一。在我國主要分布在海南的萬寧、陵水、三亞等海域, 以及香港、臺灣諸島近海域; 世界范圍內(nèi), 紅海、非洲東岸以及印度洋至太平洋中心較為多見。波紋唇魚肉質(zhì)好營養(yǎng)高, 市場需求量大。但由于濫捕濫殺、環(huán)境污染等因素, 自然海域波紋唇魚越來越少, 甚至瀕臨滅絕。因此, 有關(guān)波紋唇魚的資源保護(hù)、遺傳資源調(diào)查和人工繁殖等工作亟待加強(qiáng)。
本研究以海南陵水、馬來西亞、西沙以及南沙4個(gè)不同地理群體波紋唇魚共 101尾個(gè)體為研究對象, 對其COⅠ和Cytb基因序列進(jìn)行擴(kuò)增、克隆及測序, 目的在于分析波紋唇魚群體的 COⅠ和 Cytb基因序列變異和序列多態(tài)性情況, 研究其遺傳多樣性以及群體的遺傳關(guān)系, 為該物種保護(hù)學(xué)、遺傳學(xué)研究提供科學(xué)資料。
1 材料與方法
1.1 樣品采集
野生群體波紋唇魚分別取自海南陵水(LC)、馬來西亞(MC)、西沙(XC)以及南沙(NC)(表1), 各31、20、14、36尾,共101尾個(gè)體, 對其編號, 并用95 %乙醇保存?zhèn)溆谩?/p>

表1 波紋唇魚樣品的基本信息Tab. 1 Basic information of Cheilinus undulatus samples
1.2 DNA提取、擴(kuò)增、克隆及測序
實(shí)驗(yàn)材料為各個(gè)體0.1 g尾鰭組織。基因組DNA的提取方法為常規(guī)的“酚–氯仿”法[16]。DNA先經(jīng)冰乙醇沉淀, 后由 70%(體積分?jǐn)?shù))乙醇洗滌并干燥,最后 用50 μL TE溶解, –20℃保存?zhèn)溆谩?/p>
用于擴(kuò)增 COⅠ、Cytb基因的引物設(shè)計(jì)及合成步驟: 導(dǎo)出 Genbank中波紋唇魚的 mtDNA全序列(序列號: GU296101)、利用軟件Primer primer 5.0進(jìn)行設(shè)計(jì)、上海捷瑞生物工程有限公司合成。COⅠ基因序列的兩條引物分別為 F1 (5′CAGGATTTC AACCAGGAC 3′), R1 (5′TTTTGGACGTTAAGCTA CTG 3′); Cytb基因序列的兩條引物分別為 F2(5′GCCGACTACTTTGGATTT 3′), R2 (5′AGTTGTGAT GGATGTGCC 3′)。
兩個(gè) PCR反應(yīng)的總體積均為 25 μL: 2.5 μL 10×PCR Buffer, 2 μL dNTPs (2 mmol/L), 2 μL鎂離子(25 mmol/L), 0.3 μL TaqDNA 聚合酶 (5 U/μL), 正、反向引物各1 μL (10 μmol/L), 模板2 μL, 14.2 μL ddH2O。且PCR程序均為: 94℃預(yù)變性4min后進(jìn)行30個(gè)熱循環(huán)(94℃ 30s, 51℃ 30s, 72℃ 2min), 72℃延伸10min; 10℃保溫。
1%瓊脂糖凝膠電泳檢測純化后回收擴(kuò)增產(chǎn)物,將其與pMDTM18-T Vector*1 (寶生物工程有限公司)連接、與 DH5α感受態(tài)細(xì)胞充分混勻。液體培養(yǎng)基培養(yǎng)單克隆菌落, 后菌液 PCR, 產(chǎn)物送至上海華大基因公司測序。
(1)High speed in the analysis and design(reducing design time and cost).
1.3 數(shù)據(jù)處理與分析
雙向測序結(jié)果在BLAST上比對分析、人工完成完整序列拼接; ClustalX 軟件上對比文件輸入MEGA5.0軟件[17]分析依據(jù)Kimura 2-parameter計(jì)算群體內(nèi)及群體間的平均遺傳距離; 遺傳多樣性參數(shù),包括核苷酸多樣性 Pi以及單倍型多樣性 Hd由Dnasp5.0軟件[18]計(jì)算得出; 由軟件 Areliquin3.5[19]進(jìn)行群體的AMOVA分析; 生成單倍型數(shù)目、類型及分布以及pairwise difference模型下的遺傳分化參數(shù)。應(yīng)用NETWORKv 4.6軟件對波紋唇魚的2個(gè)線粒體基因單倍型進(jìn)行分析。
2 結(jié)果
2.1 COⅠ基因序列的分析
對4個(gè)波紋唇魚群體共101尾個(gè)體的COⅠ序列進(jìn)行測定, 經(jīng)ClustalX軟件編輯和比對后, 獲得了長度為1560 bp的同源序列。各群體COⅠ基因序列的A、T、G、C平均含量分別為24.05%、28.37%、18.54%和29.04%, 即A+T(52.42%)>G+C(47.58%)。遺傳多樣性參數(shù)統(tǒng)計(jì)顯示 101尾個(gè)體共有 23個(gè)變異位點(diǎn),占分析位點(diǎn)總數(shù)的1.5%, 共檢出20個(gè)單倍型, 總?cè)后w單倍型多樣性指數(shù)(Hd)為 0.629, 平均核苷酸差異數(shù)(K)為 1.195, 核苷酸多樣性指數(shù)(Pi)為 0.00077(表2), 顯示出波紋唇魚群體的遺傳多樣性水平偏低。
由AMOVA分析得出的直觀結(jié)論為群體間存在遺傳分化不明顯、群體內(nèi)部分化為變異主導(dǎo)的結(jié)果(表 4), 僅26.26%的變異來自群體間而73.74%的來自群體內(nèi)。群體內(nèi)及群體間的遺傳距離分析結(jié)果見表 3, 群體內(nèi)遺傳距離大小次序?yàn)? 海南陵水(0.00086)>南沙(0.00079)>西沙(0.00016)>馬來西亞(0.00000), 而群體間最大遺傳距離為陵水和南沙(0.00101), 略高于群體內(nèi)的遺傳距離。最小為馬來西亞和西沙(0.00008)。各群體間的遺傳分化指數(shù)(Fst)均較低, 支持了AMOVA分析關(guān)于群體間遺傳分化不明顯的結(jié)論, 由Nm= [(1/Fst)-1]/2計(jì)算得出的基因流均大于4[21], 同樣證實(shí)了以上結(jié)果。
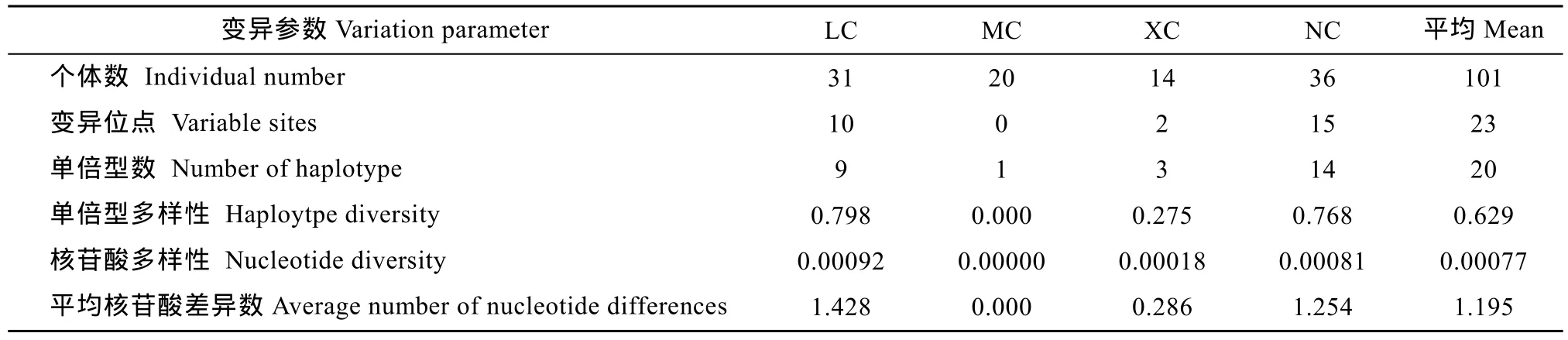
表2 波紋唇魚4個(gè)群體COⅠ基因的遺傳多樣性參數(shù)Tab. 2 Genetic diversity of four Cheilinus undulatus populations inferred from mtDNA COⅠ

表 3 波紋唇魚4個(gè)群體COⅠ基因群體間/群體內(nèi)的Kimura 2-parameter遺傳距離及遺傳分化Tab. 3 The average Kimura 2-parameter distance within and between populations and Fstof pairwise difference in CO Ⅰ sequences from four Cheilinus undulatus populations
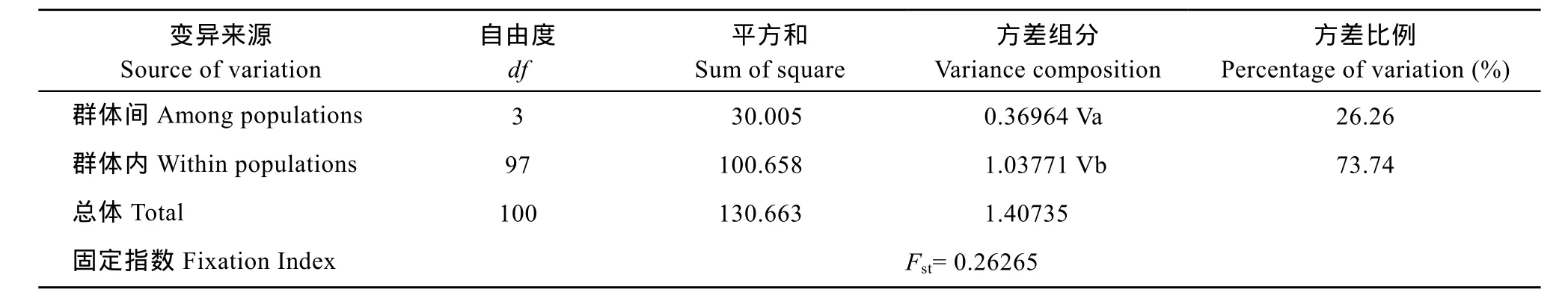
表4 波紋唇魚4個(gè)群體COⅠ基因的AMOVA分析Tab. 4 AMOVA analysis of all sequences from four Cheilinus undulatus populations of mt DNA CO geneⅠ
基于 COⅠ基因序列對波紋唇魚群體進(jìn)行單倍型分析, 共得出 20個(gè)單倍型, 其中共享單倍型有 4個(gè)(HAP1、HAP2、HAP16和HAP20), 所占比例達(dá)20.00% (4/20), 其中HAP1分布最廣泛, 包括海南陵水10尾個(gè)體、馬來西亞全部20尾個(gè)體、西沙12尾個(gè)體以及南沙17尾個(gè)體, 比例高達(dá)58.42% (59/101), HAP1甚至為馬來西亞的唯一單倍型, 除此之外, 其他共享單倍型均為 2種群共享。單倍型分布及頻率見表 5。由NETWORKv 4.6軟件生成的4個(gè)群體COⅠ基因的單倍型網(wǎng)絡(luò)圖呈星狀放射結(jié)構(gòu)(圖 1), 不同地理來源的單倍型無明顯分支, 呈交錯分布, 沒有體現(xiàn)地理差異性, 其中單倍型 HAP1 位于網(wǎng)絡(luò)圖的中心。
2.2 Cytb基因序列的分析
擴(kuò)增得到Cytb基因片段長度為1141 bp。經(jīng)軟件Dnasp4.10分析顯示101尾個(gè)體共有30個(gè)變異位點(diǎn), 27個(gè)單倍型, 總?cè)后w單倍型多樣性指數(shù)(Hd)為0.755, 平均核苷酸差異數(shù)(K)為 1.424, 核苷酸多樣性指數(shù)(Pi)為 0.00126 (表6), 顯示出波紋唇魚群體遺傳多樣性水平不高。
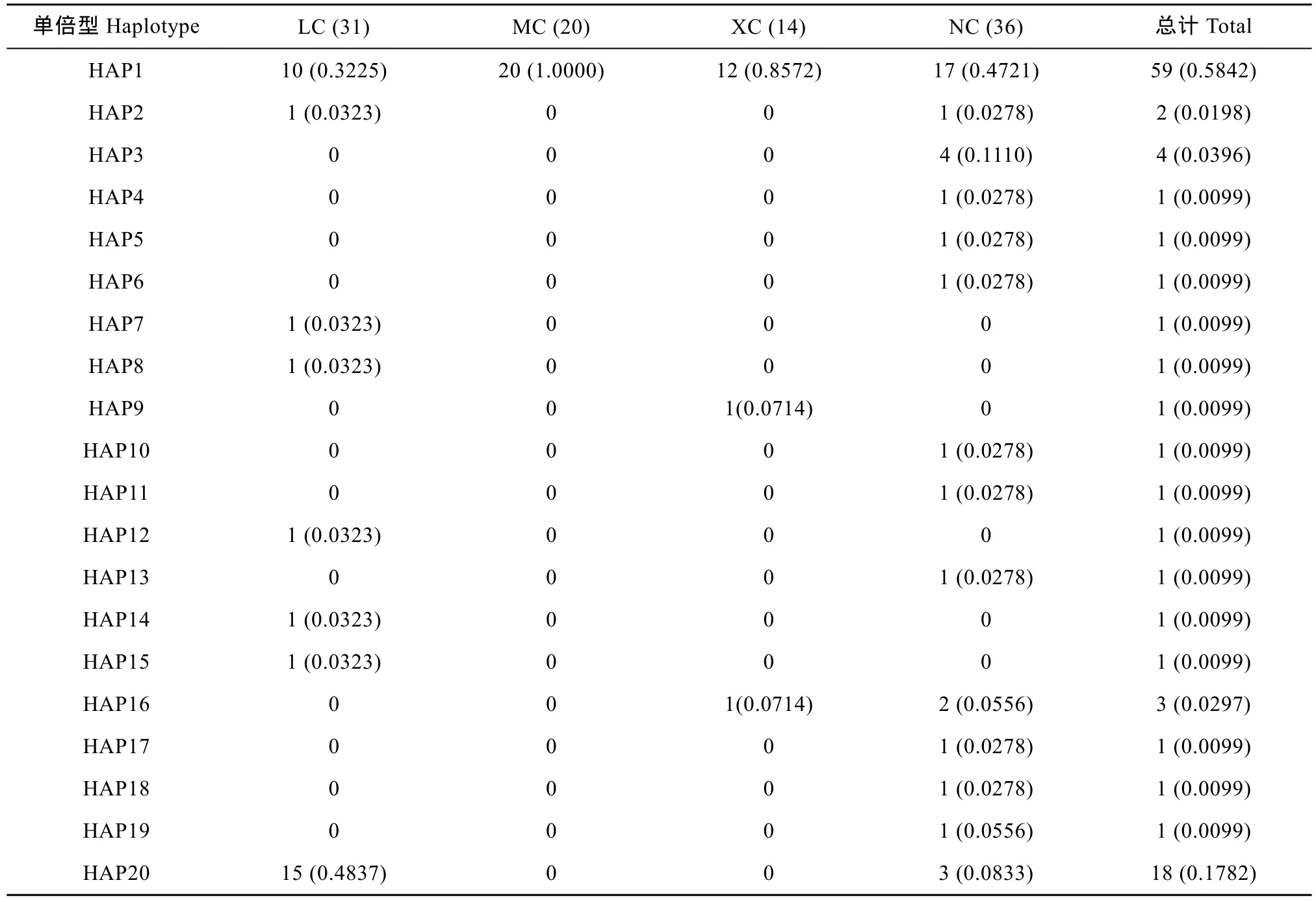
表5 波紋唇魚4個(gè)群體COⅠ基因的單倍型數(shù)目類型、頻率及分布Tab.5 Haplotype numbers and frequencies in four Cheilinus undulatus populations inferred from mtDNA COⅠ
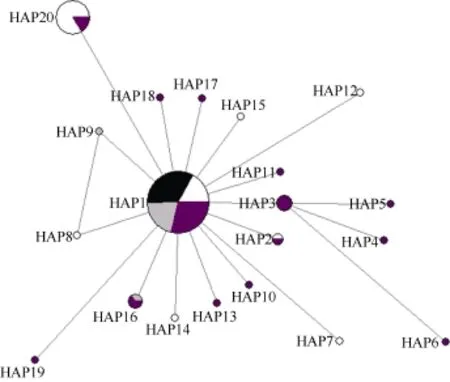
圖1 波紋唇魚4個(gè)群體COⅠ基因單倍型網(wǎng)絡(luò)圖Fig. 1 Haplotype network of Cheilinus undulatus from four populations inferred from mtDNA COⅠ
由AMOVA分析得出的直觀結(jié)論為群體間存在遺傳分化不明顯、群體內(nèi)部分化為變異主導(dǎo)的結(jié)果(表8), 僅4.22%的變異來自群體間而95.78%的來自群體內(nèi)。群體內(nèi)及群體間的遺傳距離分析結(jié)果見表7, 群體內(nèi)遺傳距離大小次序?yàn)? 南沙(0.00127)>海南 陵 水 (0.00125) >西 沙 (0.00089) >馬 來 西 亞(0.00082), 而群體間最大遺傳距離為陵水和南沙(0.00128), 略高于群體內(nèi)的遺傳距離。最小為馬來西亞和西沙(0.00089)。各群體間的遺傳分化指數(shù)(Fst)均較低, 支持了AMOVA分析關(guān)于群體間遺傳分化不明顯的結(jié)論, 雖然西沙和南沙間的 Fst(1.33236)處于0.05—015以上, 但其P值>0.05 (P=0.61798),即表明兩者之間存在遺傳分化但不顯著。
基于 Cytb 基因序列對波紋唇魚群體進(jìn)行單倍型分析, 共得出 27個(gè)單倍型, 其中共享單倍型有4個(gè)(HAP1、HAP6、HAP9和 HAP19), 所占比例達(dá)14.81% (4/27), 其中HAP19分布最廣泛, 包括海南陵水13尾個(gè)體、馬來西亞9尾個(gè)體、西沙8尾個(gè)體以及南沙14尾個(gè)體, 比例高達(dá)43.56% (44/101)。其他23個(gè)單倍型均為獨(dú)享單倍型, 單倍型分布及頻率見表 9。Cytb基因單倍型網(wǎng)絡(luò)圖結(jié)果(圖 2)與基于COⅠ基因片段構(gòu)建的單倍型網(wǎng)絡(luò)圖一致, 沒有體現(xiàn)地理差異性。
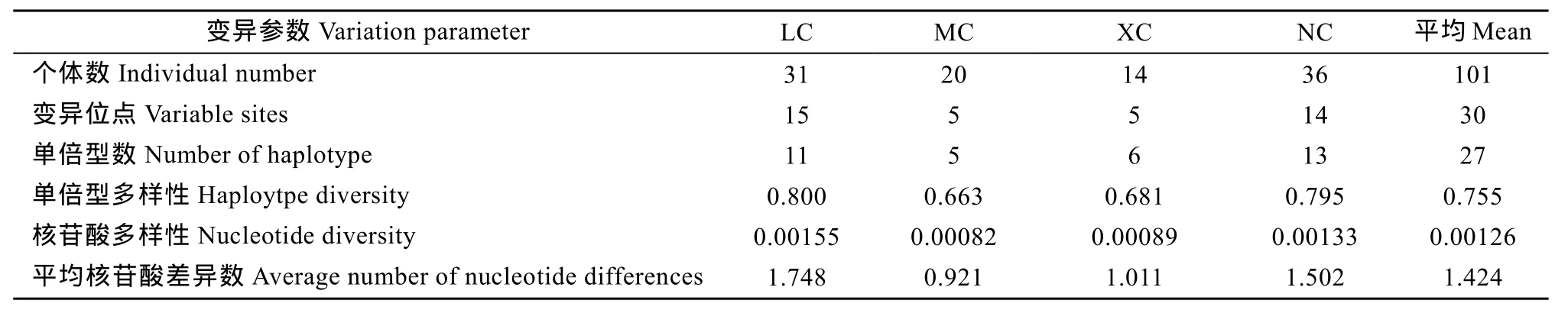
表6 波紋唇魚4個(gè)群體Cytb基因的遺傳多樣性參數(shù)Tab. 6 Genetic diversity of four Cheilinus undulatus populations inferred from mtDNA Cytb
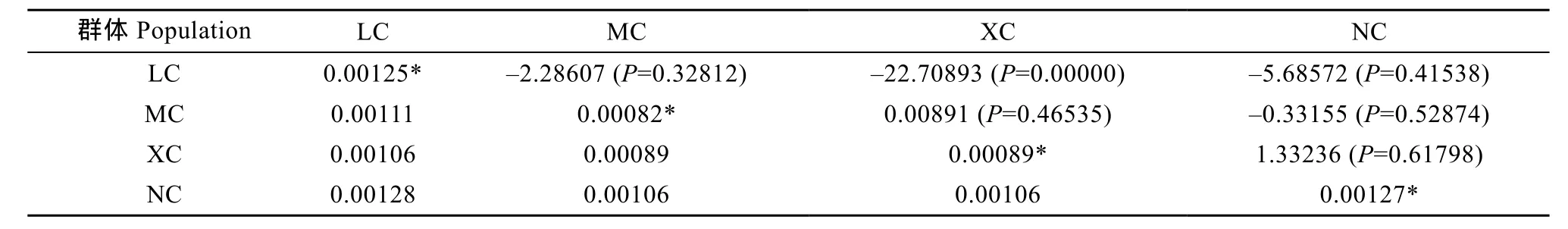
表7 波紋唇魚4個(gè)群體Cytb基因群體間/群體內(nèi)的Kimura 2-parameter遺傳距離及遺傳分化Tab.7 The average Kimura 2-parameter distance within and between populations and Fstof pairwise difference in Cytb sequences from four Cheilinus undulatus populations
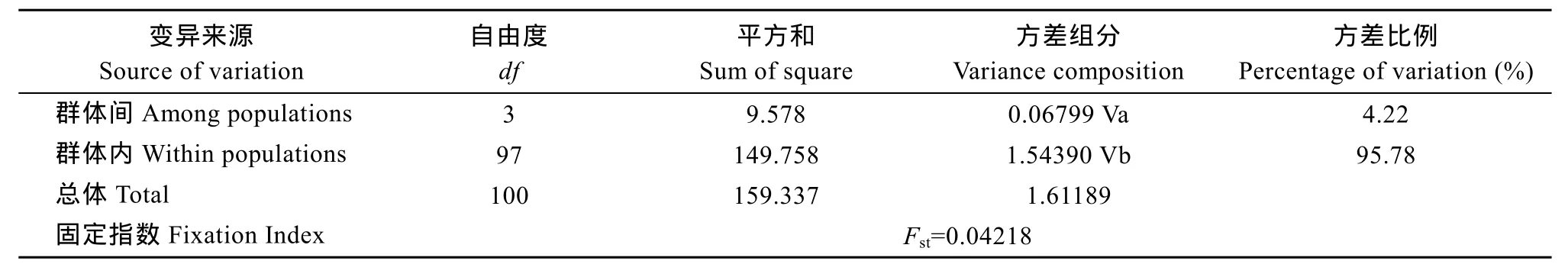
表8 波紋唇魚4個(gè)群體Cytb基因的AMOVA分析Tab. 8 AMOVA analysis of all sequences from four Cheilinus undulatus populations of mt DNA Cytb gene
3 討論
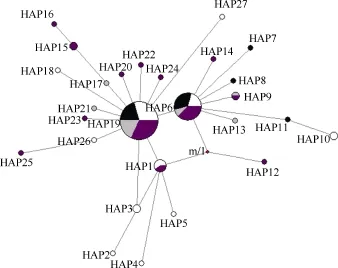
圖2 波紋唇魚4個(gè)群體Cytb基因單倍型網(wǎng)絡(luò)圖Fig. 2 Haplotype network of Cheilinus undulatus from four populations inferred from mtDNA Cytb
3.1 二個(gè)線粒體基因序列特征分析
2個(gè)線粒體基因分析得出的序列堿基含量均為A+T>G+C, Folmer等[22]認(rèn)為A+T 含量的增加也增加了密碼子第三位點(diǎn)上 A與 T顛換的可能性, 即A+T的含量越高, 線粒體DNA的進(jìn)化地位也高, 具有明顯進(jìn)化優(yōu)勢。動物線粒體基因組4種核苷酸的特性為不均一分布, 該研究結(jié)果體現(xiàn)了這一點(diǎn)。同時(shí)各群體 COⅠ基因序列的 A+T (52.42%)>G+C (47.58%), 與吳寧等[24]對6種鰻鱺COⅠ片段的研究結(jié)果相似[24]。各群體 Cytb基因序列的堿基含量結(jié)果A+T (50.82%)>G+C (49.18%), 與怒江瀕危魚類缺須盆唇魚[25]的結(jié)果相似, 而與閩浙地區(qū)香魚的研究結(jié)果[26]相反。可見, 不同的物種A+T與G+C的含量是不對等的。
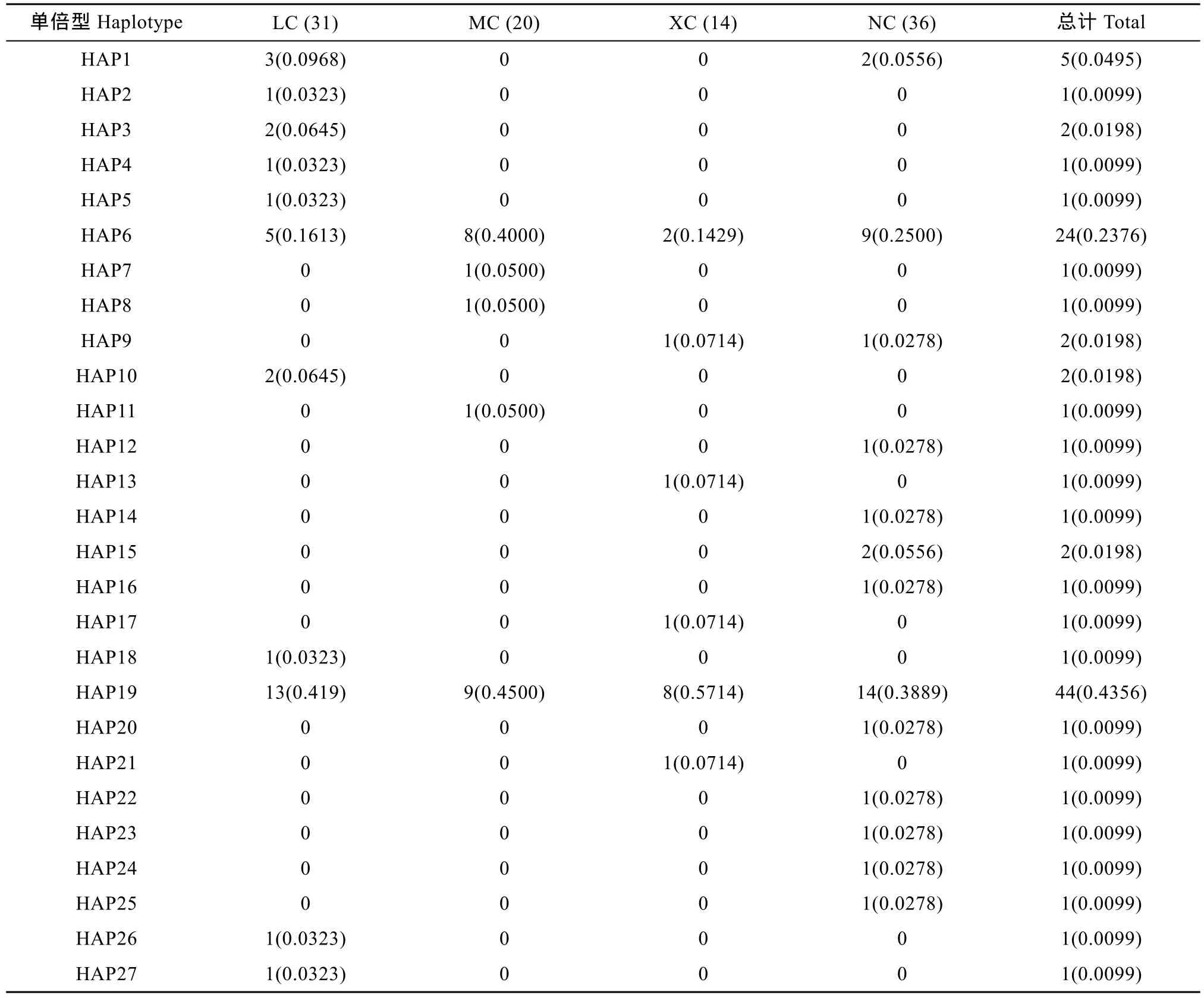
表9 波紋唇魚4個(gè)群體Cytb基因的單倍型數(shù)目類型、頻率及分布Tab. 9 Haplotype numbers and frequencies in four Cheilinus undulatus populations inferred from mtDNA Cytb
3.2 遺傳多樣性以及群體間的遺傳關(guān)系分析
本研究選取的2個(gè)mtDNA基因COⅠ和Cytb序列, 通過軟件計(jì)算得出的整體水平的遺傳多樣性參數(shù)S、Hd、K以及Pi, 結(jié)果表明COⅠ和Cytb的這些參數(shù)幾乎在同一水平上, COⅠ略小于Cytb, 證實(shí)了Meyer等[13]基于13個(gè)蛋白編碼mtDNA所含系統(tǒng)發(fā)育信息豐度的劃分, 即均為系統(tǒng)發(fā)育信息良好的一組, 其中包括Cytb、ND2、ND4、ND5和COⅠ。并且這 2個(gè)基因分析所得的遺傳分化及單倍型分布、網(wǎng)絡(luò)圖等參數(shù)基本在同一水平上, 都大同小異,即由 Fst及其 P值提示群體間存在分化, 但不顯著;單倍型在個(gè)體間均勻分布, 無明顯分支, 存在占有較大比例的單倍型共享型。因此, Cytb和COⅠ2個(gè)基因均較適于分析群體的遺傳結(jié)構(gòu)及系統(tǒng)發(fā)育。
遺傳多樣性即基因多樣性, 存在于生物個(gè)體內(nèi)、單個(gè)物種內(nèi)以及物種間, 是適應(yīng)性和進(jìn)化的基礎(chǔ), 種內(nèi)的遺傳多樣性與該物種對環(huán)境的適應(yīng)能力呈正向線性關(guān)系。單倍型間的平均遺傳距離(K)和核苷酸多樣性(Pi)是衡量遺傳多樣性2個(gè)重要指標(biāo), 而由于Pi值考慮各種mtDNA單倍型在群體中所占的比例的特性, 故其更能精確的揭示一個(gè)群體的mtDNA的多態(tài)程度[27]。Pi值越大表示群體多態(tài)程度越高, 反之亦然, 是衡量群體多態(tài)程度和群體遺傳分化的重要指標(biāo)之一。當(dāng)Pi值在0.0015—0.0047[28]時(shí), 則表明遺傳多樣性處于較低水平。本研究中由COⅠ、Cytb序列得出的整體水平 Pi值分別為0.00077和 0.00126, 均低于此范圍, 表明波紋唇魚的遺傳多樣性較低。首先, 波紋唇魚自身的建群效應(yīng)(Founder Effect)或種類差異可能導(dǎo)致上述現(xiàn)象,而人為因素更不可忽視。如人為的棲息地破壞, 尤其是對其棲息地珊瑚礁的破壞。作為一種大型珊瑚礁魚類, 其棲息地珊瑚礁的大面積的污染, 對該物種產(chǎn)生了致命的殺傷力。同時(shí)越來越先進(jìn)的捕撈技術(shù)和高殺傷力漁具也導(dǎo)致了其不可恢復(fù)的損傷, 甚至瀕臨消失。本文的研究對象僅為南中國海域的野生波紋唇魚, 由于珊瑚礁的破壞導(dǎo)致波紋唇魚的生活環(huán)境受到限制, 在一定程度上起到了生殖隔離的作用, 有一定的參考價(jià)值。但并不能忽略生殖交流對本結(jié)果的影響, 因此有必要進(jìn)一步對其他海域如非洲東海、紅海等的波紋唇魚進(jìn)行遺傳多樣性分析。
3.3 單倍型分布分析
溯祖理論[29]認(rèn)為分布最廣泛的單倍型為祖先類型, 而從COⅠ基因單倍型分析結(jié)果可看出HAP1為這4個(gè)野生波紋唇魚群體的主導(dǎo)單倍型。因此推測HAP1可能為這4個(gè)群體波紋唇魚的起源, 而HAP1又為馬來西亞群體全部個(gè)體單倍型, 由此可預(yù)測波紋唇魚的起源地可能為馬來西亞海域。同時(shí), 在COⅠ基因單倍型構(gòu)建的網(wǎng)絡(luò)圖中HAP1位于網(wǎng)絡(luò)圖的中心, 可推測這個(gè)單倍型為較為原始的單倍型, 其他單倍型由其衍生而來。而由COⅠ和Cytb基因序列對這4個(gè)群體的單倍型分析結(jié)果中均存在獨(dú)享單倍型, 暗示著群體間一定存在程度的遺傳分化。
結(jié)合COⅠ和Cytb基因序列分析得出的波紋唇魚101尾個(gè)體單倍型網(wǎng)絡(luò)圖可以看出: 4個(gè)地理群體的個(gè)體交錯在一起, 沒有體現(xiàn)明顯的地理差異性,單倍型分布 無明顯分支, 在個(gè)體間呈均勻交錯分布, 這也暗示了群體間遺傳分化水平低, 進(jìn)行了廣泛的基因交流。
利用線粒體 COⅠ和 Cytb基因作為分析手段,本研究對海南陵水、馬來西亞、西沙和南沙4個(gè)不同地理群體的波紋唇魚進(jìn)行了遺傳多樣性和遺傳分化研究。結(jié)果表明, 波紋唇魚的遺傳分化存在但不顯著, 且遺傳多樣性處于較低水平。受制于取材, 本研究不能完全反應(yīng)該物種群體分化程度和遺傳多樣性水平, 也不能衡量人為因素對其遺傳多樣性的影響, 但一定程度上為波紋唇魚的種質(zhì)資源保護(hù)工作提供了科學(xué)依據(jù), 也為后續(xù)的研究提供了參考資料。
[1] Shen S J. Fishes of Taiwan [M]. Taipei: Department of Animal Science at Taiwan University. 1993, 143—144 [沈世杰. 臺灣魚類志. 臺北: 臺灣大學(xué)動物學(xué)系. 1993, 143—144]
[2] Jie X Y, Li C H, Xu Z N, et al. Industrial aquaculture of famous and expensive marine fishes [J]. Hunan Agricultural Sciences, 2011, (11): 141—143 [頡曉勇, 李純厚, 許忠能,等. 名貴海水魚類工廠化養(yǎng)殖試驗(yàn)研究. 湖南農(nóng)業(yè)科學(xué), 2011, (11): 141—143]
[3] Qi X Z, Yin S W, Luo J, et al. Complete mitochondrial genome sequence of the humphead wrasse, Cheilinus undulatus [J]. Genetics and Molecular Research, 2013, 12(2): 1095—1105
[4] Qu Y J, Li J E. Analysis and evaluation of nutrition composition of double-headed parrotfish Cheilinus undulatus [J]. Journal of Tropical Oceanography, 2010, 29(3): 97—102 [區(qū)又君, 李加兒. 野生波紋唇魚 Cheilinus undulatus 營養(yǎng)成分分析與評價(jià). 熱帶海洋學(xué)報(bào), 2010, 29(3): 97—102]
[5] Donaldson T J, Sadovy Y. Threatened fishes of the world: Cheilinus undulatus Rüppell, 1835 (Labridae) [J]. Environmental Biology of Fishes, 2001, 62: 428
[6] Sadovy Y, Kulbicki M, Labrosse P, et al. The humphead wrasse, Cheilinus undulatus: synopsis of a threatened and poorly known giant coral reef fish [J]. Reviews in Fish Biology and Fisheries, 2003, 13: 327—364
[7] Hu J, Qi X Z, Yin S W, et al. The variation analysis of mtDNA D-loop sequence of Cheilinus undulatus [J]. Marine Sciences, 2012, 35(4): 50—56 [胡靜, 齊興柱, 尹紹武, 等.波紋唇魚mtDNA D-loop序列變異分析. 海洋科學(xué), 2012, 35(4): 50—56]
[8] Hu J, Zhu X P, Yin S W, et al. Development and characterization of microsatellite loci in a threatened marine fish, Cheilinus undulatus (humphead wrasse) [J]. Genetics and Molecular Research, 2013, 12(3): 2633—2636
[9] Guo X H, Liu S J, Liu Q, et al. New progresses on Mitochondrial DNA in fish [J]. Acta Genetica Sinica, 2004, 31(9): 983—1000 [郭新紅, 劉少軍, 劉巧, 等. 魚類線粒體DNA研究新進(jìn)展. 遺傳學(xué)報(bào), 2004, 31(9): 983—1000]
[10] Liu S F, Chen L L, Dai F Q, et al. Applicaction of DNA Barcoding gene COⅠ for classifying family Sciaenidae [J]. Oceanologia et Limnologia Sinica, 2010, 41(2): 223—232 [柳淑芳, 陳亮亮, 戴芳群, 等. 基于線粒體 COⅠ基因的DNA條形碼在石首魚科(Sciaenidae)魚類系統(tǒng)分類中的應(yīng)用. 海洋與湖沼, 2010, 41(2): 223—232]
[11] Peng J L, Wang X Z, Wang D, et al. Application of DNA barcoding based on the Mitochondrial CO Ⅰ gene sequences in classification of Culter (Pisces: Cyprinidae) [J]. Acta Hydrobiologica Sinica, 2009, 33(2): 271—276 [彭居俐, 王緒禎, 王丁, 等. 基于線粒體COⅠ基因序列的DNA條形碼在鯉科 鲌屬魚類物種鑒定中的應(yīng)用. 水生生物學(xué)報(bào), 2009, 33(2): 271—276]
[12] Irwim D, Kocher T, Wilson A. Evolution of the cytochrome b gene of mammals [J]. Journal of Molecular Evolution, 1991, 32(2): 128—144
[13] Meyer A, Kocher T D, Basasibwaki P, et al. Monophyletic origin of Lake Vietoria cichlid fishes suggested by mitochondrial DNA sequences [J]. Nature, 1990, 347: 550—553
[14] Gao T X, Bi X X, Zhao L L, et al. Population genetic structure of Roughskin sculpin Trachidermus fasciatus based on the mitochondrial Cytb sequence [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 199—207 [高天翔, 畢瀟瀟, 趙林林, 等. 基于線粒體Cytb基因全序列的松江鱸群體遺傳結(jié)構(gòu)分析. 水生生物學(xué)報(bào), 2013, 37(2): 199—207]
[15] Lybeard C, Roe K J. The phylogenetic utility of the mitochondrial cytoehrome b gene for inferring relationships among Actinopterygian fishes: In: Koeher T D, Stepien C A (Eds.), Molecular Systematies of Fishes [M]. San Diego: Academic Press. 1997, 285—303
[16] Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: a Laboratory Manual [M]. Beijing: Science Press. 1995, 27—29 [薩姆布魯克J, 弗里奇 E F, 曼尼阿蒂斯T. 分子克隆實(shí)驗(yàn)指南. 北京: 科學(xué)出版社. 1995, 27—29]
[17] Tamura K, Peterson D, Peterson N, et al. MEGA5: Molecular evolutionary analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods [J]. Molecular Biology and Evolution, 2011, 28: 2731—2739
[18] Librado P, Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data [J]. Bioinfomatics, 2009, 25: 1451—1452
[19] Excoffier L, Laval G, Schneider S. Arlequin Ver.3.0: an integrated software package for population genetics data analysis [J]. Evolution Bioinformation, 2005: 47—50
[20] Wright S. The genetical structure of population [J]. Annals of Eugenics, 1951, 15: 323—334
[21] Millar C L, Libby W J. Strategies for Conserving Clinal, Ecotypic, and Disjunct Population Diversity in Widespread Species. In: Fald D A, Holsinger K E (Eds.), Genetics and Conservation of Rare Plants [M]. New York: Oxford University Press. 1991, 149—170
[22] Folmer O, Black M, Hoeh W, et al. DNA primer for amplification of mitochondrial cytochrome coxidase subunitⅠfrom diverse metazoan invertebrates [J].Molecular Marine Biology Biotechnology, 1994, 3(5): 294—299
[23] Brown W M. The Mitochondrial Genome of Animals [M]. New York: Plenum Press. 1985, 102—110
[24] Wu N, Li Z B, Lin X Y, et al. Comparative study on MtDNA CO Ⅰgene fragment of sixAnguilla [J]. Oceanologia et Limnologia Sinica, 2010, 11(41): 930—934 [吳寧, 黎中寶,林小云, 等. 6種鰻鱺(Anguilla) 線粒體 DNA COⅠ序列的比較研究. 海洋與湖沼, 2010, 11(41): 930—934]
[25] Zhang D Y, Wang D Q, Zhang S P, et al. Populationgenetic structure analysis of endangered species Placocheilus cryptonemus in the Nujiang River based on Cytb sequences of mtDNA [J]. Journal of Fishery Sciences of China, 2009, 7(16): 477—485 [張東亞, 汪登強(qiáng), 張紹平, 等. 怒江瀕危魚類缺須盆唇魚基于 Cytb序列的群體遺傳結(jié)構(gòu)分析. 中國水產(chǎn)科學(xué), 2009, 7(16): 477—485]
[26] Li N, Chen S B, Xie Q L, et al. Polymorphisms of mitochondrial Cytb gene and D-loop region in sweetfish (Plecoglossus altivelis Temminck et Schlegel) from Zhejiang and Fujian Provinces [J]. Hereditas, 2008, 30(7): 919—925 [李娜, 陳少波, 謝起浪, 等. 閩浙地區(qū)香魚線粒體Cytb基因和 D-loop區(qū)序列多態(tài)性分析. 遺傳, 2008, 30(7): 919—925]
[27] Xiao M S, Cui F, Kang J, et al. Analysis on sequence polymorphism of the mitochondrial DNA control region and population genetic diversity of the cultivated and natural Chinese longsnout catfish (Leiocassis longirostris) [J]. Acta Hydrobiologica Sinica, 2013, 37(1): 90—99 [肖明松, 崔峰,康健, 等. 長吻(魚危)養(yǎng)殖群體與野生群體遺傳多樣性分析. 水生生物學(xué)報(bào), 2013, 37(1): 90—99]
[28] Lan H, Shi L M. The origin and genetic differentiation of native breeds of pigs in south west China: An approach from mitochondrial DNA polymorphism [J]. Biochemistry Genetics, 1993, 31: 51—60
[29] Crandall K A, Templeton A R. Empirical tests of some prediction from coalescent theory with application to intraspecific phylogeny reconstruction [J]. Genetics, 1993, 134(3): 959—969
GENETIC DIVERSITY AND DIVERGENCE OF CHEILINUS UNDULATUS OF DIFFERENT GEOGRAPHIC POPULATIONS OF THE SOUTH CHINA SEA REVEALED BY COⅠ AND CYTB GENE ANALYSES
HU Jing1,2, HOU Xin-Yuan1, YIN Shao-Wu1, ZHU Fei1, JIA Yi-He1and WANG Xiao-Jun1
(1. College of Life Sciences, Nanjing Normal University, Nanjing 210023, China; 2. Tropical Fishes Research and Development Center, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Sanya 572018, China)
The genetic diversity of Cheilinus undulatus was studied based on the sequence analysis of mtDNA (CO Ⅰand Cytb) of 101 individuals from four geographical populations including Lingshui (n=31), Malaysia (n=20), Xisha (n=14), and Nansha (n=36). Using PCR amplification and sequencing, 1560 bp of CO Ⅰand 1141 bp of Cytb sequences were obtained. For COⅠsequences, the number of haplotype is 20, haplotype diversity was 0.629, the nucleotide diversity was 0.00077, the average nucleotide difference was 1.195, and the number of polymorphic site was 23. For Cytb sequences, the number of haplotype was 27, the haplotype diversity was 0.755, the nucleotide diversity was 0.00126, the average nucleotide difference was 1.424, and the number of polymorphic site was 30. AMOVA showed a higher percentage of 73.74% with-population variation than that of 26.26% among-population variation in CO Ⅰsequences.Similarly, the percentage of 95.78% within-population variations was higher than that of 4.22% the among-population variation in Cytb sequences. The haplotype network of CO Ⅰ and Cytb genes with stellate radiation structure suggested that haplotypes distribution did not have the characteristics of clear geographical differentiation. Genetic diversity was low and the genetic divergence was inconspicuous between populations of C. undulatus. These results may be very useful to protect this species.
Cheilinus undulatus; CO ; CytⅠ b; Genetic diversity; Genetic divergence
Q346+.5
A
1000-3207(2014)06-1008-09
10.7541/2014.149
2013-09-02;
2014-04-06
國家自然科學(xué)基金項(xiàng)目(40966003); 海南省自然科學(xué)基金(809009); “十一五” 國家科技支撐計(jì)劃重點(diǎn)項(xiàng)目(2007BAD29B03);江蘇省高校優(yōu)勢學(xué)科建設(shè)工程項(xiàng)目資助
胡靜 (1987—), 女, 湖北武漢人; 碩士; 主要從事魚類種質(zhì)資源與遺傳育種研究。E-mail: hijklnm123@sina.com
尹紹武(1969—), 男, 博士, 教授; 主要從事魚類種質(zhì)資源與遺傳育種研究。E-mail: yinshaowu@163.com
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
